
記住我
Immunotherapy is playing an increasingly important role in the treatment of cancers. This has simulated heightened interest in the identification of additional novel immunotherapeutic targets. CD73 is a purine ecto-5’-nucleotidase with multiple functions playing an important role in a range of biological processes.1 2 In addition to its role in dephosphorylation of purine and pyrimidine nucleotides into corresponding nucleosides, CD73 has been shown to be involved in cell adhesion and migration.3 4 CD73 also has been shown to function as a co-stimulatory molecule for T cells.5 6 Human peripheral blood T cells can become activated by treatment with anti-CD73 antibodies, in combination with suboptimal engagement of the T cell receptor.7 8 CD73 is expressed on subsets of human CD4POS and CD8POS T cells, germinal center follicular dendritic cells, and both naïve and class switched memory B cells.9–11 A role for CD73 in B cell maturation has been proposed as reduced CD73 expression on B cells from patients with common variable immunodeficiency correlates with an inability to produce IgG, however, little is known concerning the normal physiological function of this protein.12 13 Like many glycosyl- phosphatidylinositol anchored molecules, CD73 has been shown to transmit activation signals when ligated by antibodies, although a physiological ligand for CD73 has not been identified.6 10 A number of anti-human-CD73 antibodies have been produced and were selected based on their inhibition of CD73 enzymatic activity.14–17 Some of these antibodies are being evaluated as immunotherapies in cancer clinical trials aimed at reducing intratumor levels of immunosuppressive adenosine.18–20
Here, we report both the characterization of mupadolimab, and its in vivo effects in a phase 1 dose-escalation trial in patients with advanced cancer (NCT03454451). Mupadolimab is a humanized anti-CD73 monoclonal antibody that activates CD73POS B cells and inhibits CD73 enzymatic activity. In vitro, mupadolimab induces the increased expression of markers associated with B cell maturation and antigen presentation, morphological transformation to plasmablasts, and increased secretion of IgM and IgG. Immunophenotypic analysis of peripheral blood from patients with advanced cancer receiving mupadolimab also revealed evidence of B cell activation, clonal expansion and development of memory B cells. The involvement of CD73 in B and T cell activation combined with its role in modulating tissue adenosine levels makes this an attractive therapeutic target. Our findings indicate that the immunomodulatory properties of mupadolimab may be applied to the treatment of cancers, infectious diseases, and potentially in combination with vaccines to enhance immunity in healthy subjects.
MethodsAntibodiesMupadolimab was engineered by isolating VH and VL regions from the parental hybridoma generated by immunizing mice with human CD73 and screening for inhibition of CD73 activity. Humanization was performed by inserting CDRs isolated from the hybridoma into a human framework and the antibody was expressed as a human IgG1-k antibody with the N297Q mutation introduced into the CH2 region to eliminate FcγR binding. MEDI9447 (now oleclumab) was cloned using the VH and VL chain sequences published in WO 2016/075099 AI application patent and was expressed as a human IgG1-l-TM antibody. Both antibodies were expressed in Expi-293 cells (Thermo Fisher Scientific, Cat#14635) and purified by Protein A chromatography (HiTrap Protein A, GE Healthcare Life Sciences). For flow cytometry analysis, antibodies were labeled with AlexaFluor 647 (Life Technologies). Human IgG1 isotype control used for in vitro and vivo studies was purchased from Sigma-Aldrich and BioXCell, respectively. Fab fragments were prepared using the Pierce Fab Preparation kit (Thermo Fisher Scientific). Anti-CD73 antibody clone AD2 was purchased from Abcam.
Cell linesMDA-MB-231 was purchased from ATCC (ATCC Cat# HTB-26MET, RRID:CVCL_VR67). For complete knock out of CD73 expression, CRISPR technology was employed with plasmids obtained from Santa Cruz Biotechnology.
