
記住我






Casein is the insoluble major protein component of milk comprising about 80% of the total milk protein fraction (Bhat et al., 2016). Milk caseins have a fundamental nutritional value since they provide calcium and phosphorus for bone formation and amino acid synthesis in offspring (Barth and Schlimme, 2012; Ghosh et al., 2016). Milk composition is a multifactorial complex trait. Besides genetic factors, the total protein and casein content is affected by various factors such as number of milking per day, stage of lactation, age of the animal, parity, rearing practices, seasonal components, and nutrition (Assan, 2014).
In goats, like in other mammals, four evolutionary conserved casein genes occur. The genes encoding alpha-S1-, beta-, alpha-S2-, and kappa-caseins are CSN1S1, CSN2, CSN1S2, and CSN3, respectively (Martin et al., 2002). All four genes are located in a 250 kb region between 85.978 and 86.211 Mb on Capra hircus chromosome 6 (Ferretti et al., 1990; Threadgill and Womack, 1990).
Caseins occur in milk in micelles which are large spherical colloidal particles. These micelles are buildup of several submicelles which are bound together by colloidal calcium phosphate (Fox and McSweeney, 1998). Alpha-S1-, α-S2-, and β-caseins, which are hydrophobic phosphate and calcium-sensitive caseins, are located inside the inner part of the micelle (Hui and Sherkat, 2005), which precipitate from raw milk upon acidification at pH 4.6 at 20°C⁰ (Park et al., 2017). Kappa-casein, the insensitive calcium protein, resides on the surface of the micelles where it stabilizes the micelles with a protruding hydrophobic chain (Guinee and O'Brien, 2010).
The most important economic effects of the milk casein proteins come from the relationship between protein quality, amount, coagulation, and techno-functional properties to produce yoghurts and cheese. Casein polymorphisms also affect the fat content, micellar size, mineralization, and sensory characteristic in milk. Therefore, casein polymorphisms play a significant role both in milk coagulation performance and cheese quality. Since the identification of the impact of milk protein polymorphisms on milk quality and its technological properties (Aschaffenburg and Drewry, 1955), milk protein variants have been intensively characterized in different species including goats (Marletta et al., 2007; Caroli et al., 2009; Giambra and Erhardt, 2012; Selvaggi et al., 2014; Ozdemir et al., 2018). As a side effect of the intensive work, simultaneous publications led to inconsistency in the naming and positioning of some sequence variants which were anchored in part to mature and in part to immature proteins and sometimes include mutations in non-coding regions. In addition, many studies have referred to the genetic and protein variants of caseins in goats with different nomenclature. This review gives a comprehensive overview on identified protein and DNA variants of the four casein genes in goats. One goal is to clarify names and nomenclatures, the positions of nucleotide and amino acid polymorphisms and provide a translation from DNA to protein variant. We focus on protein variants, but also describe linked nucleotide variants in non-coding regions of the gene, like the promoter, 5′ and 3’ untranslated regions, and introns, which could affect the transcription and translation and thereby production traits. Furthermore, we have derived haplotypes across all four linked genes and summarize the frequency of alleles in different breeds and the impact of milk protein variants on milk quality for cheese making, human health and human nutrition.
CSN1S1At the functional level, alpha-S1-casein is required for the efficient export of other caseins from the endoplasmic reticulum to the Golgi compartment, the casein transport in the secretory pathway, and for the biogenesis of casein micelles in milk (Chanat et al., 1999; Le Parc et al., 2010). Alpha-S1-casein has an important role in the capacity of milk to provide calcium phosphate for human nutrition (Kolb et al., 2011).
The percentage of α-S1-casein in goat milk varies from 0 to 25%. Alpha-S1-casein concentrations has been shown to be dependent on the existence of polymorphisms in the coding and regulatory sequences of the gene (Boulanger et al., 1984). In contrast, the α-S1-casein fraction in bovine milk constitutes up to 40% (Farrell et al., 2004). Strong, intermediate, low, and null expression levels of α-S1-casein in goat milk are distinguished (Moatsou et al., 2004; Caroli et al., 2007; Kupper et al., 2010). In the first group comprising CSN1S1*A, CSN1S1*A3, CSN1S1*B1, CSN1S1*B2, CSN1S1*B3, CSN1S1*B4, CSN1S1*C, CSN1S1*H, CSN1S1*L, and CSN1S1*M are “strong” variants, which are associated with a high content of α-S1-casein in milk. CSN1S1*E and CSN1S1*I are “intermediate” variants associated with a medium content. CSN1S1*D, CSN1S1*F, and CSN1S1*G are “weak” variants associated with a low content, and CSN1S1*N, CSN1S1*01, and CSN1S1*02 are “null” variants leading to the absence of α-S1-casein in the milk of animals homozygote for these alleles (Grosclaude et al., 1987; Brignon et al., 1990; Chianese et al., 1997; Grosclaude and Martin, 1997; Martin et al., 1999; Bevilacqua et al., 2002; Ramunno et al., 2005; Mestawet et al., 2013). The α-S1-casein strong variants produced around 3.6 g/L per allele, medium variants produced 1.1–1.6 g/L per allele, weak variants produced 0.45–0.6 g/L per allele, and null variants resulted in absence of the α-S1-casein in milk (Grosclaude et al., 1987; Grosclaude and Martin, 1997; Martin et al., 1999). Strong variants with high content of α-S1-casein are most desired in dairy production since they are associated with lower pH values, better coagulation properties, faster curdling rate and greater gel firmness, which contribute largely to high yield and quality of cheese, even if the rennet coagulation time was longer (Ambrosoli et al., 1988; Pirisi et al., 1994; Clark and Sherbon, 2000; Schmidely et al., 2002; Zullo et al., 2005; Caravaca et al., 2011; Devold et al., 2011; Cebo et al., 2012; Vacca et al., 2014; Johansson et al., 2015; Marković et al., 2018). Since the casein content in milk also affects the size of casein micelles and the amount and composition of milk fatty acids, cheese made from milk with strong α-S1-variants has a better structure and taste with less typical goat flavor than cheese produced from intermediate or weak variants (Delacroix Buchet et al., 1996). Generally, the lower α-S1-casein content in goat milk compared to bovine milk is associated with smaller casein micelles, with less hydrated pores (Park et al., 2007; Ingham et al., 2018; Nguyen et al., 2018). Hence, yoghurt (Miocinovic et al., 2016; Nguyen et al., 2018) and cheese (Park et al., 2007; Mestawet et al., 2014) made from goat milk have a lower dense gel structure and poor coagulation features compared to their counterparts made with cow milk.
