
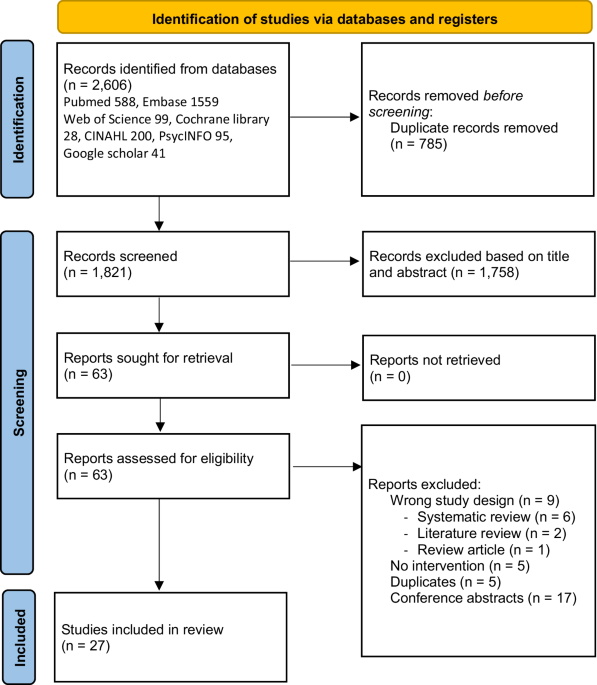
記住我
Two patient-derived primary tumor PDAC organoids (hPT1 and hPT2) were acquired and expanded for this study (Fig. 1a). The doubling time of both organoids was about 7 days, hence the organoids were passaged at a 1:2 ratio weekly. Immunostaining clearly showed that the organoids were positive for both cytokeratin and epithelial cell adhesion molecule (EpCAM), reflecting their epithelial origin (Fig. 1b). To analyze their chromosome copy number profiles, the genomic DNA of both hPT1 and hPT2 organoids was subjected to single-nucleotide polymorphism microarray (SNPa), and the data was used to derive their copy number profiles at 1 Mb resolution (Fig. 1c). We previously benchmarked this method and demonstrated that SNPa produced similar results as the comparative genome hybridization array22, which is the gold standard for chromosome copy number profile measurements23. Chromosome copy number alterations were observed across the genome of both hPT1 and hPT2 (Fig. 1c). The total chromosome number was estimated from the summation of the average copy number of each chromosome. In general, copy gains were more prevalent than losses, resulting in a sub-triploid ploidy of 57.1 and 57 chromosomes for hPT1 and hPT2, respectively. Additionally, we observed unusually high copy number alterations in chromosome 8 of hPT2, where the copy number appeared to oscillate between losses and very high gains. This pattern is in line with the typical signatures of chromothripsis, as reported in numerous cancers, including PDAC24,25. The role of chromothripsis in PDAC metastatic progression was suggested through the amplification of MYC at chromosome 826. Indeed, the copy number of MYC in hPT2 is higher than in hPT1 (Fig. 1c).
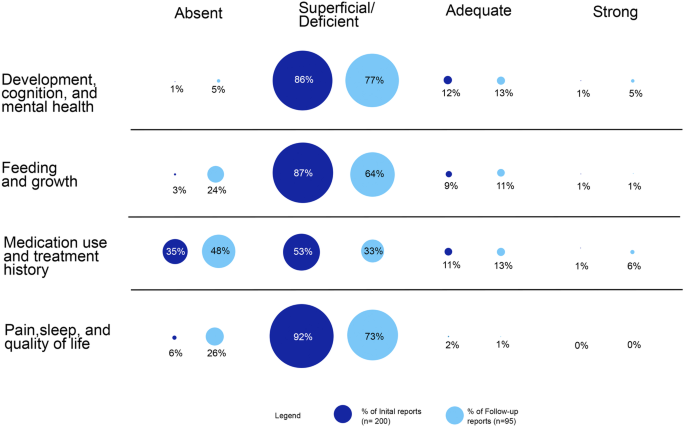
Fig. 1: Chromosome copy number alterations in patient-derived pancreatic ductal adenocarcinoma (PDAC) organoids.
