
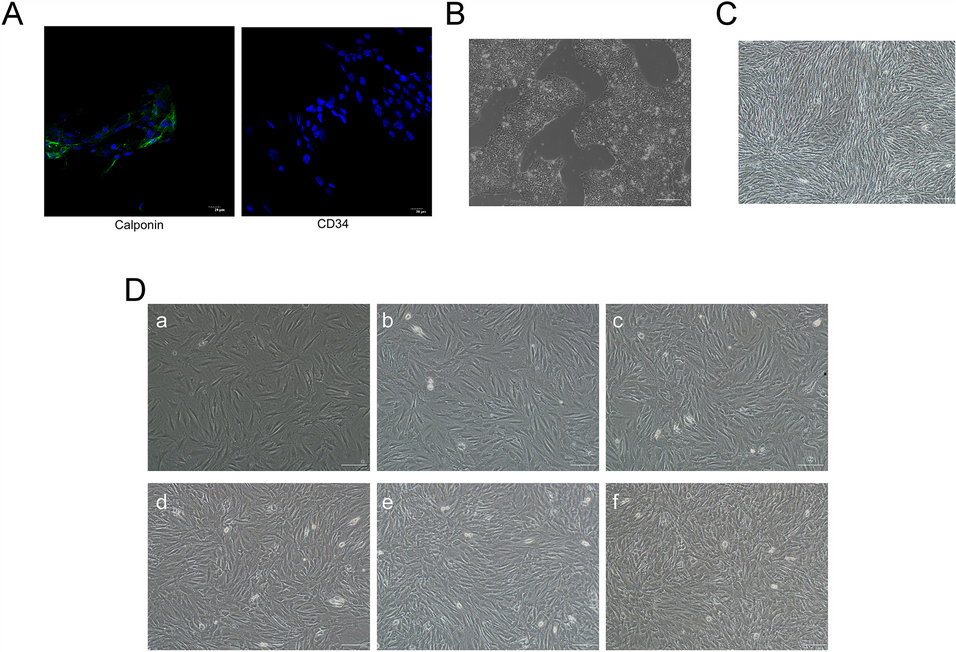
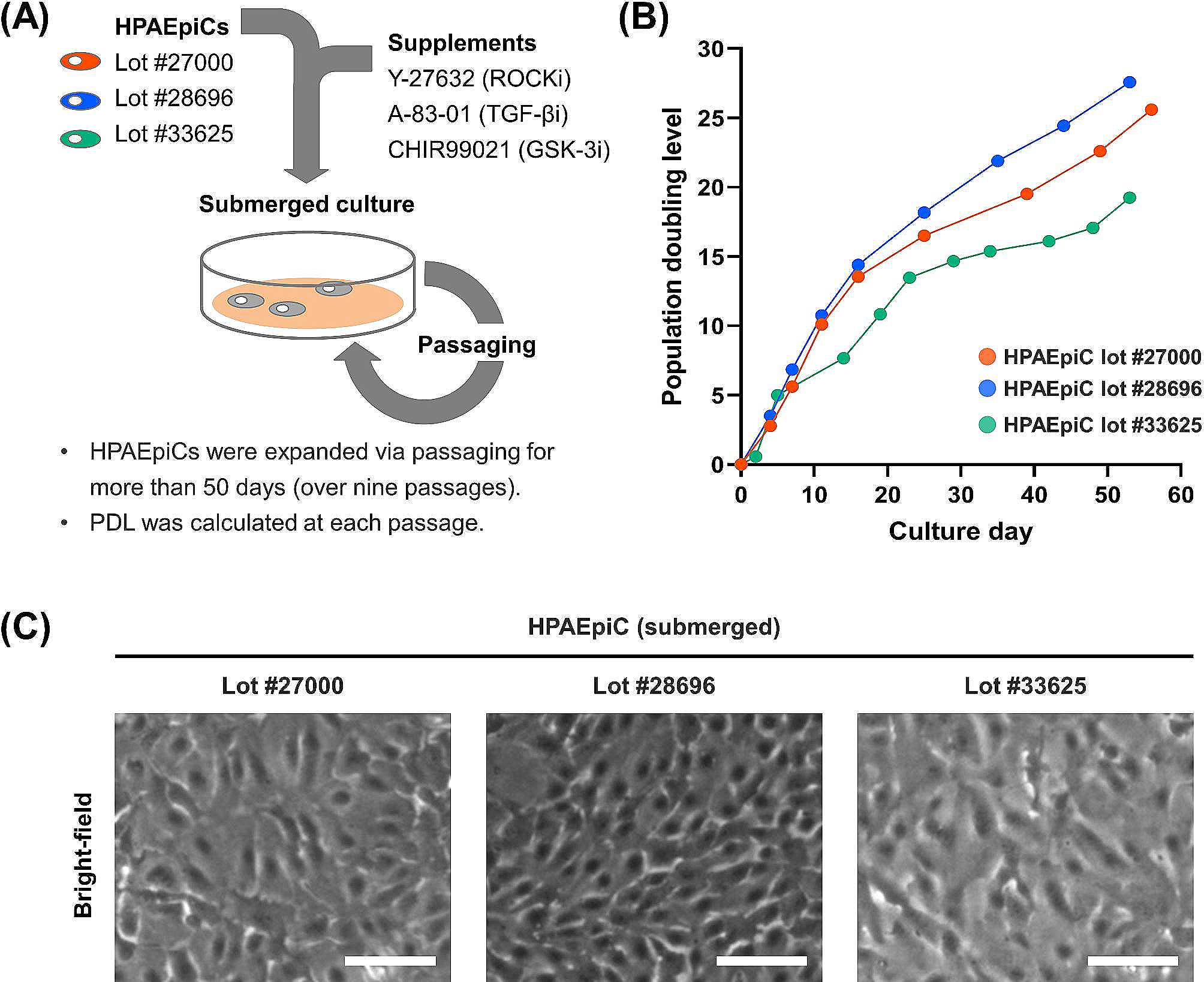
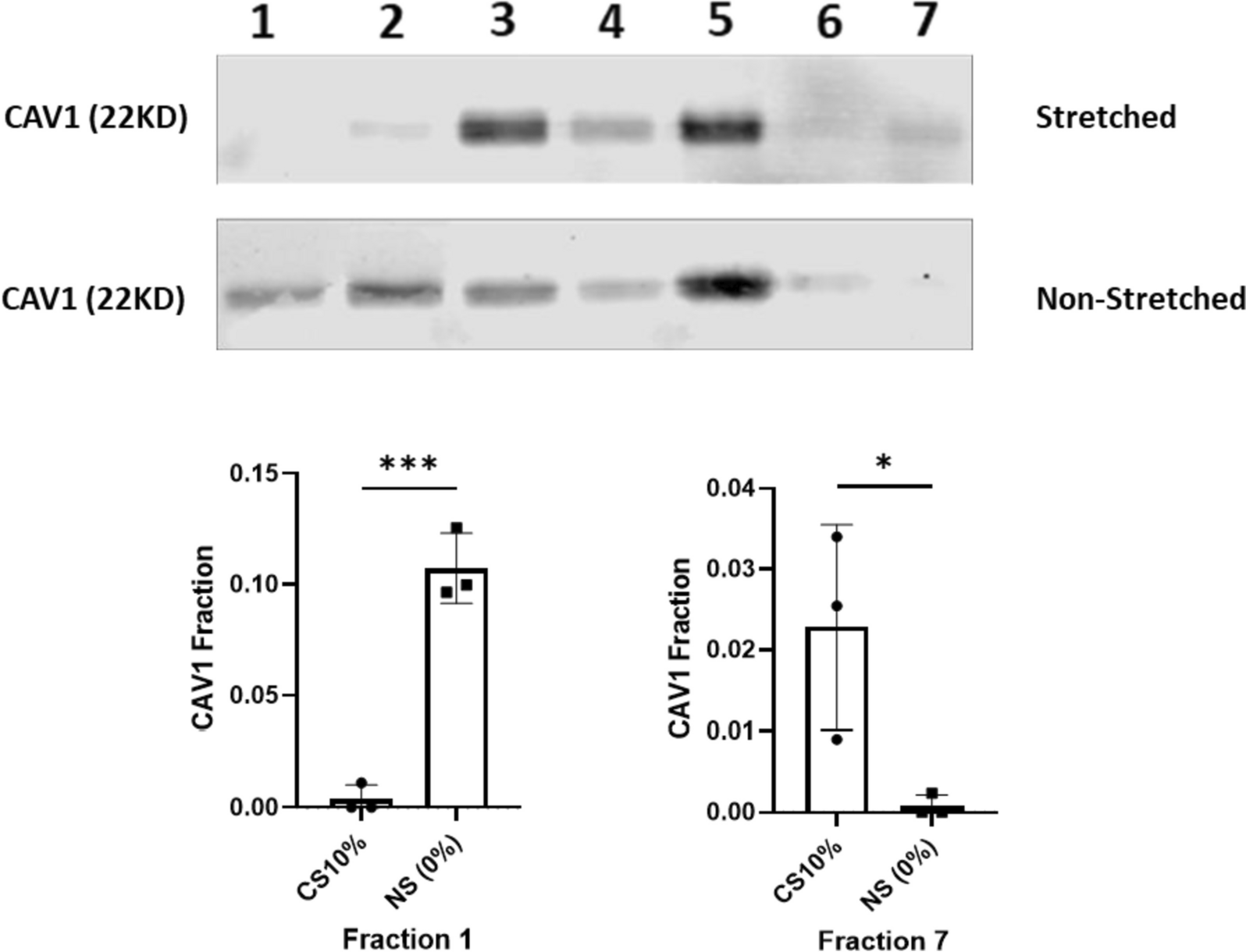
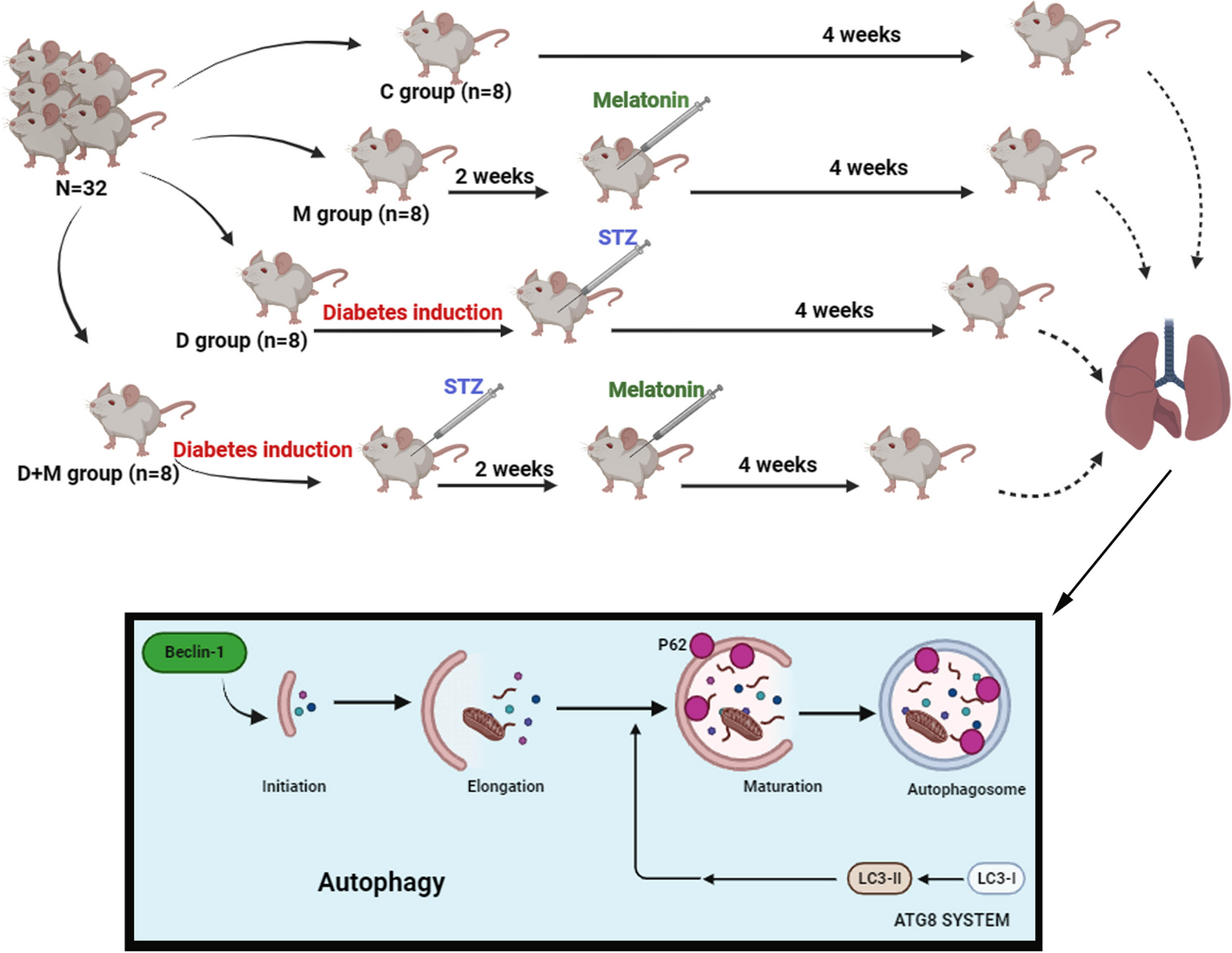
記住我


To reveal the flowering time and phenotypic changes of M. liliiflora ‘Hongyuanbao’, the entire flower bud differentiation process was observed by paraffin sections at regular intervals. Many Magnolia bloom once in either spring or summer, after completing the flower bud differentiation process during the previous year. Conversely, we found that ‘Houngyuanbao’ completed the differentiation process twice each year and flowered in both spring and summer (Fig. 1A). Based on the morphological features of floral bud differentiation in ‘Hongyuanbao’, we divided the process into the following six stages (also represented by different colors) (Fig. 1B): undifferentiated, early flower bud differentiation, sepal primordium differentiation, petal primordium differentiation, stamen primordium differentiation, and pistil primordium differentiation. Plant phenotype observation revealed that the spring flowers, which had completed flower bud differentiation in