Reagents5' adneosine monophosphate (5’-AMP), adenosine 5’-(α,β-methylene) diphosphate (APCP), and 5’-(N-Ethylcarbox-amido) adenosine (NECA) were purchased from Sigma-Aldrich. Anti-CD3 (clone HIT3a) and anti-CD28 (clone CD28.2) were purchased from BD Biosciences. Recombinant CD73-His was produced in 293-6E cells (RRID:CVCL_HF20) and purified by Ni-NTA (Novagen) and size exclusion Superdex 200 (GE Healthcare) chromatography. Anti-IgM microbeads were purchased from Miltenyi Biotec and were used at a 1:100 dilution.
Affinity measurementsBinding experiments were performed on Octet HTX at 25°C. Purified antibodies (2 µg/mL) were loaded onto Anti-Human IgG Fc Capture biosensors. Loaded sensors were dipped into a threefold dilution series of antigen (starting at 300 nM). Kinetic constants were calculated using a monovalent (1:1) binding model.
Enzymatic activity assaysTo assess the ability of anti-CD73 antibodies to inhibit CD73 activity, cells were incubated with antibodies prior to addition of 250 mM AMP. Phosphate levels were measured in the cell culture supernatant using the Sensolyte Malachite Green assay kit (AnaSpec, Cat# AS-71103). CD73 activity was also measured using the CellTiterGlo assay kit (Promega, Cat#G7571) as previously described.21 22 A cell free reaction was used to determine maximum inhibition of the CellTiterGlo reaction by AMP.
Cryo-electron microscopy methods and image processingCD73:mupadolimab complexes were formed by mixing the protein and antibody at different mole ratios, and purified by size exclusion chromatography for transmission electron microscopy (TEM) imaging. The complex sample was diluted and a 3 uL aliquot was applied to thin, carbon-coated 200-mesh copper grid (Cu-200CN, Pacific Grid-Tech) that has been glow discharged. Electron microscopy was performed using a 300 kV Titan Krios with K3 direct electron detector (Gatan, USA) in counting mode using the automated data collection program SerialEM (RRID:SCR_017293). A typical defocus range was set between −1.0 to −2.0 um. Each micrograph was recorded with a pixel size of 1.08 Å and the electron dose was at 50e/Å2. A total of 3308 micrographs were collected and then used in motion correction and CTF calculation by CTFFIND-4.1. The particles were picked up to create an initial particle stack and then used in two-dimensional classification. Total 605,141 particles were used in 3D classification and finally 254,737 particles were used in 3D model building and refinement, resulting in an overall resolution of 3.07 Å.
Flow cytometryBinding of unlabeled recombinant human antibodies was detected with PE anti-human secondary (Jackson Immunoresearch) and analyzed using the CytoFLEX flow cytometer (Beckman Coulter). For competition experiments, MDA-MB-231 cells (ATCC Cat# HTB-26MET, RRID:CVCL_VR67) were preincubated with APCP for 30 min at 37°C prior to staining with unlabeled mupadolimab or MEDI9447 and detection with secondary antibody. Antibodies to CD3 (BD Biosciences Cat# 562426, RRID:AB_11152082), CD19 (BD Biosciences Cat# 555415, RRID:AB_398597), CD73 (BD Biosciences Cat# 561254, RRID:AB_10894209), CD69 (BD Biosciences Cat# 555531, RRID:AB_395916) were purchased from BD Biosciences. Antibodies to MHC-II (BioLegend Cat# 307606, RRID:AB_314684), CD86 (BioLegend Cat# 305405, RRID:AB_314525), CD83 (BioLegend Cat# 305326, RRID:AB_2561775) were purchased from Biolegend.