The CSN1S1 gene is located on chromosome 6 between 85.978 and 85.995 Mb and spans 16,808 bp. The DNA and protein reference sequences NC_030813 (goat genome reference version LWLT01 (Bickhart et al., 2017)) and XP_017904616 (NCBI database), respectively, represent the CSN1S1*A protein variant (Boulanger et al., 1984; Brignon et al., 1989; Ramunno et al., 2004). The gene encodes 214 amino acids of α-S1-casein, the first 15 amino acids form the signal peptide sequence, followed by 199 amino acids long mature protein (Brignon et al., 1990). It consists of 19 exons ranging in size from 24 (exons 5, 6, 7, 8, 10, 13, 16) to 385 bp (exon 19) and 18 introns from 90 bp (intron 10) to 1,685 bp (intron 2) (Ramunno et al., 2004).
In the following, we briefly describe the 20 protein variants that have been identified so far (Supplementary Table S1).
The α-S1-casein CSNS1*A is the reference protein. The sub variants CSN1S1*A′ and CSN1S1*A2 represent the same reference protein, namely CSN1S1*A, though CSN1S1*A′ carries a T/C transition in intron 9 (Kupper et al., 2010) and CSN1S1*A2 a G/A transition in the 5′ flanking region (Ramunno et al., 2005). The same G/A transition is present also in variants CSN1S1*N and CSN1S1*01, which are null variants (Ramunno et al., 2005), which means absence of α-S1-casein in the milk of homozygous carriers. Hence, the G/A variation in the 5′ flanking region is not responsible for the absence of α-S1-casein in milk. Different from the other CSN1S1*A proteins, CSN1S1*A3 carries a transversion from A to C at position CHR6:85987198 (exon 10) leading to the amino acid exchange from Gln77 to Pro77 (Mestawet et al., 2013). CSN1S1*A is the predominant protein variant in most Indian breeds (frequency >0.7) (Kumar et al., 2007; Rout et al., 2010) and in a few breeds from Spain, Italy and France (frequency >0.5, Supplementary Table S2) (Ramunno et al., 1991).
The CSN1S1*B protein is divided into four variants (CSN1S1*B1, CSN1S1*B2, CSN1S1*B3, CSN1S1*B4) (Grosclaude and Martin, 1997). All CSN1S1*B variants differ from the reference protein through the substitution of Gln77 to Glu77 (Brignon et al., 1989), a substitution that is associated with the presence of a phosphate group on Serine at position 75 (Brignon et al., 1989). Among all α-S1-casein variants, CSN1S1*B1 is considered the ancestor allele showing the closest homology to its bovine and ovine counterparts (Grosclaude and Martin, 1997). CSN1S1*B1 and CSN1S1*B′ are identical proteins; the only difference is a synonymous SNP in the codon for Ser66 in exon 9 in CSN1S1*B′ (Caroli et al., 2007). The caseins CSN1S1*B2, CSN1S1*B3 and CSN1S1*B4 carry the additional T to C transition at position CHR6:85982615 (exon 4), leading to the substitution of Leu16 to Pro16 (Grosclaude and Martin, 1997; Cosenza et al., 2008). CSN1S1*B3 and CSN1S1*B4 carry the G to A transition at position CHR6:85988705 (exon 12) changing Arg100 to Lys100 (Grosclaude and Martin, 1997; Cosenza et al., 2008). CSN1S1*B4 differs from CSN1S1*B3 by the nucleotide substitution A to G at position CHR6:85993465 (exon 17) altering Thr195 to Ala195 (Grosclaude and Martin, 1997; Cosenza et al., 2008). With respect to names, it is necessary to mention that firstly, CSN1S1*B2 was named simply CSN1S1*B in old papers (Boulanger et al., 1984; Brignon et al., 1989), and secondly, the variant CSN1S1*E had also been named CSN1S1*B− (Grosclaude et al., 1987). CSN1S1*B protein variants can be found at high frequency (>0.5) in diverse breeds across the world (Supplementary Table S2).
CSN1S1*C and CSN1S1*C1 differ from the reference by five and four amino acid substitutions, respectively. The two variants carry the amino acid substitutions His8 to Ile8, Leu16 to Pro16, Gln77 to Glu77, and Arg100 to Lys100. The additional substitution Thr195 to Ala195 occurs only in CSN1S1*C (Leroux et al., 1990; Rahmatalla et al., 2021).
CSN1S1*D is characterized by a deletion of 11 amino acids at positions 59 to 69 which comprises the major phosphorylation site of the protein (Brignon et al., 1990). The deletion of the single nucleotide C at position CHR6:85986426 in exon 9 is a splice-site mutation which causes the exclusion of exon nine in the mature RNA assembly which also reduces the synthesis rate of the protein (Mahé and Grosclaude, 1989; Brignon et al., 1990). Besides the significant deletion, CSN1S1*D carries the substitutions Leu16 to Pro16 and Gln77 to Glu77, which also occur in most CSN1S1*B and CSN1S1*C protein variants (Mahé and Grosclaude, 1989; Brignon et al., 1990).
CSN1S1*E (formerly CSN1S1*B′) has the same amino acid variant as CSN1S1*B4. Besides that, CSN1S1*E has an insertion of 457–458 bp in exon 19, which is expressed but untranslated (Jansàpérez et al., 1994). This insert is a truncated long interspersed repeated element (LINE), containing part of the ORF-2, the 3′ UTR, and the original retrotransposon’s poly (A) tail. The mutant 3′ UTR likely controls the mRNA stability of transcripts and/or the translation rate which was seen as low casein content in goat milk of CSN1S1*E carriers (Jansàpérez et al., 1994). The CSN1S1*E allele is highly frequent (>0.5) in Saanen goats worldwide (Tadlaoui Ouafi et al., 2002; Maga et al., 2009), and in other breeds from Spain, France, and the United States (Supplementary Table S2) (Grosclaude et al., 1994; Jordana et al., 1996; Tadlaoui Ouafi et al., 2002; Caravaca et al., 2008; Maga et al., 2009).
CSN1S1*F contains the amino acid variants Pro16 and Glu77 as in CSN1S1*D (Grosclaude et al., 1987; Brignon et al., 1990). Most significant in CSN1S1*F is the lacks of 37 amino acids from position 59 to 95 as the consequence of a frameshift mutation due to a single nucleotide deletion in exon 9 (Grosclaude et al., 1987; Brignon et al., 1990). The deletion starts exactly at the same position and with the same mutation as in CSN1S1*D. Two additional insertions of 11 and 3 base pairs in intron nine are likely responsible for the skipping of exons 9, 10 and 11 in CSN1S1*F (Leroux et al., 1992). The content of this shortened α-S1-casein protein variant in milk is low, likely as a result of low efficiency and accuracy of the splicing machinery (Martin et al., 1999), which might result from either a reduced translation rate or intracellular degradation of the incomplete protein. Several breeds that originate from the Alpine region carry the CSN1S1*F allele at a frequency above 0.40 (Grosclaude et al., 1987; Ramunno et al., 1991). A higher expression of CSN1S1*F compared to CSN1S1*N might be attributed to a mutation in the promotor region creating an extra putative activator protein 1 (AP-1) binding motif (Ramunno et al., 2005). However, experimental validation does not exist. Particular high frequency >0.90 of CSN1S1*F was found in Toggenburg goats from America and Orobica goats from Italy (Supplementary Table S2) (Caroli et al., 2006; Maga et al., 2009).