a Representative images of primary tumor-derived PDAC organoids, hPT1 and hPT2, from two independent patients, showing the growth of the organoids over 4 days (bar = 200 µm). b Confocal sections of hPT1 and hPT2 organoids immunostained for cytokeratin (red) and EpCAM (green), both are markers of epithelial and PDAC cells (bar = 25 µm). c Genomic DNA of passage 3 of hPT1 and passage 6 of hPT2 was subjected to SNPa, and the resulting data was used to derive chromosome copy numbers of the organoids within 1 Mb binning windows. The heatmaps illustrate chromosome copy numbers across the whole genome, where blue indicates chromosome loss (<2) and red indicates chromosome gain (>2). The copy number of the frequently mutated genes in PDAC is indicated by the color of the corresponding arrowheads. The total chromosome number was estimated from the summation of the average copy number of each chromosome. Chromosome copy number alterations were observed across the genome, and both organoids had sub-triploid ploidy of ~57 chromosomes.
The copy number of some genes, including the four most common driver mutation genes27: KRAS, CDKN2A, TP53, and SMAD4, are also frequently altered in PDAC28. In both hPT1 and hPT2 (Fig. 1c), the oncogene KRAS is amplified, while the tumor suppressors CDKN2A and TP53 have reduced copy numbers. Interestingly, the copy loss of the tumor suppressor SMAD4 was only observed in hPT1, suggesting a potential phenotypic distinction between hPT1 and hPT2. Differences in copy number alterations were also observed in the other frequently altered genes, including ARID1A, TGFBR2, CTNNB1, RREB1, EGFR, and BRAF.
Genomic heterogeneity in PDAC organoids and genomic shift with prolonged cultureSNPa provides a bulk-level overview of the average copy numbers of all the cells in the PDAC organoid. To improve our cellular resolution in understanding the genomic heterogeneity of PDAC organoids, we performed scWGS on both hPT1 and hPT2. We further examined each organoid line at two passage numbers to gain insights into the genomic stability of organoid culture.
From passage 3 of hPT1 organoids (hPT1 P3), we acquired scWGS data from 705 cells. We derived the copy number profile of each cell at 5 Mb resolution. Of note, the resolution of the copy number profile from scWGS is lower than that from SNPa. Averaging the sequencing data at a bigger 5 Mb window is necessary due to the relatively shallow sequencing depth, ranging between 0.015× and 0.042× for the scWGS samples. To systematically analyze the genomic heterogeneity within each sample, we performed t-SNE analysis on the single-cell copy number profiles, followed by spectral clustering to derive clusters of closely related cells. The spectral clustering of the hPT1 P3 t-SNE plot resulted in five clusters (Fig. 2a). Genomic heterogeneity was clearly observed within the hPT1 P3 population. Specifically, two distinct groups of cells were observed. The first group of cells was contained in Clusters 1 and 2, totaling 61 cells. Compared to the rest of the population, the cells in Clusters 1 and 2 had partial copy gains in chromosomes 1, 3, 7, 8, and 16, and whole chromosome copy gains in chromosomes 11, 12, 13, and 20. The second group of cells was contained in Clusters 3–5, totaling 644 cells. Genomic heterogeneity within this group was observed through the intermittent copy losses across the cells within chromosomes 1, 3, 6, 7, 9, 14, 18, 21, and X, as well as copy gains within chromosomes 3, 8, 13, and 15. When we averaged the single-cell copy number data from the 705 hPT1 P3 cells (Fig. 2a bottom), the resulting copy number profile is in accordance with the SNPa copy number profile of hPT1 (Fig. 1c), confirming the concordance with the scWGS method. One difference between the aggregated scWGS profile and the SNPa profile is that the estimated number of chromosomes from the scWGS data is higher than the one estimated by SNPa. This slight discrepancy may be because they were derived from different methods and/or because of the different copy number resolutions, 5 Mb for scWGS and 1 Mb for SNPa. Nevertheless, the higher scWGS estimate is still within the sub-triploid ploidy range.