For analysis of ERK phosphorylation, human PBMC were stimulated with antibody for 15 min at 37°C. Following stimulation, cells were fixed with 1.6% PFA for 10 min at 37°C. Cells were resuspended in ice cold methanol and stored at −80°C. Cells were washed twice in FACS buffer, treated with Fc block (BioLegend Cat# 422302, RRID:AB_2818986) and stained with an antibody cocktail containing antibodies to: CD20, CD8, CD4, and pERK. Cells were acquired on the CytoFLEX flow cytometer. Antibodies to CD20 (BD Biosciences Cat# 563346, RRID:AB_2738150), CD8 (BD Biosciences Cat# 563919, RRID:AB_2722546), and CD4 (BD Biosciences Cat# 560649, RRID:AB_1727475) were purchased from BD Biosciences. Antibody to phospho-ERK was purchased from Cell Signaling Technologies (Cat# 13214, RRID:AB_2798151).
Chemokine analysisCCL2, CCL3, CCL4, and CCL22 levels were measured using Quantikine ELISA kits (R&D Systems).
T cell functional assaysPBMCs were labeled with Cell Trace Violet (Thermo Fisher). Labeled PBMCs were reconstituted in RPMI 1640 media with 10% FBS and 200 units/mL IL-2 and seeded at a density of 6×105 cells/well in 96 well round bottom tissue culture plates. Anti-CD3 (BD Biosciences Cat# 555336, RRID:AB_395742) and anti-CD28 (BD Biosciences Cat# 555725, RRID:AB_396068) were added to each well at a final concentration of 1 µg/mL. Mupadolimab, MEDI9447, or human IgG1 isotype control was added to each well at the final concentrations indicated in the figure legends. 5’-AMP was added to each well at final concentration of 3 mM and cells were incubated for 4 days at 37°C, 5% CO2.
For flow cytometry analysis of T cell proliferation, duplicate wells were pooled and stained with PerCP-Cy5.5 anti-CD3 (BD Biosciences Cat# 552852, RRID:AB_394493). After staining, cells were washed and fixed with 4% PFA and analyzed using the CytoFLEX flow cytometer. Data were analyzed with FlowJo software (RRID:SCR_008520) and proliferating CD3+T cells were identified by gating relative to control cells not stimulated with anti-CD3/anti-CD28 or AMP. To compile data derived from multiple donors, experimental values were normalized to the value derived from cells stimulated with anti-CD3/anti-CD28 in the absence of AMP for each donor.
To evaluate cytokine secretion, experimental cultures were set up as described above except that cells were seeded at a density of 1.5×105 cells/well and labeling with Cell Trace Violet was omitted. Interferon gamma (IFN-γ) levels were measured using the human IFN-γ AlphaLISA detection assay kit (Perkin Elmer, AL217C). Data from multiple donors were compiled as described above.
Preclinical mouse immunization modelNSG-SGM3 mice (Jackson Laboratories, stock #013062) reconstituted with human CD34POS cells from the cord blood were immunized with an emulsion of 50 ug full length spike protein from SARS-CoV-2 (ABClonal, Cat#RP01260MT) or 50 ug influenza A H1N1 (A/California/07/2009, Sino Biological Cat#11085-v08H) HA protein, mixed with Freund’s Incomplete Adjuvant (Sigma, Cat#77145) subcutaneously on both the left and right flank (25 µg/side). Mice (n=8) were then randomized into two treatment groups: half the animals were dosed daily (i.p.) with 10 mg/kg mupadolimab, and half the animals were dosed daily with 10 mg/kg hIgG1 (BioXCell, #BE0297). Animals were cheek bled on days 1 (pre-bleed), 4, 8, 11 and 15 to assess anti-spike or anti-nucleocapsid (as a negative control) antibody production over time by ELISA following a 1:50 dilution of serum. Anti-influenza antibody production was assessed by MSD on days 1 (pre-bleed), 4, 6, and 8.