Protein variant CSN1S1*G is missing 13 amino acids from position 14 to 26. The G/A transition at position CHR6:85982647 in the 5′ splice site at the beginning of intron 4 leads to the skipping of exon 4 (Martin and Leroux, 1994). The occurrence of the CSN1S1*G allele is associated with low α-S1-casein content in milk (Martin and Leroux, 1994).
CSN1S1*H differs from the reference variant CSN1S1*A by a single amino acid substitution at position 1, Arg1 to Lys1. This protein was classified as a strong variant (Chianese et al., 1997). For the Italian Garganica breed the frequency of 0.07 was reported (Sacchi et al., 2005).
For the protein variants CSN1S1*G, CSN1S1*I, CSN1S1*J, CSN1S1*K, CSN1S1*L, and CSN1S1*M there is information available for sequence variants only, but not for allele frequency in any breed. CSN1S1*I had the same amino acids sequence as variant CSN1S1*A. However, CSN1S1*I had only about half of the amount of casein in the milk than CSN1S1*A (Chianese et al., 1997), which led to the differentiation between CSN1S1*I and CSN1S1*A. The causal mutation is unidentified.
CSN1S1*J carries the combination of the amino acids substitutions Gln77 to Glu77 and Arg100 to Lys100, which are also found in other protein variants (Rahmatalla et al., 2021). CSN1S1*J detected in Bezoar ibex. This variant is similar to CSN1S1*B1 which is consider the ancestor allele, but it contains the Arg100 to Lys100 substitution (Rahmatalla et al., 2021).
The CSN1S1*K protein variant differs from the reference by the unique amino acid substitution from Ile44 to Val44 as a results of a to G transition at position CHR6:85984154 (exon 7) (Rahmatalla et al., 2021). The CSN1S1*K was found in wild Alpine ibex (Rahmatalla et al., 2021).
CSN1S1*L harbors the two amino acids Pro16 and Glu77 instead of Leu16 and Gln77 in the reference sequence and additionally the amino acid substitution from Arg90 to His90 (Chianese et al., 1997). The transition of G to A at CHR6:85987327 in exon 11 leads to the substitution of Arg90 to His90.
CSN1S1*M carries the Leu16 to Pro16 substitution, which occurs also in other protein variants. CSN1S1*M also contains the amino acid substitution of Ser66 to Leu66 which results from a C/T transition at CHR6:85986426 in exon 9 (Chianese et al., 1998; Bevilacqua et al., 2002).
CSN1S1*N, CSN1S1*01 and CSN1S1*02 are null variants resulting in truncated gene products and subsequently no casein in the milk. CSN1S1*N descends from protein variant CSN1S1*M, but contains also the nucleotide deletion of a single C at CHR6:85986426 in exon 9 that also occurs in variants CSN1S1*D and CSN1S1*F, where it causes exon skipping. However, in CSN1S1*N the one-nucleotide deletion leads to a premature stop codon. The amount of the truncated transcript is reduced to one-third and the amount of α-S1-casein in milk in homozygous carriers is null (Ramunno et al., 2002; Ramunno et al., 2005). CSN1S1*01 is a truncated protein where the last seven exons are missing. The truncation results from a deletion of about 8.5 kb starting from nucleotide 181 of intron 12. In homozygous carriers, α-S1-casein is absent in milk (Grosclaude et al., 1987; Leroux et al., 1990; Cosenza et al., 2003). CSN1S1*02 is another null variant, leading to the absence of the respective protein in milk. Martin et al. (1999) proposed a large insertion that has not been identified so far. Albeit the goats carrying the null alleles homozygous do not have contain α-S1-casein in their milk, the Indian breeds Ganjam and Local MP have an allele frequencies of 0.40 and 0.45, respectively (Rout et al., 2010).
Goat milk that lacks α-S1-casein has a higher pH value and a low content of chemical components like total solids, protein, casein, Nitrogen, fat, and Phosphorus compared to milk with strong α-S1-casein variants (Ambrosoli et al., 1988; Pirisi et al., 1994; Clark and Sherbon, 2000; Devold et al., 2011; Johansson et al., 2015; Marković et al., 2018). In addition, milk containing α-S1-casein null variants is characterized by large casein micelles with weak gel firmness and long coagulation time (Devold et al., 2011; Johansson et al., 2015). The gross yield of cheese and the content of fat and non-fat solids as well as the fat to dry matter ratio and the total solids recovery rate from milk that lacks α-S1-casein is low (Pirisi et al., 1994). Furthermore, textural parameters of cheese such as hardness and plasticity were low in cheese that is produced from goat milk lacking α-S1-casein (Pirisi et al., 1994). In contrast, milk containing strong α-S1-casein variants produce firmer curd that is associated with high gross yield of cheese with a better composition (Devold et al., 2011; Johansson et al., 2015).
CSN2Beta-casein is the most hydrophobic casein as a result of a highly charged N-terminal domain and an anionic phosphoserine cluster (Swaisgood, 2003). Beta-casein is the major protein in goat milk (Clark and Mora Garcia, 2017) comprising up to 50% of the total casein content. In comparison, human milk casein contains 60%–70% and bovine milk casein 36% β-casein (Guo, 2009; Elagamy, 2016). Beta-casein is a primary source of essential amino acids and facilitates the mineral transport for infants (Sadler and Smith, 2013). The peptides derived from β-casein during digestion have antihypertensive and immune-stimulating properties (Silva et al., 2006; Lamothe et al., 2007).
The CSN2 gene is located on chromosome 6 between 86.006 and 86.015 Mb (NC_030813, goat genome reference version LWLT01) (Bickhart et al., 2017). The 9,072 bp long gene consists of nine exons ranging in size from 24 (exon 5) to 492 bp (exon 7) (Roberts et al., 1992; Hayes and Petit, 1993). The protein contains 222 amino acids, which comprise 207 functional amino acids and 15 amino acids of a signal peptide (XP_005681778 (NCBI database).
So far, eight protein variants have been identified (Table 1). Variants CSN2*A, CSN2*B, CSN2*C, CSN2*D, and CSN2*E are “strong” variants associated with high β-casein content in milk (5 g/L per allele) (Roberts et al., 1992; Mahé and Grosclaude, 1993; Neveu et al., 2002; Galliano et al., 2004; Cosenza et al., 2005; Caroli et al., 2006). The variants CSN2*C2 and CSN2*F1 were associated with intermediate β-casein content (3.3 and 2.7 g/L per allele, respectively) (Nicolai et al., 2021). The two null alleles CSN2*0 and CSN2*0′ are associated with a lack of β-casein in milk (Persuy et al., 1999). In the CSN2 gene, the existence of the strong variants were associated with the highest milk protein content and yield in Sarda goats from Italy (Vacca et al., 2014).