Fig. 2: Single-cell whole-genome sequencing reveals genomic heterogeneity within PDAC organoids and the genomic shift with culture.
a From passage 3 of hPT1 (hPT1 P3), we acquired scWGS data of 705 cells. The single-cell copy number data derived from the scWGS were subjected to t-SNE analysis, followed by spectral clustering to derive the number of clusters within the sample. The spectral clustering of the hPT1 P3 t-SNE plot resulted in five distinctive clusters. The single-cell chromosome copy number profiles of each cluster were plotted in the form of heatmaps, with the resolution of a 5 Mb binning window. Genomic heterogeneity is evident within the population. Clusters 1 and 2 show a distinct group of cells (totaling 61 cells) with partial chromosome copy gains in chromosomes 1, 3, 7, 8, and 16, in addition to whole chromosome copy gains in chromosomes 11, 12, 13, and 20. Genomic heterogeneity can also be observed in the rest of the cells (Clusters 3–5, totaling 644 cells) through the intermittent copy losses within chromosomes 1, 3, 6, 7, 9, 14, 18, 21, and X, as well as intermittent copy gains within chromosomes 3 and 15. The bottom heatmap shows the averaged copy number from the 705 hPT1 P3 cells. The copy number pattern across the genome is in accordance with the hPT1 copy number profile derived from the SNPa in Fig. 1c. The total number of chromosomes was estimated to be 62.9, which is slightly higher than the one estimated by the SNPa, but both of them are still within the sub-triploid ploidy range. b From passage 14 of hPT1 organoids (hPT1 P14), we acquired scWGS data of 744 cells. The spectral clustering of the hPT1 P14 t-SNE plot resulted in six distinctive clusters. The copy number profiles of hPT1 P14 Clusters 1–3 resemble the cells in hPT1 P3 Clusters 1 and 2, however, the proportion of these cells increases within the hPT1 P14 population (totaling 303 cells), suggesting a clonal population expansion with the extended culture of 11 passages. Genomic heterogeneity within the hPT1 P14 Clusters 1–3 cells can be observed in chromosomes 1, 4, 5, 11, 13, 16, and 22. The copy number profiles of the rest of the cells (Clusters 4–6, totaling 441 cells) look similar to the hPT1 P3 Clusters 3–5 cells, except for the partial loss of chromosome 8 within all of the hPT1 P14 Cluster 4 cells and some cells in Clusters 5 and 6. The bottom heatmap shows the averaged copy number from the 744 hPT1 P14 cells. The total number of chromosomes was estimated to be 64.4, which is higher than hPT1 PT3, indicating copy number gains with extended culture. c The CNV between hPT1 P3 and hPT1 P14 was derived from the averaged copy number data. Chromosome copy gains were observed in chromosomes 1, 3, 6, 7, 8, 9, 11, 12, 13, 14, 16, 17, 18, 20, 21, and X, which correspond to the clonal expansion in hPT1 P14 Cluster 1–3. All combined, the extended culture resulted in approximately 62.7 Mb copy gains and 20.9 Mb copy losses. d From passage 6 of hPT2 (hPT2 P6), we acquired scWGS data of 873 cells. The spectral clustering of the hPT2 P6 t-SNE plot resulted in five distinctive clusters. Similar to hPT1 organoids, genomic heterogeneity can be observed within the hPT2 population. In contrast to the other clusters, the cells within Cluster 1 have intermittent copy losses in chromosome 18, instead of copy gains. Eleven cells within Cluster 1 have uniquely partial gains in chromosome 9 and partial diploid in chromosome 4. The copy number profiles of the rest of the cells (Clusters 2–5, totaling 788 cells) are generally similar to each other, with some level of genomic heterogeneity in the form of intermittent copy losses within chromosomes 3, 9, 13, 14, 17, 19, and X. The bottom heatmap shows the averaged copy number from the 873 hPT2 P6 cells, which is in accordance with the hPT2 copy number profile derived from the SNPa in Fig. 1c. The total number of chromosomes was estimated to be 64.5. e From passage 16 of hPT2 organoids (hPT2 P16), we acquired scWGS data of 999 cells. The spectral clustering of the hPT2 P16 t-SNE plot resulted in eight distinctive clusters. The copy number profile of hPT2 P16 Clusters 1, 2, and 8 (totaling 356 cells) resemble the cells in hPT2 P6 Clusters 2–5, with the addition of copy gains in chromosome 20 in some cells. The rest of the cells in Clusters 3–7 (totaling 643 cells) have partial copy gains in chromosome 9, resembling the 11 cells in hPT2 P6 Cluster 1, again suggesting a clonal population expansion with the extended culture. However, in contrast to the cells in hPT2 P6, these 643 cells also have copy gains in chromosomes 4 and 18. The bottom heatmap shows the averaged copy number from the 999 hPT2 P16 cells. The total number of chromosomes was estimated to be 65.3, which is higher than hPT2 P6, indicating copy number gains with extended culture. f The CNV between hPT2 P6 and hPT2 P16 was derived from the averaged copy number data. Significant chromosome copy gains were observed within chromosome 9, which were caused by the clonal expansion in hPT2 P16 Clusters 3–7. Other copy gains were also observed in chromosomes 3, 16, 18, and 20. The extended culture resulted in approximately 25.7 Mb copy gains and 9.8 Mb copy losses.