Immunophenotyping of mupadolimab treated patient PBMCsFresh blood from patients with cancer was analyzed by flow cytometry. Expression of cell surface markers associated with B cell and T cell activation were assessed using Fc blocking reagent (Miltenyi Biotech, Catalog #130-059-901) and antibodies directed to CD19 BV421 (Clone HIB19; BD Biosciences, Cat #562440), CD73 PE (Clone AD2; BD Biosciences, Cat #550257), CD3 PE-Cy7 (Clone UCHT1; BioLegend, Cat #300420), CD27 APC (Clone L128; BD Biosciences, Cat #337169), Mouse anti-human CD45RA APC-Fire 750 (clone HI100, BioLegend, Catalog #304152), Mouse anti-human CD8 BVD650 (clone SK1, BD Biosciences, Catalog #565289), Mouse anti-human CD4 PE-CF594 (clone SK3, BD Biosciences, Catalog #566914), Rat anti-human CXCR5 APCR700 (clone RF8B2, BD Biosciences, Catalog #565191). Flow cytometry data was analyzed using FlowJo V.10.7.
B cell receptor analysisSequencing of the CDR3 regions of human variable chains was performed using the immunoSEQ B cell receptor (BCR) Assay (Adaptive Biotechnologies, Seattle, Washington, USA). Genomic DNA was extracted from peripheral blood mononuclear cells (PBMCs) and was amplified in a bias-controlled multiplex PCR, followed by highthroughput sequencing and the abundance of each unique BCR region was quantified (37–39).
Clinical trial designClinical results are reported from the completed portion of an open label, multicenter, dose-escalation study of the safety, tolerability and pharmacokinetics of single agent mupadolimab in patients with advanced cancer (NCT03454451).
Blood was collected prior to treatment initiation and at 0.5 hours, 2 hours, 24 hours, 7 days, and 21 days postinfusion. Mupadolimab concentration was determined with an electrochemiluminescent immunoassay at Charles River Lab. B cells were used to assess of CD73 target occupancy with mupadolimab through validated flow cytometry assay. The total CD73 and free CD73 (CD73 that is not bound by mupadolimab) was stained with PE-labeled AD2 clone (BD Biosciences, Cat #550257) and AF488-labeled 1E9 clone (in-house), respectively.
All investigators participating in the trial were required to disclose potential conflicts of interest.
Statistical analysisStatistical analysis was performed using GraphPad Prism V.8. In all the figures, Error bars represent mean±SD *p<0.05, ***p<0.001, and ****p<0.0001 as determined by t-test.
ResultsMupadolimab inhibits CD73 enzyme function and reverses adenosine-mediated immunosuppressionMupadolimab is a humanized anti-CD73 IgG1κ antibody engineered with an N297Q mutation in CH2 of the heavy chain to eliminate Fc effector functions such as the ability to fix complement and initiate antibody dependent cellular cytotoxicity (ADCC, online supplemental figure S1A). Mupadolimab binds human CD73 with a KD of 200 pM measured using biolayer interferometry (online supplemental figure S1B).
The ability of this antibody to inhibit the enzymatic activity of cellular CD73 was evaluated by measuring free phosphate levels generated on conversion of AMP to adenosine. Mupadolimab inhibited CD73 activity in the human breast cancer cell line MDA-MB-231 to a baseline level that was established by comparison to both the CRISPR CD73 knockout cell line and comparison to treatment of cells with saturating amounts of APCP, a small molecule inhibitor of CD73 enzymatic activity (figure 1A). In contrast, MEDI9447, another anti-CD73 monoclonal antibody that binds a non-overlapping epitope, only partially reduced CD73 activity.21 Mupadolimab also eliminated the enzymatic activity of CD73 expressed on primary human PBMCs while MEDI9447 demonstrated a partial effect on enzymatic activity (figure 1B). These data show that mupadolimab completely inhibits the catalytic activity of cellular CD73 expressed by tumor cells and normal lymphocytes. Unlike that of membrane-bound CD73, the enzymatic activity of soluble CD73 is not inhibited by mupadolimab binding compared with MEDI9447 (online supplemental figure S2A,B) suggesting differences in structural conformation between membrane-bound and soluble CD73.
