
記住我
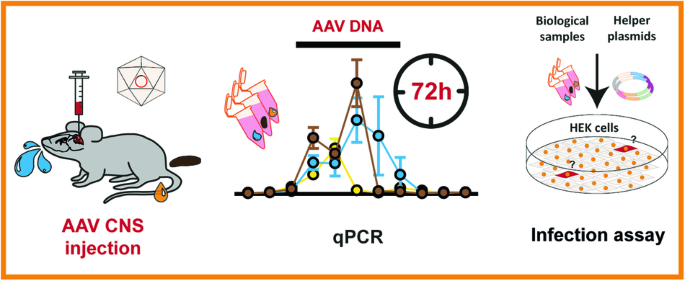
In order to generate a CAR with the same glycan-specificity we exchanged the scFV of a second generation CAR [25] with the CRD of CD301. We additionally equipped the γ-retroviral vector with an eGFP as reporter. In a similar construct, we cloned a CD301-CAR construct with a threefold Glycine-Serine-linker [(G4S)3] between the CRD and the IgG to increase the flexibility of the domain (CD301-linker). To investigate the impact of a N-terminal myc-tag in binding properties of a CRD-containing CAR we also fused a myc-tag at the 5ʹ-end of the CD301 CRD (CD301-linker-myc) (Fig. 1A + B). All viral constructs were expressed in NK92 cells to evaluate the expression and correct orientation of the CD301-CAR construct. The transduction efficiencies varied from 2.2% to 6.4%. After sorting, we performed flow cytometric analysis using an anti CD301 antibody specific for the CRD region to recognize the CAR constructs (Fig. 1C + E). All three CAR constructs were detected at the cell surface with comparable expression levels ranging from 81 to 96%, respectively. The use of an anti-myc antibody revealed a comparable expression level (96%) of the CD301-linker-myc construct as we observed with the anti-CD301 antibody (Fig. 1D). All constructs showed also stable expression after extended period of cultivation (Supplementary Fig. 1A + B)
Binding capacity of CD301-CAR constructsAfter determination of the expression of the CD301-CAR constructs, we tested the binding properties and specificity of the CRD domain fused to the CAR backbone. To do so, we made use of fluorescently labeled glycoconjugates (Tn and STn) fused to polyacrylamide (PAA) in a FACS-based binding assay (Fig. 2A). Compared to the negative controls aminoglucitol and untransduced NK92 cells (grey histograms), we observed significant binding of Tn-PAA and STn-PAA to all CD301-CARs. In addition, no binding of other glycan-structures, like Sia3-, Lewis X (Lex)- or Sialyl-Lex were detected (Fig. 2B). CD301-linker-CARs (red and blue histograms) showed an enhanced binding of Tn- and STn-PAA compared to CD301-CAR without linker (green histograms) resulting in an increase of binding from two to six fold compared to wildtype cells. STn binding was enhanced in a similar range (CD301-CAR: 2-fold, CD301-linker: 4-fold, CD301-linker-myc: 7-fold). Taken together, our data showed that the CRD of CD301 specifically binds to its glycan-structure ligands.
Fig. 2: Binding properties of CD301 CARs.
A Binding assay with fluorescently labeled PAA-glykoconjugates. NK92 cells expressing the respective CARs were incubated with biotinylated PAA-glycoconjugates premixed with Streptavidin-Alexa 647 and analyzed by flow cytometry. Binding of Tn- PAA, STn-PAA or the negative control (aminoglucitol) to NK92 cells are depicted as triplicates (WT: grey, CD301 CAR: green, CD301-linker: red, CD301-linker-myc: blue). B Evaluation of binding specificity. Binding of indicated PAA-glycoconjugates are shown as mean of fold binding compared to WT cells. Error bars showing standard deviation of triplicates. p < 0.05, p < 0.01 or p < 0.001 p < 0.0001 were indicated by *, **, *** or **** respectively.
NK92 cells expressing CD301-CARs can mediate antitumor activity against several targetsNext, we wanted to test, if our newly generated fusion constructs are able to transmit the signals for degranulation and killing after binding to target cells. In order to prove this, seven different breast cancer cell lines and one human mammary epithelial cell line (MCF10A) were screened for the expression of CD301 ligands at the cell surface by staining with a recombinant, soluble CD301 and analyzed in flow cytometry (Fig. 3A, B). High expression of Tn or STn antigens were detected on estrogen receptor positive cell lines MCF7, T47D, KPL1 and BT474, whereas the estrogen receptor negative cell lines MDA-MB-468 and MDA-MB-231(triple negative), SK-BR3 (Her2 positive),the non-tumorigenic epithelial cell line MCF10A and the myelogenous leukemia cell line K562 showed lower expression or were negative (Fig. 3B).