the previous year, were mainly produced at the top of the last year branches. As the falling of the spring flowers, new branches grown, together with the first time progress of bud differentiation on top the branches from April 22 to May 30. Then the first time flower buds turned to be bigger and bloomed on June (summer flower), as the second flower bud differentiation would begin and finished at the end of August. The flower bud of the second time would bloom in the next spring. (Fig. 1). This phenomenon appeared that variation existed in the development and opening process of flower buds between spring and summer, which possibly be attributed to variation in plant nutritional status and environmental conditions [9, 29].
Fig. 1
Observation of flower buds and plant phenotypes in Magnolia liliiflora ‘Hongyuanbao’. A The characteristic of twice flowering of ‘Hongyuanbao’. White bars, 2 mm; Red bars, 50 μm. B The time course of flower bud differentiation. Red bars, 50 μm. C The plant phenotypes changes of ‘Hongyuanbao’ during twice flowering process. S1, S2, S3 represent the early, middle, and later stage of the first flower bud differentiation, respectively. S4, S5, S6 represent the early, middle, and later stage of the second flower bud differentiation, respectively. The arrows pointed to the bud in April represent to the first flower bud differentiation process and the arrows in June represent to the second flower bud differentiation process
Untargeted metabolomic comparison of primary metabolites across the first and second flower bud differentiationTo explore the mechanisms of the twice flowering trait, we harvested buds in April and June with early stage (undifferentiated, S1 & S4), middle stage (sepal primordium and petal primordium differentiation, S2 & S5), and late stage (stamen primordium and pistil primordium differentiation, S3 & S6) (Fig. 1). The primary metabolites of the two bud differentiation periods were analyzed using gas chromatography-mass spectrometry (GC-MS). In most metabolomic data analyses, confounding factors orthogonal to the variables of interest may obscure the intended class separation. To analyze these data, we compared two different analyses: principal components analysis (PCA) and orthogonal projection to latent structures discriminant analysis (OPLS-DA). For PCA, our classification variables was obscured, and the variables were dispersed across several principal components. Therefore, better visualization and subsequent analysis could not be carried out (Additional file 1). Thus, we tested OPLS-DA as an alternative method, which could filter out the dis-related orthogonal variables in classification variables. We were then able to analyze both the nonorthogonal and orthogonal variables (Fig. 2). The value of the Q2 is deemed to represent the prediction ability of OPLS-DA model, and for biological samples, Q2 ≥ 0.4 is ideal [30]. It showed that the OPLS-DA score plot for each comparison between the primary and secondary flower buds had overall cross-validation coefficients, and Q2(y) was 58, 78 and 77% respectively. It suggested that OPLS-DA model was more suitable than PCA for sample category prediction of ‘Hongyuanbao’. The coordinates value for to1 and t1 showed a clear separation of the first predicted component between the two groups (Fig. 2A-C), showing the inter-group and intra-group differences.