 Figure 1
Figure 1 Mupadolimab blocks CD73 enzymatic activity and restores T cell proliferation and cytokine production in an AMP-mediated immunosuppressive environment. (A) CD73 catalytic activity was measured with MDA-MB-231 cells or MDA-MB-231 CD73 CRISPR cells in the presence of mupadolimab, MEDI9447, or 1 mM APCP by adding 250 mM AMP and measuring phosphate levels in the cell culture supernatant. (B) Human PBMCs were incubated with AMP in the presence of mupadolimab, MEDI9447, or isotype control antibody at the saturating concentration of 300 nM. After 4 hours of incubation, the AMP remaining in the supernatant was quantified using the CellTiterGlo assay as described in the methods. Percentage of inhibition was plotted. Data represent nine individual donors and error bars represent mean±SD ****p<0.0001 as determined by t-test. (C) T cell proliferation for 13 donors and (D) IFN-γ production for 6 donors treated with 500 nM mupadolimab or MEDI9447 or 890 nM isotype control. Data for (C) and (D) normalized across donors. Error bars represent mean±SD *p<0.05, ***p<0.001, ****p<0.0001 as determined by t-test. APCP, adenosine 5’-(α,β-methylene) diphosphate; PBMCs, peripheral blood mononuclear cells.
To model the immunosuppressive effects of adenosine on T cells, human PBMCs were obtained from healthy donors and cultured under T cell activating conditions using anti-CD3 and anti-CD28 stimulation. Addition of AMP resulted in decreased T cell proliferation and cytokine secretion. Mupadolimab restored T cell proliferation in all donors tested, consistent with the antibody preventing conversion of AMP into immunosuppressive adenosine (figure 1C). MEDI9447 restored T cell proliferation in a subset of donors but did not reach the magnitude of the mupadolimab response. IFNγ production was evaluated as an additional indicator for T cell response and similar results were observed (figure 1D). These data demonstrate that mupadolimab inhibits CD73 enzymatic activity and blocks the suppressive effects of adenosine on T cell proliferation and cytokine secretion.
Characterization of the mupadolimab binding siteA structural analysis of mupadolimab-CD73 complexes was performed using cryogenic TEM (cryo-TEM) at a resolution of 3.07 Å. The CD73 homodimer binds with two mupadolimab antibodies to form a circular granule at a closed conformer (figure 2A).23 A total of 17 hydrogen bonds, 2 π bonds and 2 salt bridges contribute to the antibody-CD73 interaction (figure 2B). The mupadolimab binding epitope is positioned at the N-terminal and adjacent to substrate binding site when CD73 is in closed conformation (figure 2C). The antibody binds to epitope amino acid residues at the 205–301 positions. Consistent with these structural findings, mupadolimab was found to compete with substrate binding to CD73 as shown in online supplemental figure S3 using APCP, a non-hydrolyzable analog of AMP.