Next, we performed scWGS analysis on passage 14 of hPT1 organoids (hPT1 P14). As we passaged the organoids weekly, hPT1 P3 was cultured for 11 weeks to reach hPT1 P14. At P14, we acquired scWGS data from 744 cells. The spectral clustering of the hPT1 P14 t-SNE plot resulted in six clusters (Fig. 2b). From the copy number profiles, two distinct groups of cells were also observed in hPT1 P14. The copy number profiles of the first group of cells in Clusters 1–3 resembled the cells in hPT1 P3 Clusters 1 and 2. However, the proportion of these cells in hPT1 P14 grew to ~40.7% (303 cells out of 744 cells) from ~8.7% in hPT1 P3, suggesting a clonal population expansion with extended culture. Genomic heterogeneity within the hPT1 P14 Clusters 1–3 cells was observed within chromosomes 1, 4, 5, 11, 13, 16, and 22, which were absent in hPT1 P3 Clusters 1 and 2 cells. The copy number profiles of the 441 cells in hPT1 P14 Clusters 4–6 resembled the hPT1 P3 Clusters 3–5 cells, with the exception of the partial loss of chromosome 8 within all of the hPT1 P14 Cluster 4 cells and some of the cells in Clusters 5 and 6. In all, the total number of chromosomes was estimated to be 64.4, which is slightly higher than in the hPT1 P3, indicating copy number gains with the extended culture. Indeed, when we derived the CNV between hPT1 P3 and P14 from the averaged copy number profile (Fig. 2c), 62.7 Mb copy gains were observed across chromosomes 1, 3, 6, 7, 8, 9, 11, 12, 13, 14, 16, 17, 18, 20, 21, and X and 20.9 Mb copy losses were observed in other chromosomes. Most of the observed genomic shifts correspond to the clonal expansion in hPT1 P14 Clusters 1–3.
Similar scWGS analyses were performed for the hPT2 organoids. From passage 6 of hPT2 organoids (hPT2 P6), we acquired scWGS data from 873 cells. The spectral clustering of the hPT2 P6 t-SNE plot resulted in five clusters (Fig. 2d). Three distinct groups of cells were observed within the single-cell copy number profiles of hPT2 P6 cells. The differences between these three groups of cells revolved around chromosomes 4, 9, and 18. First, the 85 cells within Cluster 1 had intermittent copy losses in chromosome 18, instead of the copy gains that were observed in the other clusters. Second, within Cluster 1, 11 cells had uniquely partial gains in chromosome 9, accompanied by partial diploid in chromosome 4. The rest of the 788 cells in Clusters 2–5 had copy gains in chromosomes 4 and 18, and intermittent losses in chromosome 9. Genomic heterogeneity in these 788 cells was observed in the form of intermittent copy losses within chromosomes 3, 9, 13, 14, 17, 19, and X. The unusual oscillating copy number profile in chromosome 8, as illustrated in Fig. 1c, was also observed under scWGS in all cells. Furthermore, the averaged copy number profile of hPT2 P6 under scWGS (Fig. 2d bottom) also highly resembled the SNPa copy number profile of hPT2 P6 in Fig. 1c. The total number of chromosomes for the hPT2 P6 cells was estimated to be 64.5, again, slightly higher than the number estimated by the SNPa copy number profile (Fig. 1c). After an extended culture for 10 additional passages, the scWGS data of hPT2 P16 revealed a clonal expansion of the cells that had partial copy gains in chromosome 9, overtaking the population majority from ~1.3% in hPT2 P6 to ~64.4% in hPT2 P16 (Clusters 3–7, totaling 643 cells out of 999 cells, Fig. 2e), again suggesting a clonal population expansion with extended culture. However, in contrast to hPT2 P6, these 643 hPT2 P16 cells had copy gains in both chromosomes 4 and 18. The rest of the 356 cells in hPT2 P16 Clusters 1, 2, and 8 resembled the cells in hPT2 P6 Clusters 2–5, with the addition of copy gains in chromosome 20 in some cells. From the averaged copy number profile, we estimated the chromosome number of hPT2 P16 to be 65.3, which was slightly higher than the number estimated for hPT2 P6, again suggesting a gain of copy number with extended culture. Indeed, from the CNV analysis between hPT2 P6 and P16 (Fig. 2f), significant copy gains were observed within chromosome 9, which were caused by the clonal expansion in hPT2 P16 Clusters 3–7, and also other copy gains in chromosomes 3, 16, 18, and 20. All combined, the extended culture resulted in approximately 25.7 Mb of copy gains and 9.8 Mb of copy losses.