Fig. 2
Comparisons of metabolite profiles in the first and second flower buds. A-C orthogonal projection to latent structures discriminant analysis (OPLS-DA) score plot for the different stages, comparing metabolite between the first and secondlower buds. n = 5. t1 represents the score values of the main components and reveals the differences between groups; to1 displays the score value of the orthogonal component and the difference within the group; Q2 represented the prediction ability of OPLS-DA model. D-F Heatmap of the classification in the different stage comparing between the first and second flower bud differentiations on metabolite profiles. S1, S2, S3 represent the early, middle, and later stage of the first flower bud differentiation, respectively. S4, S5, S6 represent the early, middle, and later stage of the second flower bud differentiation, respectively
To investigate the relationship between metabolites and the twice flowering phenotype, the metabolic profiles of both the primary and the secondary flower buds were used to conduct hierarchical cluster analysis (Fig. 2). A total of 41 differential metabolites were identified (Additional file 2). Based on the Mass Bank and Kyoto Encyclopedia of Genes and Genomes (KEGG) and LECO-Fiehn Rtx5 Database analyses [31, 32], all 41 metabolites were analyzed and divided into six groups: Carboxylic acids and derivatives (including 12 compounds), Organooxygen compounds (13), Fatty Acyls (5), Prenol lipids (3), Indoles and derivatives (2), and other compounds (6). Compared to first flower bud differentiation, the second flower bud showed higher levels of most sugars and organic acids, while the levels of glutamate, pyroglutamate, malate, citrate and 2-oxoglutarate were decreased. It revealed that in second flower buds, sugars including D-Talose 1 and 6-deoxy-D-glucose 1 were increased at the early stage and the levels of sucrose, trehalose, galactose 2, xylose 1 and some other sugars at the middle stage were higher. The second flower buds had higher levels of certain sugars than first flower buds at the later stage, including trehalose, erythrose 2, and d-glucoheptose 1. The data suggested that many metabolites, especially sugars, are necessary for the differentiation of the second flower bud.
Correlation among metabolites in M. liliiflora ‘Hongyuanbao’To study the correlation between the metabolites in different flower bud differentiation stages of M. liliiflora ‘Hongyuanbao’, Pearson correlations were used to calculate the degree of similarity within metabolite profiles in the three different developmental stages. MetaboAnalyst 4.0 (http://www.metaboanalyst.ca) [33] was used to map the correlation network among metabolites. S1, S2, S3 and S4, S5, S6 represented the early (undifferentiation), middle (sepal differentiation and petal differentiation), and later (stamen differentiation and pistil differentiation) stages of the first and second bud differentiation respectively. It revealed that between the first and second flower buds, 22, 41, and 25 metabolites differed across the early, middle, and later stages (Fig. 3A-C). In order to screen key differential metabolites and metabolic pathways by analyzing the differential metabolite interactions at each flower bud differentiation stage, correlation networks were built. These networks showed that in the differential metabolites at the early stage, Methylmalonic acid, 4-hydroxycinnamic acid, 4-Hydroxybenzoic acid and Hydroxypyruvic acid interacted the most closely with other differential metabolites. Sucrose, Oxalic acid and Glutamic acid were found to interact the most closely with other differential metabolites at the middle stage. Glutaric acid, Palmitic acid, Myristic acid, Trehalose and Linolenic acid interacted the most closely with other differential metabolites at the later stage (Fig. 3). Besides that, it was found that the activity and association of metabolites were more prominent during the comparison of S2 and S5 than the comparison of S1 and S4, or S3 and S6 (Fig. 3). The results suggested that among those differential metabolites, the sugar metabolites were closely associated with other metabolites, and the sugar metabolism pathway might play a vital role during the first and second flower bud differentiation processes.