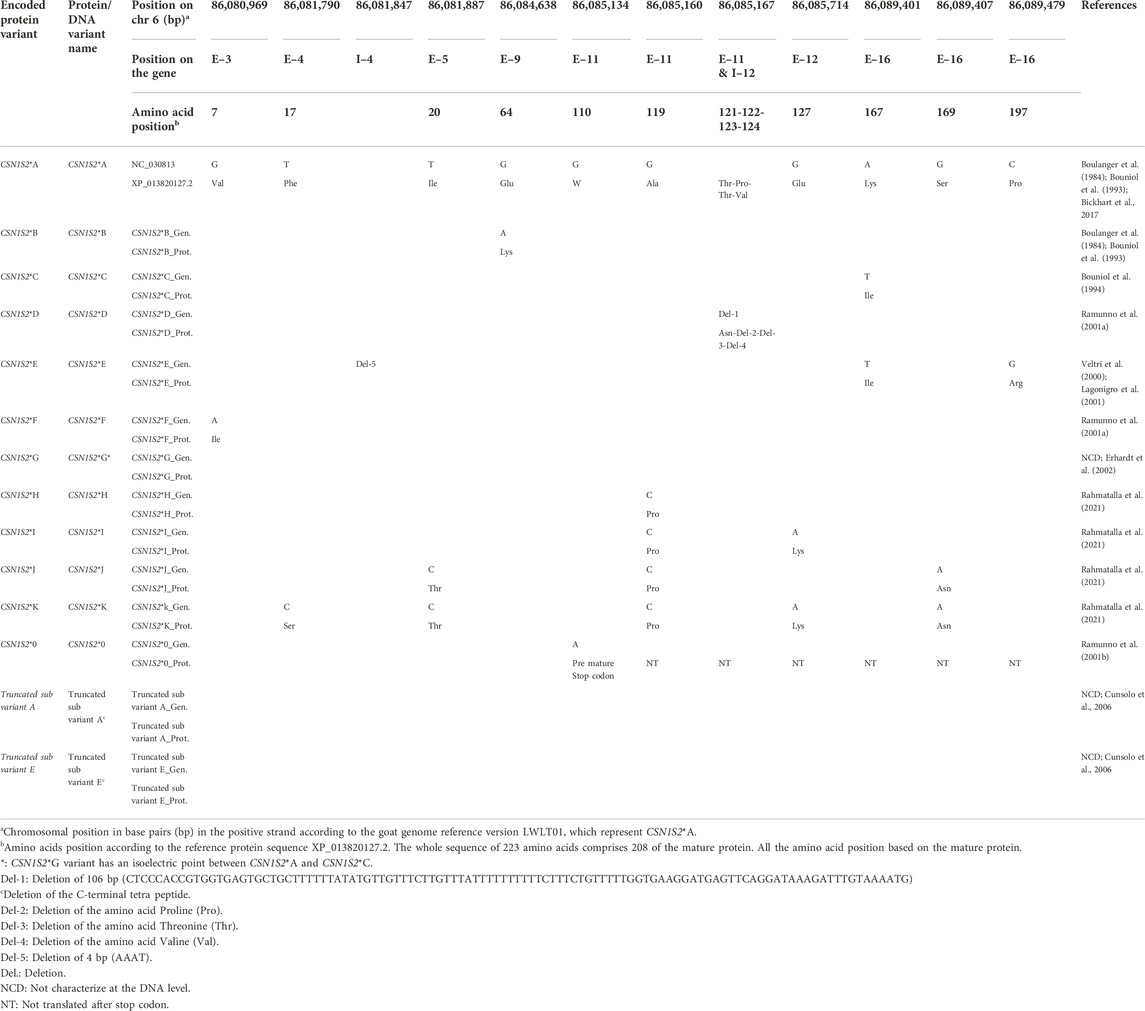
TABLE 1. Nucleotide and amino acid variants within the gene and mature protein of the CSN2.
The reference protein is variant CSN2*C (Wang et al., 2001; Neveu et al., 2002). CSN2*A differs from the reference by the amino acid substitution of Val177 to Ala177 (Neveu et al., 2002) as a result of an A/G transition at position CHR6:86008016 in exon 7 (Roberts et al., 1992). CSN2*A is considered the ancestral allele in goats because the amino acid alanine at position 177 is also found in sheep, cattle and buffalo (Chessa et al., 2005). The variant CSN2*A1 differs from CSN2*A by an additional G/A SNP in exon 9, which belongs to the 3′ untranslated region (Cosenza et al., 2005). CSN2*A was predominant with a frequency above 0.90 in Girgentana and Argentata dell’Etna from Italy, and in most Indian goats (Supplementary Table S3) (Marletta et al., 2005; Rout et al., 2010). A high frequency above 0.70 was evident in French Creole goats, West African breeds, and Buren goat from Germany (Supplementary Table S3) (Mahé and Grosclaude, 1993; Caroli et al., 2007; Kupper et al., 2010).
The CSN2*B variant was characterized via isoelectric protein electrophoresis, where CSN2*B migrates faster than CSN2*A (Mahé and Grosclaude, 1993). The mobility differences between CSN2*A and CSN2*B might be due to an extra phosphate group in CSN2*B (Mahé and Grosclaude, 1993).
CSN2*C1 and CSN2*C2 encode the same protein CSN2*C, the reference protein sequence. The protein variant CSN2*C is predominant with a frequency of ≥0.70 in most Italian goats, Angora and Hair goats from Turkey, Rajasthan and South goats from India as well as Banat White goats from Romania (Supplementary Table S3) (Chessa et al., 2005; Caroli et al., 2006; Chessa et al., 2007; Kusza et al., 2016). The two subtypes differ in a synonymous transition of G to A in the 3′ untranslated region of exon 9 in CSN2*C (Chessa et al., 2008; Nicolai et al., 2021). This single nucleotide polymorphism likely leads to lower amount of ß-casein if the animals is homozygouse for the CSN2*C2 subtype compared to CSN2*C1. An impact on the mRNA stability and or translation rate might be causal for the low amount of the protein (Nicolai et al., 2021). The variant CSN2*C1 was found with a frequency of more than 0.10 in Bunte Deutsche Edelziege and Weiße Deutsche Edelziege goats from Germany and Italian Girgentana goats (Kupper et al., 2010; Tortorici et al., 2014).
CSN2*D differs from the reference protein in the amino acid substitution from Val207 to Asn207 (Galliano et al., 2004). DNA sequence comparisons show a AC/TT substitution at positions CHR6:86007341 and CHR6:86007342 in exon 8 that is responsible for the amino acid change.
CSN2*E is characterized by the amino acid exchange from Ser166 to Thr166 which results from the G/T transversion at position CHR6:86008049 in exon 7 (Caroli et al., 2006). CSN2*E was found at a low frequency (0.08) only in Italian Frisa goats (Caroli et al., 2006).
