
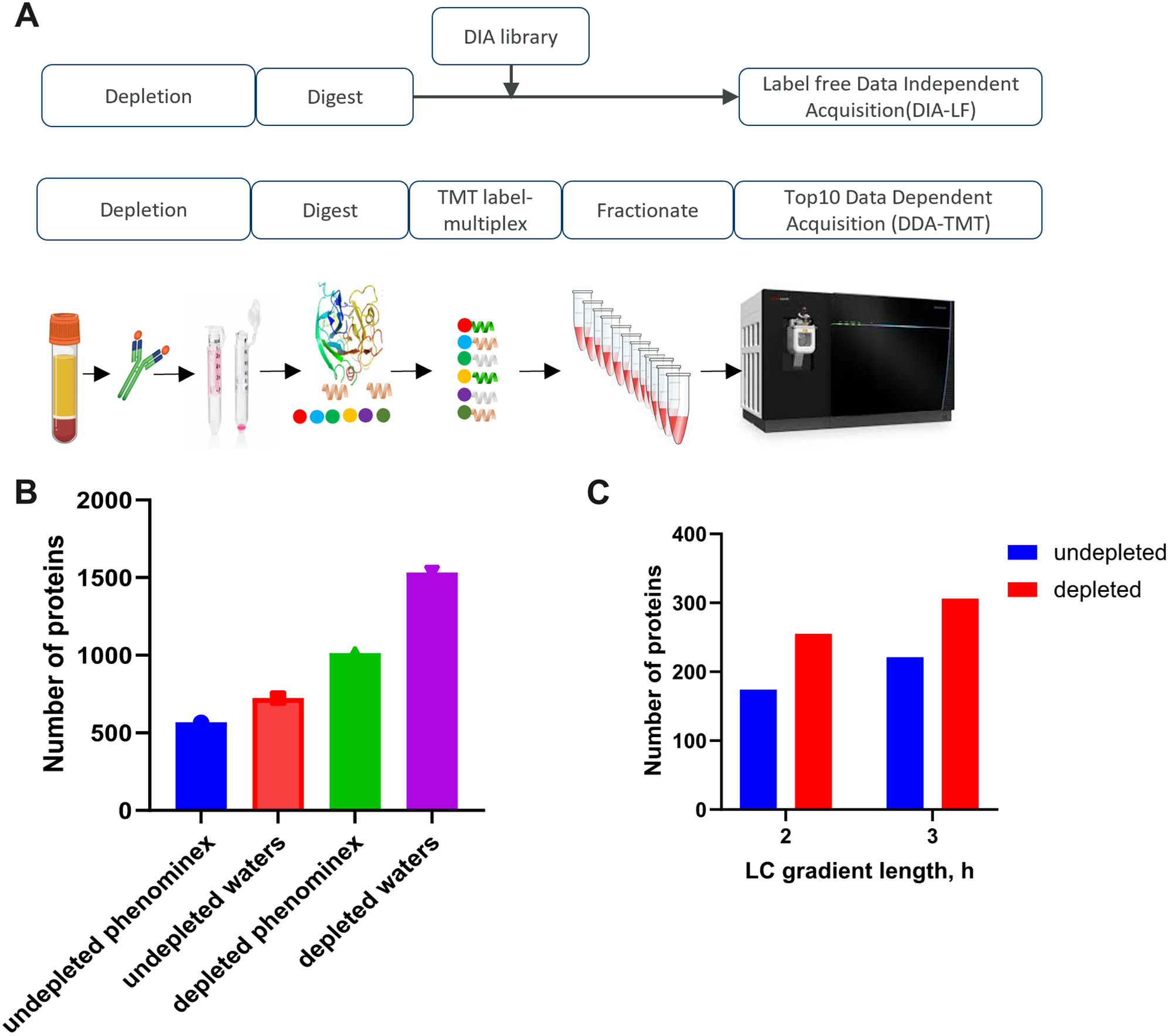
記住我



The COVID-19 cohort consisted of 3 males and 3 females with an age range from 54 to 81 (median 79.5 years). Time from first positive SARS-CoV-2 (ante mortem) PCR to death ranged from 2 to 34 days (median: 9 days) and the post-mortem interval ranged from 7 to 57 h (median 18.5 h). Details about patient metadata are provided in Table 1 and (Additional file 2: Table S1). For both COVID-19 patients and controls, two individual lung tissue pieces were collected at the point of autopsy. Tissue pieces originating from the same patient (Additional file 1: Fig. S1A) were treated as replicates, were processed as described in the materials and methods section and then subjected to comprehensive proteomics analysis. The resulting protein list containing the mean quantitation data from two technical replicates (or the single quantitative value if observed in only one technical replicate) was then filtered to maintain only those proteins with reported values in at least four samples of each group (COVID-19 and control).