Fig. 3: CD301-CARs display specific cytotoxicity against breast cancer cell lines.
A Recombinant CD301 is equipped with a N-terminal myc and was fluorescently labeled with a biotinylated anti-myc antibody and Strepatividin-FITC. B Detection of CD301 ligands on target cells: breast cancer cell lines were stained with fluorescently labeled recombinant CD301 and analyzed by flow cytometry. C NK92 cells expressing the CD301-CAR, CD301-linker-CAR or CD301-linker-myc CAR were cocultured for 3 h with calcein labeled targets with an effector target ratio of 5:1 and measured as quadruplicates. D MTT assay of transduced - and WT NK92 cell. Columns represent the median of triplicates. Error bars show standard deviation. p < 0.05, p < 0.01 or p < 0.001 p < 0.0001 were indicated by *, **, *** or **** respectively.
Consequently, we evaluated the lytic activity of the NK92/CAR cells towards the different breast cancer cell lines with an E:T ratio of 5:1 (Fig. 3C). Compared to untransduced counterparts, NK92 cells expressing CD301-CARs showed increased cytotoxicity towards Tn/STn positive cell lines MCF7, T47D, KPL1 and BT474, whereas the negative cell lines or cells with low ligand expression were less effectively killed. Exceptions are the cell lines MDA-MB-231 and MCF10A, to which the CD301 construct shows a significant cytotoxicity. In a direct comparison, we see more specific cytotoxic activity of the constructs with linker sequences (and myc-tag) and improved correlation of killing efficiency and ligand expression (Supplementary Fig. 3). As NK92 cells exhibit killing activity against K562 cell line lacking the MHC complex required to inhibit NK activity, we compared the natural cytotoxicity of the wildtype and the transduced populations. NK92 WT and NK92 CAR cells disclosed a relevant lysis of K562 cells indicating that the transduction and selection procedures do not affect the cytotoxic properties of NK92 cells.
Parallel to cytoxicity measurements we assessed the cell metabolic activity as a marker for vitality of the effector cells in MTT assay. We detected no differences between the different NK92 cell lines (Fig. 3D). Thus, the results obtained from cytotoxicity assays indicated a specific killing of the CD301-CARs constructs.
The secretion of lytic granules results in surface exposure of the lysosomal-associated protein CD107a. As shown in Fig. 4, we analyzed the percentage of CD107a positive NK92 cells after incubation with different target cell lines.
Fig. 4: CAR expression leads to enhanced degranulation of NK92 cells upon engagement with CD301 ligand-positive breast cancer cells.
A Degranulation of CD56+ NK92 CAR cells was analyzed by flow cytometry assessment of CD107a surface expression after 4 h of co-culture with MCF7, T47D, MDA-MB-468 or K562 cells (CD56-) (E:T 1:1). Parental NK92 cells were included for comparison. Unstimulated effector cells or stimulated with PMA/ionomycin served as basal and positive controls, respectively. B Evaluation of degranulation assay. Columns represent mean percentage of CD107a positive NK92 cells measured in triplicates. Error bars show standard deviation. p < 0.05, p < 0.01 or p < 0.001 p < 0.0001 were indicated by *, **, *** or **** respectively.
By using Tn/STn -expressing breast cancer cell lines T47D and MCF7, NK92 CAR cells underwent a strong upregulation of CD107a expression that was very limited in wildtype NK92 cells. Although the percentage of GFP-positive NK92 cells were comparable (Supplementary Fig. 5A), the degranulation levels in the presence of T47D cells varied between the CAR constructs. Linker CARs led to a more effective degranulation compared to CD301-CAR without linker, presumably due to linker-dependent improved accessibility of the target structures.
Importantly, the incubation with ligand-negative MDA-MB-468 cells only led to a slight increase of CD107a positive cells demonstrating the specificity of the newly generated CAR constructs.
PMA/ionomycin treatment was used as a control for maximal degranulation. For measuring CAR-independent degranulation, NK92 cells were incubated with K562 cells. In contrast to the other NK92 cell lines this non-specific stimulus produced only marginal effects in NK92 cells expressing CD301-linker-myc CAR, although these cells still show enhanced degranulation towards MCF7 and T47D cells.
A common feature of antigen receptors like T cell receptor and synthetic CARs is ligand-induced downmodulation considering a hallmark of lymphocyte activation [27]. Therefore, we investigated surface expression of CD301-CARs after incubation with target cells for 3- or 24 h, respectively, PMA treatment was included as a control for activation. To investigate the impact of degradation by proteases (sheddases), we applied in parallel TNF protease inhibitor 2 (TAPI-2), a broad-spectrum inhibitor of sheddases.TAPI-2 inhibits PMA-induced shedding of various cell surface proteins [28,29,30].