The protein variant CSN2*F differs from the reference by the substitution of Asp47 to Thr47 (Chianese et al., 2007; Moatsou et al., 2007). The underlying SNP is a C/A transversion at position CHR6:86008407 in exon 7. By mistake, this protein variant had been named CSN2*E by Caroli et al. (2006) although CSN2*E already existed. It was correctly renamed as CSN2*F by Martin et al. (2013). CSN2*F1 and CSN2*F represent the same β-casein protein. CSN2*F1 differs from CSN2*F by a synonymous transition G/A in the 3′ untranslated region in exon 9 (Nicolai et al., 2021).
CSN2*G (previously CSN2*F) is characterized by the amino acid substitution from Pro148 to Leu148 and found in Wild Alpine ibex (Rahmatalla et al., 2021). The nucleotide mutation is a G/A substitution at position CHR6:86008103 in exon 7 (Rahmatalla et al., 2021).
Observations of milk lacking β-casein led to the conclusion that at least two null alleles exist (Dall’Olio et al., 1989; Mahé and Grosclaude, 1989). Variant CSN2*0 has one nucleotide deletion (T/-) at position CHR6:86008401 in exon 7 which introduces a premature stop codon at position 58 in the mature protein (Persuy et al., 1999). The CSN2*0 null variant was detected in a frequency below 0.10 in Creole goats from France and some Italian and Indian goats (Supplementary Table S3) (Mahé and Grosclaude, 1993; Marletta et al., 2005; Sacchi et al., 2005; Rout et al., 2010; Vacca et al., 2014). However, the frequency was 0.19 in Italian Garganica goats (Albenzio et al., 2009). CSN2*0′ carries the single nucleotide substitution C to T at position CHR6:86008047 in exon 7, which introduces a premature stop codon at position 167 of the mature protein (Rando et al., 1996). The CSN2*0′ null variant was found at a frequency below 0.10 in some Italian breeds and in Arbi goats from Tunis (Supplementary Table S3) (Chessa et al., 2005; Gigli et al., 2008; Vacca et al., 2009a; Tortorici et al., 2014). In CSN2*0′, another SNP in the promoter region was detected which negatively affects the transcriptional activity of the gene (Cosenza et al., 2007; Cosenza et al., 2016).
No information was available for the allele frequency of CSN2*C2, CSN2*D, CSN2*F, CSN2*F1, and CSN2*G.
The β-caseins in milk are primarily classified into the two types A1 and A2. A1 and A2 differ in the generation of β-casomorphin-7 (BCM-7) peptides during digestion (Jianqin et al., 2016). The peptide β-casomorphin-7 gained attention because it was associated with gastrointestinal disturbances, ischemic heart disease, arteriosclerosis, type 1 diabetes, and sudden infant death syndrome in humans (Birgisdottir et al., 2002; Tailford et al., 2003; Ho et al., 2014). The peptide β-casomorphin-7 is uniquely derived from the digestion of β-casein type A1, but not from A2. Different from cow milk, goat milk mainly contains the β-casein type A2 (Cieslinska et al., 2012). Therefore, β-casein of the type A2 contributes to the healthy properties of goat milk because it does not produce the peptide β-casomorphin-7 during digestion (De Noni and Cattaneo, 2010).
It is worth noting that with the increasing awareness of consumers’ behavior and their interest in healthy eating and functional food, different companies launched β-casein type A2 milk from cattle on the market. Costs are associated with the separate handling of this type of milk during processing. Since goat milk naturally contains β-casein type A2, this milk plays a crucial role in providing functional milk and milk products to the food market. A positive benefit for human health is realized here without additional marketing costs.
CSN1S2The alpha-S2-casein protein plays an important role in the transport of calcium phosphate. The alpha-S2-casein content in goat milk is relatively high comprising about 19%, while it constitutes up to 10% in bovine milk (McMahon and Brown, 1984). The antimicrobial peptide cathelicidin-1 which derives from the digestion of alpha-S2-casein has the activity to inhibit the growth of Gram-positive and Gram-negative bacteria (Zucht et al., 1995; López-Expósito et al., 2008; Gigli, 2016).
The CSN1S2 gene resides on chromosome 6 between 86.077 and 86.094 Mb, which is about 16,695 bp long and consists of 18 exons (NCBI, accession no. NC_030813). The gene encodes 223 amino acids comprising 208 functional amino acids and 15 amino acids of a signal peptide (https://www.uniprot.org/uniprot/P33049).
The DNA [NC_030813, goat genome reference version LWLT01 (Bickhart et al., 2017)] and protein reference sequence (XP_013820127.2, NCBI database) represent the CSN1S2*A variant (Boulanger et al., 1984; Bouniol et al., 1993; Bickhart et al., 2017). CSN1S2*A is considered the ancestor variant for goat according to the evolutionary path from sheep and cattle (Sacchi et al., 2005). This variant is common with a frequency of ≥0.70 in distinct Saanen goats, most Italian, German and Indian goats, Dwarf goats from Nigeria, and Nubian goats from Sudan (Supplementary Table S4) (Boulanger et al., 1984; Bouniol et al., 1994; Ramunno et al., 2001b; Erhardt et al., 2002; Marletta et al., 2004; Chiatti et al., 2005; Marletta et al., 2005; Caroli et al., 2007; Chessa et al., 2007; Kupper et al., 2010; Rout et al., 2010; Yue et al., 2013).
So far, 14 alpha-S2-casein protein variants have been identified (CSN1S2*A, CSN1S2*B, CSN1S2*C, CSN1S2*D, CSN1S2*E, CSN1S2*F, CSN1S2*G, CSN1S2*H, CSN1S2*I, CSN1S2*J, CSN1S2*K, CSN1S2*0, truncated sub-variant A, and truncated sub-variant E) (Table 2) (Boulanger et al., 1984; Bouniol, 1993; Bouniol et al., 1994; Veltri et al., 2000; Ramunno et al., 2001a; Ramunno et al., 2001b; Lagonigro et al., 2001; Erhardt et al., 2002; Cunsolo et al., 2006; Rahmatalla et al., 2021). While the protein variants CSN1S2*A, CSN1S2*B, CSN1S2*C, CSN1S2*E, CSN1S2*F, and CSN1S2*G are associated with normal amounts of α-S2-casein of about 2.5 g/L per allele (Boulanger et al., 1984; Grosclaude et al., 1987; Bouniol et al., 1994; Ramunno et al., 2001a), CSN1S2*D is associated with a reduced amount of about 1.5 g/L per allele (Ramunno et al., 2001a). In CSN1S2*0 homozygous goats, alpha-S2-casein was not detectable (Ramunno et al., 2001b).
TABLE 2. Nucleotide and amino acid variants within the gene and mature protein of the CSN1S2.
CSN1S2*B differs from the reference (CSN1S2*A) by a G/A substitution at CHR6:86084638 in exon 9, which is responsible for the amino acid substitution Glu64 to Lys64 that affects a phosphorylation site (Boulanger et al., 1984; Bouniol et al., 1994). CSN1S2*B occurs at a frequency above 0.40 in Egyptian and Lithuanian Native goats (Othman and Ahmed, 2006; Baltrėnaitė et al., 2009).