This produced a matrix of 3431 proteins which was then subjected to statistical analysis using a linear model approach [16]. All 3431 proteins including the fold changes (COVID-19 versus control) and statistics are listed in (Additional file 3: Table S2). At all times, a minimum alpha level of 0.05 and a minimum fold-change of 1.5 was maintained and borderline significant proteins with low fold-change were excluded from the analysis, as indicated by the dynamic cut-off (Fig. 1A). This approach resulted in identification of 239 significant differentially abundant proteins between COVID-19 and control lung samples (Fig. 1A). The significantly more abundant proteins in COVID-19 samples (dynamic p-value cut off < 0.05 and fold change to control > 1.5) were then used for enrichment analysis of either biological processes (using String database and Cytoscape for visualization; Fig. 1B) or metabolic and signalling pathways (using Reactome pathway analysis; Fig. 1C).
Fig. 1
The most upregulated proteins in lung tissues of terminal COVID-19 patients are involved in immune and stress response, extracellular matrix remodelling, as well as clot formation and fibrinolysis. A Volcano plot of the LFQ diaPASEF dataset after limma analysis (limma dynamic p-value < 0.05) including box dot plots of the most prominently changed proteins. B Grouping of selected significantly enriched biological processes (limma dynamic p-value < 0.05) based on similarity after gene ontology enrichment analysis using String and Enrichment map in Cytoscape with significantly more abundant proteins in COVID-19 (limma dynamic p-value < 0.05, fold change to control > 1.5) as input. C Selected significantly enriched Reactome pathways (enrichment FDR corrected p-value < 0.05) with the same input protein list (as for B)
Among the most prominently upregulated proteins in COVID-19 lungs were members of the complement activation cascade (e.g. C4, C3, C5, B, C2, H, C9), different interferon-induced proteins (e.g. Interferon-induced GTP-binding protein Mx1 (MX1)) as well as inter-alpha-trypsin inhibitor family (ITIH) proteins, known acute phase proteins that are upregulated during inflammation [18]. Also other inflammatory proteins were more abundant in COVID-19 lungs, including different isoforms of alpha-1-acid glycoprotein [19] as well as vitamin D binding protein (GC), which not only transports vitamin D but also acts as immune system activator [20, 21]. In addition, we report a prominent upregulation of proteins active in blood clot formation, including prothrombin (F2), coagulation factor XII (F12), antithrombin-III (SERPINC1), plasminogen (PLG) and others (Additional file 3: Table S2).
Correspondingly, both String GO and Reactome pathway analyses resulted in the enrichment of the same major processes/pathways, including fibrin clot formation, platelet activation, immune activation as well as cytokine, interleukin and interferon signalling (Fig. 1B, C). In addition, several regulators of the extracellular matrix (ECM) organization and structure seemed to be affected by SARS-CoV-2 infection (Fig. 1B, C). Extracellular proteins which are higher in the COVID-19 group were mainly involved in processes of clot formation (fibrinogens and plasminogens), lipid transport (apolipoproteins) as well as de-novo collagen synthesis (Serpin H1 and prolyl 4-hydroxylase subunit alpha-1; Additional files 3, 4). However, while these extracellular proteins were higher expressed in COVID-19 patients, expression of other ECM proteins appeared to be completely abolished in the terminal infected lungs, as will be discussed in the following subchapter. The list of all enriched biological processes and pathways (with proteins significantly more abundant in COVID-19 samples as input (limma dynamic p-value < 0.05)) can be found in (Additional file 4: Table S3).
Expression of prominent ECM constituents and regulators is diminished in COVID-19 affected lungsInterestingly, it seems that while SARS-CoV-2 infected lungs cope with a cytokine storm, complement system activation and increased clot formation (Figs. 1B, C and 2B), they also undergo severe ECM restructuring. Correspondingly, quantitative proteomic profiles of the infected lungs revealed reduced abundance of a number of different ECM proteins, including constituents of the basement membrane as well as different types of collagen and fibrillin (Fig. 2A, Additional file 3: Table S2). In line with this, Ingenuity Pathway Analysis (IPA) of significantly altered proteins resulted in the enrichment of cell migration and cell movement processes, which are known to be associated with structural changes of the ECM (Fig. 2B).