Next, to investigate the evolutionary distance between the clusters of cells, the copy number profiles of each cluster were averaged and used to derive phylogenetic trees for hPT1 and hPT2. The maximum parsimony method29 was used to build the trees from the early passage clusters, followed by the minimal distance pairings of the late passage clusters. This approach enabled us to reveal the correlation between the clusters of cells from different culture periods and to quantify the copy number differences between the clusters, as a measure of genomic heterogeneity. The hPT1 phylogenetic tree revealed two distinct groups of clusters that were separated by ~1500 Mb of copy number differences (Fig. 3a). Closer to the diploid root, the first group includes hPT1 P3 Clusters 3–5 (P3 C3–5), and hPT1 P14 Clusters 4–6 (P14 C4–6). P14 C5–6 are paired to P3 C4, while P14 C4 is paired to P3 C5. These pairings indicate the correlation between the late passage clusters to the early ones, and again confirm the existence of the same clonal population across the extended culture. The estimated chromosome number ranges from 56.1 to 62.8. The other group of clusters includes P3 C1–2, and P14 C1–3, which are paired to P3 C1. The estimated chromosome number ranges from 67.3 to 71.1. Unlike the first group, the clusters in this second group spanned within ~500 Mb of copy number differences, indicating a lower level of genomic heterogeneity within these clusters.
Fig. 3: Phylogenetic tree analysis reveals the correlation between the identified sub-population clusters and enables the quantification of genomic heterogeneity.
The single-cell chromosome copy number data of the clusters identified in Fig. 2 were averaged and used to derive phylogenetic trees for hPT1 and hPT2. The maximum parsimony method was used to build the trees from the early passage clusters, followed by the minimal distance pairings of the late passage clusters. These trees reveal the correlation between the clusters of different culture periods and quantify the copy number differences between the clusters, as a measure of genomic heterogeneity. a hPT1 phylogenetic tree reveals two distinct groups of clusters that are separated by ~1500 Mb of copy number differences. Closer to the diploid (2 N) root, the first group includes hPT1 P3 Clusters 3–5 (P3 C3–5), and hPT1 P14 Clusters 4–6 (P14 C4–6). P14 C5–6 are paired to P3 C4, while P14 C4 is paired to P3 C5. The estimated chromosome number ranges from 56.1 to 62.8. The other group of clusters, spread within ~500 Mb of copy number differences, includes P3 C1–2, and P14 C1–3, which are paired with P3 C1. The estimated chromosome number ranges from 67.3 to 71.1. b The clusters in the hPT2 phylogenetic tree are distributed within ~750 Mb of copy number differences, which is smaller than the hPT1 tree, suggesting lower genomic heterogeneity within the hPT2 population. The hPT2 P6 Clusters 1–5 (P6 C1–5) are spread within ~300 Mb and are closely related to each other. hPT2 P16 Clusters 1, 3, 4, 7, and 8 (P16 C1, C3–4, C7–8) are paired to P6 C2, while P16 C2 and C5–6 are paired to P6 C3. The estimated chromosome number ranges from 63 to 66.1.
The clusters in the hPT2 phylogenetic tree were distributed within ~750 Mb of copy number differences, which was smaller than the hPT1 tree, suggesting lower genomic heterogeneity within the hPT2 population (Fig. 3b). The hPT2 P6 Clusters 1–5 (P6 C1–5) are spread within ~300 Mb and are closely related to each other. hPT2 P16 Clusters 1, 3, 4, 7, and 8 (P16 C1, C3–4, C7–8) are paired to P6 C2, while P16 C2 and C5–6 are paired to P6 C3. The estimated chromosome number ranges from 63 to 66.1. The copy number changes associated with each edge of the phylogenetic trees can be found in Supplementary Dataset 1.