CSN1S2*C distinguishes from the reference by the transversion A/T at CHR6:86089401 in exon 16 which changing Lys167 to Ile167 (Bouniol et al., 1994). This variant occurs at a frequency ≥0.40 in Vallesana and Roccaverano Italian goats, Dwarf Cameroon goats, Borno goats from Nigeria, Hair goats from Turkey, as well as Maharashtra and Rajasthan from India (Supplementary Table S4) (Sacchi et al., 2005; Caroli et al., 2007; Chessa et al., 2007).
CSN1S2*D contains a 106 bp deletion comprising the last 11 bp of exon 11 and the first 95 bp of the following intron (Ramunno et al., 2001a). This deletion is responsible for the lack of the three codons Pro122, Thr123, and Val124. Furthermore, the last undeleted nucleotide (A) of exon 11 together with the two nucleotides (AT) of the following intron generate a codon for the amino acid Asn121 instead of Thr121.which is followed by a new GT dinucleotide splicing donor site. The rare CSN1S2*D variant was found in Italian Argentata dell’Etna and Girgentana goats and Hungarian goats with a frequency of 0.01 (Kusza et al., 2007; Gigli et al., 2008).
CSN1S2*E is characterized by a C/G mutation at CHR6: 86089479 in exon 16, leading to the amino acid exchange Pro197 to Arg197 (Chianese et al., 1998; Veltri et al., 2000; Lagonigro et al., 2001). Additionally, the of A/T transversion of CSN1S2*C changing Lys167 to Ile167 is present also in CSN1S2*E. Furthermore, a deletion of 4 bp (AAAT) was identified in intron 4 (Lagonigro et al., 2001). CSN1S2*E was found at low frequency (below 0.11) in some Italian goats, Hair goats from Turkey, and South goats from India (Sacchi et al., 2005; Caroli et al., 2006; Chessa et al., 2007; Gigli et al., 2008; Albenzio et al., 2009; Vacca et al., 2009b; Palmeri et al., 2014; Vacca et al., 2014). In addition to the full-length protein variants CSN1S2*A and CSN1S2*E, two truncated sub-variants were identified, which lack the C-terminal tetrapeptide (Gln199, Pro200, Leu201, Gln202, Pro203, Leu204). These truncated variants likely originate from differential splicing of pre-messenger RNA during the translation process of CSN1S2*A and CSN1S2*E variants (Cunsolo et al., 2006).
CSN1S2*F differs from the reference by a G/A transition at CHR6:86080969 in exon 3 substituting Val7 to Ile7 (Ramunno et al., 2001a). CSN1S2*F is common (frequency ≥0.40) in Italian Garganica, Orobica, Frisa, Sarda goats, and Weiße Deutsche Edelziege (Sacchi et al., 2005; Caroli et al., 2006; Vacca et al., 2009b; Kupper et al., 2010).
There is not much known about the nucleotide and protein sequence of the variant CSN1S2*G. This protein has an isoelectric point between CSN1S2*A and CSN1S2*C (Erhardt et al., 2002).
CSN1S2*H differs from CSN1S2*A by a G/C substitution at CHR6:86085160 in exon 11 leading to the substitution of Ala119 by Pro119 (Rahmatalla et al., 2021). This variant is predominant in Sudanese goats (Supplementary Table S4) (Rahmatalla et al., 2021).
CSN1S2*I contains the same mutations that distinguish CSN1S2*G from the reference and additionally a G/A substitution at CHR6:86085714 in exon 12 that is responsible for the change of Glu127 to Lys127 (Rahmatalla et al., 2021). So far, CSN1S2*I has been detected in Nubian and Desert goats only at a frequency of 0.21 and 0.10, respectively (Rahmatalla et al., 2021).
CSN1S2*J diverges from the reference by a T/C substitution at CHR6:86081887 in exon 5 and G/A substitution at CHR6:86089407 in exon 16 that account for the substitutions Ile20 to Thr20 and Ser169 to Asn169, respectively (Rahmatalla et al., 2021). In addition, the mutation which distinguishes CSN1S2*H from CSN1S2*A is present also in CSN1S2*J (Rahmatalla et al., 2021).
CSN1S2*K carries the same three amino acid substitution that occur in CSN1S2*J (Rahmatalla et al., 2021) and additionally the amino acid substitutions Phe17 to Ser17 and Glu127 to Lys127 as a result of the nucleotide substitutions of T to C at CHR6:86081790 in exon 4 and G to A at CHR6:86085714 in exon 12, respectively (Rahmatalla et al., 2021) The CSN1S2*J and CSN1S2*K were identified in wild Nubian ibex (Rahmatalla et al., 2021).
CSN1S2*0 carries a stop codon at the amino acid position 110 as a result of a G to A transition at CHR6:86085134 in exon 11, which disrupts the codon for Try110 (Ramunno et al., 2001b). As a consequence, a truncated non-functional protein of 109 instead of 208 amino acids is generated. Homozygous CSN1S2*0 carriers do not have α-S2-casein in milk. Nevertheless, the allele frequency in the Indian breeds Ganjam and Local MP is 0.55 and 0.48, respectively (Rout et al., 2010).
No information was available for the allele frequency of CSN1S2*J, CSN1S2*K, and the truncated sub-variants of CSN1S2*A and CSN1S2*E.
CSN1S2 variants affected the milk protein content and milk yield in Sarda goats from Italy (Vacca et al., 2014). In Sarda goats, CSN1S2*AC genotyped animals produce the highest fat and protein yield, which affect the technological properties for cheese processing, while animals carrying the CSN1S2*CF genotype have the highest daily milk yield (Vacca et al., 2014). Interestingly, the variants CSN1S2*A, CSN1S2*C, and CSN1S2*F were associated with a high amount of α-S2-casein in milk and produced 2.5 g/l per allele (Boulanger et al., 1984; Bouniol et al., 1994; Ramunno et al., 2001a). In Chinese dairy goats, animals carrying the CSN1S2*FF genotype have higher milk fat and total solids content than CSN1S2*AA and CSN1S2*AF genotypes, but do not have lower milk yield (Yue et al., 2013). However, Lan et al. (2005) found significant lower milk yield of Xinong Saanen goats carrying the CSN1S2*FF genotypes compared to other genotypes. The contradictory effects of CSN1S2*FF genotyped animals might be attributed to the small sample size in the study of Lan et al. (2005). The variation in the α-S2-casein level in milk is associated with the unique physicochemical characteristic of milk. Thereby, α-S2-casein levels affect the cheese-making properties (Selvaggi and Tufarelli, 2011).