Fig. 2
The most prominently downregulated proteins in COVID-19 patient lung tissue mainly belong to the ECM constituent and fibril/filament formation. A String protein interaction network of significantly downregulated proteins in COVID-19 patients (dynamic p-value < 0.05; interaction confidence level 0.7 (high)); B Data from String was further corroborated with IPA predicted “activated” or “deactivated” significantly enriched diseases and functions in COVID-19 patients with significantly altered proteins (p-value < 0.05) as input. Higher Z-score represents predicted increase of the given pathway/function in COVID-19 patients while lower Z-score predicts a decrease (Z-Score cut-off ± 1.5)
Proteomics findings were further corroborated by histopathological analysis after staining using the pentachrome method, which allows for simultaneous staining of both collagen (yellow) and sulfated mucopolysaccharides (light blue), in addition to nuclei (black), muscle (red) and elastin (purple) [22]. As visible on Fig. 3A, COVID-19 affected lungs have almost no residual yellow colouring, suggesting massive loss of collagen structure. In addition, structural organisation of elastic fibres also seems deregulated in diseased lungs.
Fig. 3
Histopathology of COVID-19 patient lung tissue reveals loss of extracellular matrix and prominent infiltration of immune cells. A Movat pentachrome stain of lung tissue slices from COVID-19 patients. a–e 20 × images, b 10 × overview. Whilst overall tissue structure is still well defined, none stained positive for collagen and reticular fibres. B a 10 × overview, b–f 20 × images of healthy lung tissue controls. Except f all of the healthy controls display a strong positive staining result (brownish yellow) for collagen and reticular fibres. The reduced signal in f could be explained by an incidental focal inflammation. C a–f 20 × images of H&E stains of COVID-19 samples. Samples display a varying amount of infiltrated immune cells, from minor, early onset inflammation (a, b, f) to advanced inflammation (d, e) to alveoli filled with infiltrate (c). D a–f 20 × images H&E stains of healthy control patients display clinically unremarkable lung parenchyma. Displayed images are representatives of 10 pictures taken per each of the six COVID-19 and control patient analysed in this study
In addition to reduced collagen and fibril expression, our COVID-19 lungs also display decreased abundance of a number of different myosin and tropomyosin isoforms, as well as laminins, which are critical components of the basement membrane. Viral penetration in lungs is known to cause cytoskeleton rearrangement [23] and consequent infection can lead to the disruption of alveolar-capillary barrier, leading to lung injury and reduced gas flow [24, 25]. One of the key protein groups responsible for maintaining the integrity of the barrier (through formation of tight junctions) are cadherins, especially E-cadherin (Cadherin-1) [24]. Correspondingly, in our dataset we observe a trend towards lower expression of a number of cadherin isoforms, including cadherin-1, 5, 13 as well protocadherin-1 (Additional file 1: Fig. S1 and Additional file 3: Table S2, respectively), suggesting a tight-junction breach in the lungs infected by SARS-CoV-2. As a result of such injury, structural organization of the underlying laminin-rich basement membrane can occur and recent reports indeed describe the loss of laminin expression and disruption of the laminin structural arrangements in lungs of COVID-19 patients [26]. Our study further corroborates these findings, as next to reduced abundance of cadherins, laminins seem to be diminished in lungs of COVID-19 patients (Fig. 2; Additional file 3: Table S2).
Lastly, lungs of terminally ill COVID-19 patients also display a prominent reduction in the expression of several members of the major histocompatibility complex (MHC) class II, including HLA-DPA1, HLA-DRB1 and HLA-DRB5, all of which are reported to be reduced in antigen presenting cells of critically ill COVID-19 patients [27].
Signalling is prominently affected as a consequence of SARS-CoV-2 infectionAs expected upon viral infection and according to the IPA analysis of canonical pathways, COVID-19 affected lungs show a strong activation of interferon and acute phase response signalling (Fig. 4A), accompanied by the higher expression of a number of interferon-induced proteins (Additional files 3 and 5) as well as signal transducers and activators of transcription (STATs; Additional files 3 and 5, Fig. 4B), known acute inflammatory responders [28].
Fig. 4
Lungs of COVID-19 patients depict pronounced changes in global, kinase mediated signalling. A IPA analysis of canonical pathways with significantly altered proteins before multi-testing correction (p-value < 0.05) as input revealed prediction of lower activation of a number of different, major kinase signalling pathways. Higher Z-score (marked in yellow) predicted activation, while lower Z-score (purple colour) represents prediction of a lower activation of a given pathway in lungs of COVID-19 patients. FDR control on pathway level (enrichment FDR corrected p-value < 0.05). B Lungs of COVID-19 patients show reduced abundance of several key kinases but prominent activation of STATs, typical mediators of inflammation (*Student’s t-test p-value < 0.05, ** Student’s t-test p-value < 0.01)
In addition, severe SARS-CoV-2 infection seems to induce host’s tumor suppressor phosphatase and tensin homolog (PTEN) signalling (Fig. 4A). PTEN is a known antagonist of PI3K/AKT axis and an upstream negative regulator of mammalian target of rapamycin (mTOR) signalling [29], both of which were predicted to be downregulated in our dataset (Fig. 4A). Such an overexpression of PTEN followed by concomitant inhibition of key cellular responsive pathways can be detrimental for the antiviral response, as it leads to suppression of antibody production and consequently a worse outcome [30].