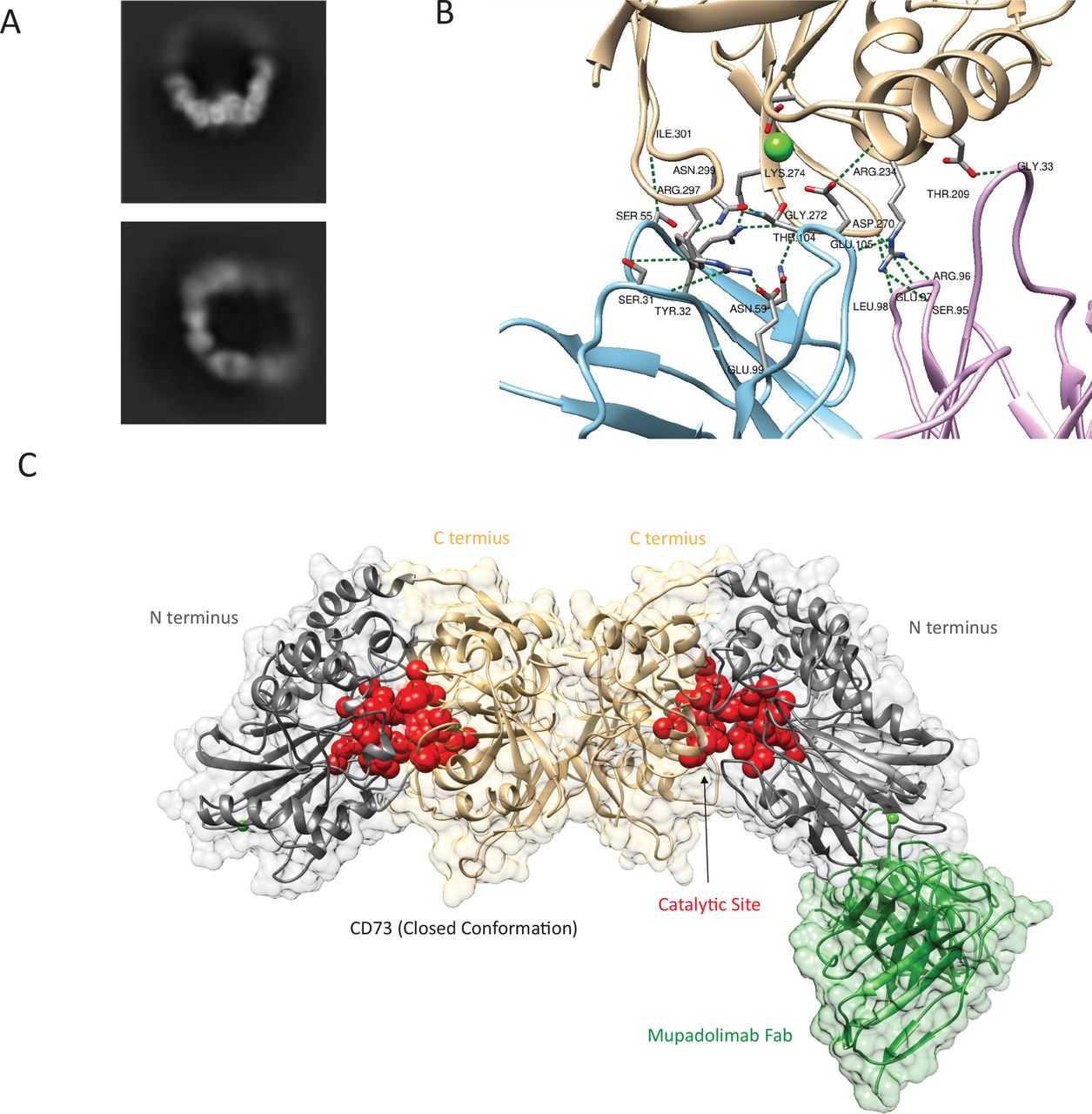 Figure 2
Figure 2 Cryo-TEM analysis of CD73 and mupadolimab complex. (A) Represented 2D classification results of CD73-mupadolimab 2:2 complex. (B) The interaction between mupadolimab CDR and CD73. The heavy chain and light chain of CDR are shown in is blue and pink, respectively. CD73 is colored in wheat. The calcium ions are shown as green sphere. (C) 3D models of CD73 and the Fab region of the antibody based on density map. The N- and C-terminal domain of the CD73 dimer were shown in gray and wheat, respectively. The mupadolimab Fab is shown in green. Residues involved in the catalytic site of CD73 are shown in red. Due to the high flexibility and low density of the half of the structure, the constructed model only contains one Fab molecule.