Fig. 3
Metabolite-Metabolite Interaction Network in different stages. S1, S2, S3 represent the early, middle, and later stage of the first flower bud differentiation, respectively. S4, S5, S6 represent the early, middle, and later stage of the second flower bud differentiation, respectively. The circles of different sizes represent interactions between metabolites. The larger the circle, the more closely it interacts with other metabolites. The straight lines between the circles represent interactions between metabolites
KEGG enrichment analysis of sugar metabolites and the expression of MlTPS genesThe results of KEGG enrichment analysis showed that sucrose and trehalose in the sucrose and starch metabolic pathways were significantly up-regulated in the second flower buds than the first flower buds (Additional file 3, Fig. 4A). T6P is a signal of sucrose availability and is synthesized from UDP-glucose and glucose-6-phosphate by TPS genes [18, 21]. But only five TPS genes were found in transcriptome from the bud mixtures during twice flowering process, which named as MlTPS1, MlTPS5, MlTPS6, MlTPS7 and MlTPS9 respectively. The expression levels of MlTPS genes in twice flowering process were then analyzed by qRT-PCR. It showed that the expression levels of MlTPS genes had significantly differences during the twice differentiation process. Compared to the first flower bud differentiation, MlTPS1, MlTPS7 and MlTPS9 were the most prominent in expression, and MlTPS5 was barely discernible in the second flower bud differentiation process (Fig. 4B). The overall trends demonstrated that the expression of MlTPS genes were increased significantly at the middle stage in the second flower bud differentiation (Fig. 4), suggestive of potential function role of MlTPS genes in the second flower bud differentiation.
Fig. 4
Important metabolite pathways during the first and second flower bud differentiations. A Enrichment of the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway. Relatively high numbers of metabolites were annotated as starch and sucrose metabolism with rich factor. B The levels of expression of MlTPS genes in the three stages of the first and second flower bud differentiation processes in Magnolia liliiflora ‘Hongyuanbao’. S1, S2, S3 and S4, S5, S6 represented the early (undifferentiation), middle (sepal differentiation and petal differentiation), and later (stamen differentiation and pistil differentiation) stage of the first and second flower bud differentiation, respectively. S1 was used as an internal control. (Mean ± SD, n = 3, p < 0.05)
Trehalose promoted early floweringIn order to confirm the effects of sucrose and trehalose on the twice flowering trait in M. liliiflora ‘Hongyuanbao’, the plants grown in the nursery of Zhejiang A&F University were sprayed with a solution of sucrose and trehalose once the plants finished the spring flowering on April 22, 2019. Leaf spraying commenced with three concentrations (30 mM, 60 mM, 90 mM) of sucrose and trehalose (Fig. 5A). The plants sprayed with deionized water were used as the control (CK). The leaves of all the plants were sprayed every 5 days and the samples were taken for microscopy observation on the 20th and 35th day following the first spraying.
Fig. 5
The effect of spraying on the leaves of M. liliiflora ‘Hongyuanbao’ plants with the solution of trehalose or sucrose. A Treatment with different concentrations of trehalose or sucrose solutions. The control and six treatment groups had three plants each. (n = 3). B The 20th and 35th day after treatment were chosen to observe the process of flower bud differentiation. Ten buds were harvested each time for dissection. Red bars, 50 μm; White bars, 2 mm
The results of paraffin section showed that on the 20th day, plants treated with trehalose showed accelerated development relative to the control and sucrose treatment plants. Under trehalose treatments, buds had entered the floral transition, while the buds under sucrose treatments and the control remained undifferentiated (Fig. 5B). However, the rate of development did not correlate directly with trehalose concentration. Plants treated with 90 mmol/L trehalose had only reached early flower bud differentiation while plants treated with 30 and 60 mmol/L had reach sepal and petal primordium differentiation respectively. On the 35th day, both trehalose and sucrose treatments could promote floral development process. For plants treated with trehalose, the 60 and 90 mM treatments had completed flower bud differentiation, and the 30 mM had reached the pistil differentiation stage. The plants sprayed with 60 mM of sucrose were in the pistil developmental stage, while plants sprayed with 30 mM and 90 mM of sucrose were in stamen developmental stage, the same as CK. Taken together, these results demonstrate that spraying of ‘Hongyuanbao’ leaves with different concentrations of sugars, either sucrose or trehalose, have variable effects on flower bud differentiation, with trehalose showing prominent promotion effects on the process of flower bud differentiation (Fig. 5B).