CSN3Kappa-casein (k-casein) is a calcium insensitive protein that forms a protective layer around the calcium-sensitive caseins α-S1, α-S2, and β, resulting in stable casein micelles (Quaglia and Gennaro, 2003). The formation, stabilization, and aggregation of casein micelles affect the physical characteristics and technological properties of milk (Alexander et al., 1988; Glantz et al., 2010). Kappa-casein is the only casein that is highly glycosylated (Haratifar and Guri, 2017). The mature kappa-casein protein has a labile peptide bond whose cleavage by chymosin or rennin produces a soluble hydrophilic glycopeptide called caseino-macropeptide (amino acids 106–171) and an insoluble peptide or para-k-casein (amino acids 1–105) (Yahyaoui et al., 2003). The caseino-macropeptide is important for milk coagulation.
Kappa-casein comprises about 20% in goat and sheep milk, in comparison, only 12–14% in cow milk (Fox and Brodkorb, 2008; Molik et al., 2012). Kappa-casein is essential for lactation. For example, mice lacking this gene (CSN3−/−) fail to lactate because of destabilization of the micelles in the lumen of the mammary gland (Shekar et al., 2006). Kappa-casein contributes to the nutritional properties of milk. The protein has beneficial effects on oral health (Vacca-Smith and Bowen, 1995; Johansson and Lif Holgerson, 2011) and the digestion of k-casein generates an antimicrobial peptide (Malkoski et al., 2001).
The CSN3 gene is located on chromosome 6 between 86.197 and 86.211 Mb [NC_030813, goat genome reference version LWLT01 (Bickhart et al., 2017)]. The gene spans 14,114 bp and comprises five exons (Martin et al., 2002). The protein contains 192 amino acids which include 21 amino acids of the signal peptide and 171 amino acids of the mature protein (Mercier et al., 1976; Coll et al., 1993). The mature protein is encoded in only two exons, exons 3 (9 amino acids) and exon 4 (162 amino acids) (Yahyaoui et al., 2003).
The CSN3 gene in goats is highly polymorphic. To date, 35 variants are known that yield 24 protein variants and 11 synonymous mutations which are detectable only at the DNA level (Supplementary Table S5). In this review we follow the new nomenclatures of the k-casein variants recently described by Gautam et al. (2019). The DNA (NC_030813, goat genome reference version LWLT01 (Bickhart et al., 2017)) and protein reference sequences of CSN3 (NP_001272516, NCBI database) represent the protein variant CSN3*B as the reference (Yahyaoui et al., 2001; Jann et al., 2004). We supplement the information with nucleotide positions in the coding sequence that is given in the reference X60763 (NCBI database) (Supplementary Table S5) (Coll et al., 1993) to facilitate the comparison between the position in the new goat reference genome of k-casein variants with previous studies that used the position in the coding sequence to name variants.
The reference protein CSN3*B is the predominant k-casein variant that occurs at a frequency >0.70 in most Italian and East Africa goats, Toggenberg and Buren goats from Germany, and Polish Saanen, fawn and white improved goats (Supplementary Table S6) (Chessa et al., 2003; Yahyaoui et al., 2003; Sacchi et al., 2005; Caroli et al., 2006; Kiplagat et al., 2010; Kupper et al., 2010; Strzelec and Niżnikowski, 2011; Rahmatalla et al., 2021).
The difference between CSN3*A and CSN3*B is the amino acid substitution Ile119 to Val119 which is the result of the transition of A to G at CHR6:86209184 in exon 4 (Coll et al., 1993; Yahyaoui et al., 2001). CSN3*A occurs also at a frequency ≥0.7 in Ionica and Montefalcone from Italy, Bunte and Weise Deutsche Edelziege, and Thueringer Waldziege from Germany as well as Ardi and Syrian goats from Saudi Arabia (Supplementary Table S6) (Caroli et al., 2001; Angiolillo et al., 2002; Kumar et al., 2009; El-Shazly et al., 2017). CSN3*A′ and CN3*A″ encode the same protein variant CSN3*A, but, carry the synonymous mutations T to C at CHR6:86209207 and C to T at CHR6:86209258 in exon 4, respectively (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*A″ was found at low frequency in goats from Kenya and Maasai from Ethiopia. In that study, CSN3*A″ was named CSN3*O (Kiplagat et al., 2010).
CSN3*B′, CSN3*B″, CSN3*B‴ have the equivalent CSN3*B protein variant. In addition to the transition of G to A at CHR6:86209184 that alters the amino acid Val119 to Ile119, CSN3*B′, CSN3*B″, CSN3*B″ carry the synonymous mutations C to T at CHR6:86208883, C to T at CHR6:86209003, and A to G at CHR6:86209297 in exon 4, respectively (Supplementary Table S5) (Chessa and Bolla, 2003; Yahyaoui et al., 2003; Jann et al., 2004; Prinzenberg et al., 2005; Kiplagat et al., 2010). Previously, CSN3*B, CSN3*B′, CSN3*B″, and CSN3*B‴ had the names CSN3*D, CSN3*D′, CSN3*D″, and CSN3*P, respectively (Jann et al., 2004; Kiplagat et al., 2010). The homozygous CSN3*B genotype was associated with a higher milk protein content in Saanen goats (Catota-Gómez et al., 2017) and longer milk coagulation time in Spanish Murciano-Granadina goats compared to CSN3*A carriers (Caravaca et al., 2011).
CSN3*C differs from the reference variant CSN3*B in the three amino acid substitutions Val65 to Ile65, Ala156 to Val156, and Ser159 to Pro159 (Yahyaoui et al., 2001). The amino acid substitutions occurred as a result of a G to A, C to T, and T to C transitions at position CHR6:86209022, CHR6:86209296, and CHR6:86209304 in exon 4 (Yahyaoui et al., 2001), respectively. CSN3*C occurs at a frequency below 0.20 in some breeds from Italy, Canaria Spanish goats, goats from Germany and Turkey, Saanen goats worldwide, and Lithuania goats (Supplementary Table S6) (Chessa et al., 2003; Yahyaoui et al., 2003; Prinzenberg et al., 2005; Sacchi et al., 2005; Caroli et al., 2006; Gigli et al., 2008; Baltrėnaitė et al., 2009; Vacca et al., 2014; Catota-Gómez et al., 2017). CSN3*C′ differs from CSN3*C through a synonymous mutation in codon 131 (Prinzenberg et al., 2005).
In the CSN3*D (previously named CSN3*B), the amino acid substitutions Val65 to Ile65 and Ser159 to Pro159 that exist in CSN3*C were also found. In addition, CSN3*D carries the Gln44 to Arg44 substitution because of an A to G transition at CHR6:86208960 in exon 4 (Caroli et al., 2001; Yahyaoui et al., 2001). This variant occurs at a frequency of less than 0.40 in Italian and German goats and Ardi goats from Saudi Arabia (Supplementary Table S6).
CSN3*D′ differs from CSN3*D by a synonymous mutation in codon 56 and occurs at a frequency of 0.01 in Girgentana goats from Italy (Gerlando et al., 2015).