Mupadolimab directly activates human B cells in vitroCD73 is expressed on subsets of human hematopoietic cells and has previously been implicated to play a role in lymphocyte activation and adhesion.3 10 11 24 We performed a flow cytometry-based screen to identify differentially expressed cell surface markers on immune cells following in vitro treatment with mupadolimab. Mupadolimab strikingly activated B lymphocytes, resulting in the upregulation of activation markers (CD69 and CD83) and antigen presentation machinery (CD86 and MHC-II) to similar levels achieved with the positive control of BCR crosslinking via anti-IgM (figure 3A). Mupadolimab-mediated B cell activation also resulted in increased cell surface expression of CD27, IgG, CD38, and CD138, all markers consistent with induction of B cell maturation (figure 3B). Activation was dose-dependent with concentrations of 1 µg/mL of mupadolimab achieving near maximal induction of CD69 in vitro (figure 3C). B cell activation has not previously been described in relation to CD73 signaling and is seemingly unique to mupadolimab, as both MEDI9447 and clone AD2, two anti-CD73 antibodies that do not cross-block mupadolimab, fail to induce activation (figure 3C). Naïve peripheral blood B cells stimulated with mupadolimab in vitro underwent morphological changes consistent with differentiation into plasmablasts (figure 3D). Mupadolimab-mediated B cell activation was restricted to CD73POS B cells and required bivalent binding as mupadolimab immunoglobulin Fab fragments had minimal effect on expression levels of CD69 (figure 3E). Mupadolimab induction of CD69 was blocked with ibrutinib, a covalent BTK inhibitor (figure 3F), and mupadolimab induced the phosphorylation of ERK selectively in B cells similar to BCR stimulation with anti-IgM (figure 3G), demonstrating that mupadolimab directly activates B lymphocytes through canonical B cell signaling pathways. B cell activation was not a consequence of the concentration of extracellular adenosine as addition of a broad range of concentrations of NECA, a potent and stable agonist for adenosine receptors, did not block the induction of activation markers by mupadolimab (figure 3H).25 Data from additional experiments evaluated gene expression in normal B cells stimulated with either anti-IgM or mupadolimab and revealed no differences in gene expression patterns (data not shown).
 Figure 3
Figure 3 Mupadolimab directly activates B lymphocytes and induces maturation into antibody secreting plasmablasts in vitro. (A) Purified B cells from 3 to 5 healthy donors were incubated overnight with human IgG1 isotype control or mupadolimab (10 μg/mL) or anti-IgM microbeads, a positive control for BCR stimulation. Expression of activation markers CD69, CD83, CD86, or MHC-II was measured on viable cells by flow cytometry. Error bars represent mean±SD *p<0.05, **p<0.01, and ****p<0.0001 as determined by t-test. (B) Time dependent increases in the expression of CD27, IgG, IgM, CD38, and CD138 on purified B cells cultured in the presence of mupadolimab or isotype control (1 µg/mL). Mean fluorescence intensity (MFI) was determined for each marker and was normalized to the untreated control for each donor. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 as determined by two-way ANOVA. (C) B cell activation is unique to mupadolimab as other anti-CD73 antibodies do not induce CD69 expression. Expression of CD69 (MFI) was measured by flow cytometry. (D) Representative images of purified B cells cultured with isotype control (left panel) or mupadolimab for 2 days. (E) Purified B cells were incubated overnight with 10 μg/mL human IgG1 isotype control or mupadolimab or equimolar mupadolimab Fab. CD69 and CD73 were measured on B cells by flow cytometry. (F) Human PBMCs were incubated overnight with a fixed concentration of mupadolimab (10 μg/mL) along with ibrutinib or vehicle control over a range of concentrations. Expression of CD69 on B cells (CD19POSCD3NEG) was measured by flow cytometry. (G) Human PBMCs from healthy donors (n=2) were incubated for 15 min with 10 μg/mL human IgG1 isotype control, mupadolimab, or anti-IgG+anti IgM, a positive control for BCR stimulation. Phospho-ERK was measured by flow cytometry on B cells, CD4+T cells, and CD8+T cells. (H) Human PBMCs were incubated overnight with 10 μg/mL mupadolimab with or without NECA over a range of concentrations. expression of CD69 on B cells (CD19POSCD3NEG) was measured by flow cytometry and MFI is reported. ANOVA, analysis of variance; BCR, B cell receptor; NECA, 5’-(N-Ethylcarbox-amido) adenosine.