Expression of TPS genes and flowering-related genes under sucrose treatmentsTo more clearly understand whether trehalose and sucrose influence the twice flowering phenotype, we explored the expression of both sugar-responsive genes and genes involved in flowering regulation. Specifically, we focused on the TPS family of MlTPS1, MlTPS5, MlTPS6, MlTPS7 and MlTPS9, and key floral integrator genes MlFT, MlLFY, MlCO, and MlAP1. Previous studies have revealed that plants respectively sprayed with 60 mM of trehalose and sucrose could obviously promote flower bud differentiation process relative to the control and other treatment plants (Fig. 5B). Thus, the expression patterns of MlTPS genes and floral integrator genes in ‘Hongyuanbao’ leaves under treatments with 60 mM of trehalose and sucrose and CK condition were examined (Fig. 6).
Fig. 6
Expression levels of TPS and flowering genes in Magnolia liliiflora ‘Hongyuanbao’. 15d, 20d, 25d, 30d, 35d and 40d represented the days after the treatments with 60 mM of trehalose or sucrose. The bud’s stage at 20th day were undifferentiated under sucrose treatment and CK, and petal differentiation under trehalose treatment. At 35th day, the buds were stamen developmental stage under CK, pistil stage under sucrose treatment and accomplished the flower bud differentiation process trehalose treatment. (Mean ± SD, n = 3, p < 0.05)
It was apparent that the MlTPS genes were widely expressed throughout the flower bud differentiation period. In the treatment with a solution of 60 mM trehalose, relative to CK, MlTPS1 and MlTPS5 were increased during the middle (25d) and later stage (40d), and the expression of MlTPS6 was decreased following an initial increase. While the expression of MlTPS7 and MlTPS9 remained unchanged. The results indicated that MlTPS1 and MlTPS5 might be response to trehalose treatment to regulate flowering. Under the treatment with 60 mM sucrose, the expression level of MlTPS1 was barely detectable, and the expression of MlTPS5 decreased following an initial increase. The expressions of MlTPS6 and MlTPS7 was always higher than that of the CK, and the MlTPS9 expression remained unchanged, indicating that MlTPS6 and MlTPS7 might have an important function in sucrose treatment accelerating flowering. Moreover, the flowering integrators also responded to sugar treatment (Fig. 6). The expression of MlFT, which correlated with the timing of differentiation, was higher with trehalose and sucrose treatments than CK, suggesting that MlFT might play a vital role in flowering. The level of expression of MlLFY was higher in the beginning of flower differentiation compared to the later stages, suggestive of its functional role in the development of floral meristem. The expression of MlCO was raised moderately by the sugar treatments, while MlAP1 was not responsive. These results demonstrated that MlTPS genes might be response to sugar signal to regulate floral differentiation and the acceleration in flowering promotion might depend on the enhanced expression of MlFT and MlLFY.
Analysis of the expression of transcription factor SPL following sugar treatmentSPL genes are regulated by diverse flowering signals and involved in age pathway to regulate flowering [34]. In other species, SPL3 has been suggested as a participant in the T6P pathway and to affect the process of flowering [15]. Thus, we wanted to know whether SPL3 homologous genes in ‘Hongyuanbao’ could respond to trehalose and sucrose treatments. We identified two MlSPL3 genes (MlSPL3–1 and MlSPL3–2) from the ‘Hongyuanbao’ transcriptome and determined whether they could respond to trehalose and sucrose treatments on 20th day. MlSPL3–1 showed significantly higher expression relative to CK under both sugar treatments, with the trehalose treatments showing relatively higher expression than the sucrose treatment (Fig. 7A). While MlSPL3–2 had significant expression under trehalose treatments, and the overall expression of MlSPL3–2 was higher following spraying with trehalose than sucrose (Fig. 7B). These results suggest that the T6P pathway can affect the expression of flower-patterning gene MlSPL3 via the age pathway to promote the floral differentiation process.
Fig. 7
Analysis of the expression of SPL3 gene after treatment with sugars. A The relative expression of SPL3–1 on 20th day. (Mean ± SD, n = 3, p < 0.05). B The relative expression of SPL3–2 on 20th day. (Mean ± SD, n = 3, p < 0.05)
留言 (0)