In addition to the phylogenetic tree analysis, we also performed an unsupervised hierarchical clustering analysis of the sub-population clusters of hPT1 and hPT2 (Supplementary Fig. 1), where the distance between clusters was calculated using the Canberra distance method. Indeed, the correlation between the clusters identified by the hierarchical clustering highly resembles the correlations derived from the phylogenetic tree, confirming the findings of the phylogenetic analysis.
Copy number alterations in PDAC organoids translate to transcript regulationTo investigate the functionality of these copy number alterations, the transcriptomes of hPT1 and hPT2 were analyzed through RNA-seq. The gene expression fold change between the two organoids was quantified and its profile across the whole genome was averaged at every 1 Mb. CNVs between hPT1 and hPT2 were derived from the SNPa copy number profiles that also have 1 Mb resolution. The matching resolution enables the comparison between gene expression regulation and the CNVs between the two organoids (Fig. 4a). The variation within the gene expression regulation profile is much higher than the CNV, which may be due to the trans-regulation of transcription30. However, the agreement between the CNV and gene expression regulation profile can be observed across the whole genome, especially where CNVs occur, such as in chromosomes 1, 3, 6, 7, 10, 14, 15, and 21. Indeed, such agreement was also observed in some of the frequently altered genes in PDAC28, including ARID1A, CTNNB1, RREB1, EGFR, BRAF, MYC, and SMAD4. CNVs of hPT1 and hPT2 positively correlated to the gene expression regulation between the two organoids, with a statistically significant Pearson correlation coefficient of 0.42 (Fig. 4b), suggesting “gene dosage” effects19,20,21 and functionality of chromosome copy number changes.
Fig. 4: Copy number alterations translate to transcript regulations.
a Both hPT1 P3 and hPT2 P6 organoids were subjected to RNA-seq. The gene expression fold change (log2FC, blue) between the two was averaged for every 1 Mb region across the whole genome. The plot shows good accordance between the gene expression fold change and the copy number variations (log2CNV from SNPa copy number profiles, orange) of the two organoids, especially in chromosomes 1, 3, 6, 7, 10, 14, 15, 19, and 21. The general agreements between log2FC and log2CNV are also observed for the commonly mutated genes in PDAC. b The CNV of each 1 Mb region was plotted against the corresponding gene expression fold change, resulting in a positive correlation between CNV and gene expression fold change. The correlation has a Pearson correlation coefficient (R) of 0.42, which is statistically significant, with p < 0.0001.
Gene set enrichment analysis (GSEA)31,32 of the regulated genes identified several “hallmark” gene sets33 that are significantly enriched in hPT1 and hPT2 (Supplementary Fig. 2). A couple of cell cycle-related gene sets (E2F targets and G2M checkpoint gene sets) were enriched in hPT1 (Supplementary Fig. 2A). Interestingly, SMAD4 has been shown to suppress cell proliferation through the TGF-β pathway34 and SMAD4 deficiency in the KRAS G12D PDAC mouse model promotes cell proliferation35. These previous findings suggest the possibility that the enrichment of the cell cycle-related gene sets might be driven by the copy loss and downregulation of SMAD4 in hPT1 (Figs. 1c and 4a). Moreover, the gene set that is upregulated by the activation of WNT signaling through the accumulation of CTNNB1 (WNT-beta catenin signaling gene set) is also enriched in hPT1, which might be driven by the copy gain and upregulation of CTNNB1 in hPT1 (Figs. 1c and 4a). Both of these enrichments in hPT1 suggest the functional impact of chromosome copy number changes on cellular phenotypes. Furthermore, the genes that are upregulated in hPT1 also drive the enrichment of the epithelial-mesenchymal transition gene set, metabolism-related gene sets (hypoxia, glycolysis, cholesterol homeostasis, oxidative phosphorylation, reactive oxygen species pathway, and bile acid metabolism gene sets), and the genes that are upregulated through activation of mTORC1 complex (mTORC1 signaling gene set). Meanwhile, the upregulated genes in hPT2 resulted in the enrichment of genes that are important for cell polarity (apical surface gene set), immune response-related genes (allograft rejection and complement gene sets), and genes that are downregulated by KRAS activation (KRAS signaling down gene set) (Supplementary Fig. 2b).
留言 (0)