We observed downregulation of 40–65% of surface CD301-CARs on NK92 cells incubated with CD301 ligand positive target cells after 3 h. The strongest effect was measureable after incubation of linker CD301-CAR expressing NK92 cells with T47D, which was not as pronounced in NK92 cells expressing CD301-CAR without linker sequence. MDA-MB-468 cells providing only low levels of CD301 ligands and induced only slight downregulation of the linker CD301-CARs. Compared to NK92 cells treated with PMA alone, the addition of TAPI-2 led to an increase of ~5%. After 24 h, a partial recovery of the surface CD301-linker-CARs was detectable, while the amount of CD301-CARS without linker remained reduced. To reveal, if CD301 domains are cleaved from the CAR constructs, we performed competing binding assays using the supernatants after 24 h co-culture in combination with fluorescently labeled recombinant CD301 (Supplementary Fig. 4). We measured only slight inhibitory effects of maximal 10% reduction of binding to MCF7 cells. These results lead us to the assumption, that the major part of down-regulation is not due to degradation by sheddases.
CD301-CARs are selectively activated by target structures on cancer cells resulting in a different extent of activationTo further confirm that the increased killing of NK92 CAR cells was strictly Tn/STn-specific and could be ascribed only to CAR activation, we analyzed cytotoxicity using different E:T ratios. NK92 CD301-CAR cells showed, even at low E:T ratios, an extremely high cytotoxicity to Tn/STn-positive MCF7 and T47D cells, which were not killed by wildtype NK92 cells. The differences between the CAR constructs were only marginal within a range of 20%. CD301-linker-CAR turned out to be the most effective construct (Fig. 6A). These tendencies was also observed after long term cultivation of NK92 cells indicating robust performance of the NK92 CAR cells (Supplementary Fig. 1C).
Likewise, T47D cells were also selectively killed by NK92 CAR cells but not by the wildtype NK92 counterparts (Fig. 5A). Again, CD301-linker-CAR showed the highest lytic activity. In contrast NK92 expressing CD301-CAR or CD301-linker-myc-CAR mediated significantly lower killing of T47D cells. As further proof of specificity, both NK92 CAR and NK92 WT cells failed to lyse efficiently Tn/STn-negative MDA-MB-468, MCF10A cells and three different neuroblastoma cell lines (Supplementary Fig. 6).
Fig. 5: Ligand induced downregulation of CAR constructs.
Surface expression of CD301 CARs were analysed after 3 h or after 24 h of co-culture with MCF7, T47D, MDA-MB-46. PMA treated cells served as postiv control for activation. In parallel 50 µM TAPI-2, an inhibitor for MMPs and TACE, was added to PMA treated cells for 3 h. Error bars show standard deviation. Significance levels were calculated in relation to non-treated cells (n.t.) p < 0.05, p < 0.01 or p < 0.001 p < 0.0001 were indicated by *, **, *** or **** respectively.
Additionally, we tested IFN-γ release by NK92 CAR and NK92 cells following target engagement. Results of this assay fully mirrored those already obtained with CD107a expression analysis shown in Fig. 4, as PMA/ionomycin massively, but non-specifically, stimulated either effector populations. MDA-MB-468 and MCF10A cells were negligibly recognized, while CAR expressing NK92 cells produced significant levels of IFN-γ upon interaction with CD301 ligand positive cell lines MCF7 and T47D (Fig. 6B). Again, CD301-linker-CAR induces the highest IFN-γ secretion especially in the presence of T47D.
Fig. 6: Lytic activity of CD301-CAR expressing NK92 cells.
A Cytotoxicity of NK92 cell expressing CD301-CAR (green), CD301-linker-CAR (red) or CD301-linker-myc CAR (blue) in comparison to wildtype NK92 cells (grey) was investigated against MCF7, T47D, MDA-MB-468 cells and MCF10A, at different E:T ratios. B IFN-γ release was analyzed in supernatants of CAR expressing NK92 CAR or wildtype NK92 cells stimulated with MCF7, T47D, MDA-MB-468 cells and MCF10A (E:T 1:1), respectively. Untreated effector cells and effector cells treated with PMA/ionomycin served as controls. Results are reported as mean values ± SD of triplicates. p < 0.05, p < 0.01 or p < 0.001 p < 0.0001 were indicated by *, **, *** or **** respectively.
The data indicates that the lectin based CD301-CAR is fully functional and confers glycan-selective directed and enhanced activity.
留言 (0)