However, not only downstream targets of PTEN are affected by SARS-CoV-2 infection. A number of signalling pathways was predicted as deactivated in COVID-19 lungs (Fig. 4A), including other major kinases responsible for cellular proliferation and cell cycle progression (Fig. 4A, B).
These findings are in line with a recent large time-course phosphoproteomics study of SARS-CoV-2 infection in Vero E6 cells (cells highly susceptible to SARS-CoV-2 infection), which described that the viral infection promotes the host’s p38-MAPK cascade while shutting down key mitotic kinases, including phosphoinositide-3-kinase (PI3K), RAC-alpha serine/threonine-protein kinase 1 and 2 (AKT/2), cAMP-dependent protein kinase (PRKACA/B), Rho-associated protein kinase (ROCK1/2) and others [31]. It seems that together with the overexpression of PTEN, SARS-CoV-2 infection indeed leads to a full proliferation halt in the hosts’ lungs.
Furthermore, interleukin signalling is also predicted to be decreased in lungs of terminal COVID-19 cases (Fig. 4A). Although IL-6 has been reported to be increased in plasma of COVID-19 patients [32, 33] and higher neutrophilic IL-8 expression was observed in severe COVID-19 cases [34], we did not detect such trends with our proteomics approach locally in the lung tissue post mortem. However, pre-mortem blood draw of the six COVID cases in our study showed elevated IL-6 serum levels [9]. Overall observed deactivation of various interleukin pathways in COVID-19 patients might be partially due to anti-IL-6 treatment (one patient) and/or potential glucocorticoid treatment (at least two COVID-19 patients received prednisolone while at the ICU). Glucocorticoids (including dexamethasone) are known to supress interleukin signalling [35,36,37].
Lastly, insulin signalling is predicted to be activated in the lungs of COVID-19 patients (Fig. 4A). This is not surprising and can almost act as a positive control of this study, as four out of six COVID-19 patients (70%) were diabetic, but none of the controls (Additional file 2: Table S1). Correspondingly, glucose metabolic disorder was the most probable predicted disease in the IPA analysis (see Fig. 2B).
Histopathological stratification of COVID-19 cases is reflected by distinct proteomics signaturesOut of the six end-stage COVID-19 patients involved in this study, for three (50%) the pathologically ascribed cause of death was bacterial (secondary) pneumonia (Table 1 and Additional file 2), while for the other three it was diffuse alveolar damage (DAD; in one patient DAD was combined with fungal pneumonia; Table 1 and (Additional file 2: Table S1). Interestingly, despite the low sample number, the proteomic profiles reflect these histopathological differences, as can be seen in the principal component analysis (Fig. 5A) as well as in the volcano plot (Fig. 5B; significantly changed proteins before multi-testing correction marked in black; p-value < 0.05).
Fig. 5
Proteomic profile of lung tissue from COVID-19 patients differs based on the cause of death: pneumonia or DAD/ARDS. A Principal component analysis of the COVID-19 samples demonstrates that samples cluster depending on whether the patients died of pneumonia or DAD/ARDS; B proteomic profiles correspond to the cause of death, with major bacterial infection response proteins upregulated in pneumonia samples and not in DAD/ARDS. C Gene ontology analysis of biological processes (GOBP) of significantly more abundant proteins in the pneumonia cohort (p-value < 0.05, log2 fold change (to DAD/ARDS) > 0.5). D GOBP of significantly more abundant proteins in the DAD/ARDS cohort (p-value < 0.05, log2 fold change (to pneumonia) > 0.5)
Among the proteins upregulated in the pneumonia cohort were mainly proteins involved for leucocyte and neutrophil activation and general response to bacterial infection (e.g. lysozyme C (LYZ), bactericidal permeability-increasing protein (BPI), peptidoglycan recognition protein 1 (PGLYRP1) and others), as well as oxidative stress defence (e.g. catalase (CAT), peroxiredoxin-1 (PRDX1), Myeloperoxidase (MPO); (Additional file 6: Table S5). This was also corroborated by GOBP enrichment analysis of the significantly more abundant proteins in the pneumonia cohort as input (p-value < 0.05, log2 fold change (compared to DAD) > 0.5; Fig. 5C). In addition, in the pneumonia cohort also a wound repair response can be observed (Fig. 5C). However, as expected, more prominent ECM and basement membrane reorganization was detected in the DAD group (Fig. 5D). In addition, cytoskeleton organization as well as cellular respiration was prominently affected in DAD samples. A complete list of enriched GOBP terms is included in (Additional file 7: Table S6).
留言 (0)