The CSN3*E protein variant differs from the reference by the amino acid substitution of Asp90 by Gly90 as the results of an A to G transition at position CHR6:86209098 in exon 4 (Angiolillo et al., 2002). CSN3*E occurs in Montefalcone goats from Italy at a frequency up to 0.45 (Angiolillo et al., 2002; Yahyaoui et al., 2003).
CSN3*F differs from the reference protein in the amino acid substitution of Ser159 by Pro159 which occurred due to the T to C transition at CHR6:86209304 in exon 4 (Yahyaoui et al., 2003). This variant was detected in breeds from Italy at a frequency below 0.15 and in Wild Spanish at a frequency of 0.98 (Yahyaoui et al., 2003). The variants CSN3*F′ and CSN3*F encode the same CSN3*F protein. CSN3*F differs from CSN3*F′ by a synonymous mutation in codon 77. CSN3*F′ occurs at a low frequency (0.01) in Raigarh goats from India (Gautam et al., 2019).
CSN3*G is characterized by G to A and T to C mutations at CHR6:86209022, and CHR6:86209304 in exon 4, respectively, in comparison to the reference variant CSN3*B. The mutations cause the amino acid exchange Val65 to Ile65 and Ser159 to Pro159, respectively (Yahyaoui et al., 2003). An addition synonymous mutation is present in codon 43 (Yahyaoui et al., 2003). CSN3*G was found at a frequency of less than 0.15 in a few breeds from Italy and Hair and Angora goats from Turkey (Yahyaoui et al., 2003; Prinzenberg et al., 2005; Gigli et al., 2008; Gerlando et al., 2015). Variant CSN3*G′ has the same amino acid substitutions as variant CSN3*G, but does not contain the synonymous mutation in codon 43 (Supplementary Table S5). The name CSN3*G′ was introduced by Gautam et al. (2019); before that this variant had been named CSN3*F or CSN3*L (Jann et al., 2004; Kiplagat et al., 2010). CSN3*G″ contains the same amino acid substitution as variant CSN3*G and in addition two synonymous mutations in codon 43 and 56 (Vacca et al., 2014; Gerlando et al., 2015; Gautam et al., 2019). The name CSN3*G″ was also introduced by Gautam et al. (2019) before this variant had been named CSN3*N and CSN3*S (Vacca et al., 2014; Gerlando et al., 2015).
CSN3*H differs from CSN3*B in the amino acid substitution from Asn53 to Ser53 due to the transition of A to G in exon 4 at CHR6:86208987 (Jann et al., 2004).
CSN3*I varies from the reference in the amino acid substitution Val65 to Ile65 as a result of a G to A transition at CHR6:86209022 in exon 4 (Jann et al., 2004).
CSN3*J varies from the reference CSN3*B in the substitution of the amino acid Tyr61 to Cys61 because of the transition of A to G in exon 4 at CHR6:86209011. (Jann et al., 2004).
CSN3*K distinguishes from the reference variant by the amino acid substitution Gln44 to Arg44 that is caused by an A to G transition at CHR6:86208960 in exon 4 (Jann et al., 2004). CSN3*K and CSN3*I were found at a frequency ≤0.10 in Bunte and Weise Deutsche Edelziege, and Buren goats from Germany (Kupper et al., 2010). In addition, CSN3*K was detected in Taggar and Saanen goats from Sudan (Rahmatalla et al., 2021).
CSN3*L contains the same amino acid substitution Gln44 to Arg44 that occurs in CSN3*K. The additional substitution of Ser159 by Pro159 differentiate CSN3*L from the reference. This substitution to Pro159 has been detected before in the other protein variants CSN3*C, CSN3*D, CSN3*F, and CSN3*G (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*L had been named before as CSN3*D (Kiplagat et al., 2010). This protein variant was found in Afar goats from Ethiopia only at a frequency of 0.05 (Kiplagat et al., 2010).
CSN3*M (provisionally named CSN3*Y) differs from the reference protein by three non-synonymous mutations. CSN3*M is characterized by a G to A transition at CHR6:86209097 resulting in the amino acid exchange of Asp90 to Asn90, a T to C transition at CHR6:86209263 resulting in a Val145 to Ala145 substitution, and a T to C transition at CHR6:86209304 resulting in the amino acid change from Ser159 to Pro159 (Chessa et al., 2003; Prinzenberg et al., 2005). CSN3*M occurs at a frequency of ≤0.05 in few Italian goats and Red Sokoto goats from Cameroon and Nigeria (Supplementary Table S6) (Chessa et al., 2003; Prinzenberg et al., 2005; Sacchi et al., 2005; Caroli et al., 2007; Gigli et al., 2008; Vacca et al., 2014). CSN3*M′ (previously named CSN3*N) differs from the variant CSN3*M by a synonymous mutation in codon 156 (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*M′ occurs at a frequency of 0.01 in Short Eared goats from Somalia (Kiplagat et al., 2010).
CSN3*N, named before as CSN3*M, carries the two amino acid substitutions Asp90 to Asn90 and Val145 to Ala145 (Kiplagat et al., 2010; Gautam et al., 2019).
The CSN3*O protein variant (previously CSN3*Q) carries the substitutions of Val65 to Ile65, Ile119 to Val119 and Ser159 to Pro159 compared to the reference (Kiplagat et al., 2010; Gautam et al., 2019). CSN3*O and CSN3*N found in a few goats from Africa at a frequency of less than 0.05 and 0.15, respectively (Kiplagat et al., 2010; Rahmatalla et al., 2021).
The replacements of Gln44 to Arg44 and Val65 to Ile65 lead to the variant CSN3*P (Gautam et al., 2019).
CSN3*Q carries the substitutions Gln44 to Arg44, Ile119 to Val119 and Ser159 to Pro159, that had been detected and described before in other k-casein variants. Furthermore, CSN3*Q has the amino acid substitution Thr82 to Ala82 which is the result of an A to G transition at CHR6:86209073 in exon 4 (Gautam et al., 2019). CSN3*Q and CSN3*P were found at a frequency of 0.01 in Blank Bengal Indian goats (Gautam et al., 2019).
CSN3*R distinguishes from the reference in the amino acids Pro80 to Leu80 and Ile119 to Val119. The exchanges from Pro80 to Leu80 and Ile119 to Val119 result from a transition of C to T and A to G in exon 4 at CHR6:86,209,068 and CHR6:86209184, respectively (Gautam et al., 2019).
CSN3*S is distinguished from the reference by the amino acid substitutions Val67 to Ala67 and Ile119 to Val119. The substitution of Val67 to Ala67 is a result of a T to C transition at CHR6:86209029 in exon 4 (Gautam et al., 2019). The frequency of CSN3*S and CSN3*R is about 0.01 in Siroh goats from India (Gautam et al., 2019).
CSN3*T carries also the Ile119 to Val119 substitution. In addition, the substitution Ser37 to Arg37 is present which results from an A to C transversion at CHR6:86208938 in exon 4 (Gautam et al., 2019).
CSN3
留言 (0)