To assess functional consequences of B cell activation with mupadolimab, we first measured the concentration of IgG and IgM secreted by healthy donor PBMCs into the culture supernatant 6 days after in vitro treatment. The addition of mupadolimab resulted in a 3-fold increase in IgM and IgGλ levels (IgGκ not measured since mupadolimab is an IgGκ) relative to an isotype control, demonstrating that mupadolimab stimulates antibody secretion and possibly class switching to IgG (figure 4A). Mupadolimab also induced the production of B cell cytokines such as CCL3, CCL4, CCL2, and CCL22 (figure 4B–E), but did not induce IFNγ, IL-2, IL-6, IL-10, or TNFα from healthy donor PBMCs (data not shown). Collectively, these experiments demonstrate that mupadolimab activates B lymphocytes, resulting in morphological and immunological changes consistent with B cell differentiation and antibody secretion, and this property is independent of adenosine-modulatory activity.
 Figure 4
Figure 4 Mupadolimab activates human B cells to secrete immunoglobulin and cytokines associated with B cell activation. (A) Human PBMCs were incubated for 6 days with 1 mg/mL human IgG1 isotype control or mupadolimab. IgG1 and IgM secreted into the culture supernatant was quantified by ELISA. IgG-k was not measured as addition of mupadolimab, an IgG1-k antibody, would have confounded the results. Data are represented as mean±SEM of 6 independent experiments, each in duplicate. Purified B cells were incubated for 5 days with 10 µg/mL human IgG1 isotype control or mupadolimab or anti-IgM microbeads, a positive control for BCR stimulation. Concentration of CCL2 (B), CCL3 (C), CCl4 (D) and CCL22 (E) in supernatant were measured by ELISA. Two representative donors are reported and data are represented as mean±SD of duplicate measurements. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 as determined by two-tailed, unpaired t-test.
Mupadolimab enhances antigen-specific humoral immunity in a humanized mouse modelMupadolimab does not cross-react with murine or rat CD73. Therefore, we used NSG-SGM3 mice reconstituted with human cord blood to determine if mupadolimab could enhance antigen-specific immune responses to SARS-CoV-2 spike protein and to influenza hemagglutinin. Though these mice have no mouse B, T, or NK cells, they have been reconstituted with a human immune system that does contain human B cells that can be activated with mupadolimab.26 27 These mice were immunized with either purified SARS-CoV-2 trimeric spike protein or with influenza hemagglutinin in incomplete adjuvant, along with mupadolimab or a human IgG1 isotype control. Mice vaccinated with the spike protein plus mupadolimab produced antigen-specific human antibodies, while mice receiving spike protein with an isotype control did not mount a response (figure 5A). These antibody responses were antigen specific, as mice treated with mupadolimab made antibodies to the immunizing spike protein; but not to the control nucleocapsid SARS-CoV-2 viral protein (figure 5B). Similar results were obtained with the influenza hemagglutinin antigen, although low levels of antibody response were seen in the control group compared with the animals receiving mupadolimab (figure 5C). This could be explained by pre-existing immunity from cells transferred in the donor cord blood. These results demonstrate that mupadolimab stimulates antigen specific humoral responses and would be expected to potentiate antibody responses to viral and possibly other antigens.
 Figure 5
Figure 5 Mupadolimab enhances antigen-specific responses in preclinical animal model. NSG-SGM3 mice were immunized with an emulsion of full-length spike protein from SARS-CoV-2 plus Freund’s incomplete adjuvant. Mice were then randomized into two treatment groups: half the animals (n=4) were dosed daily (i.p.) with 10 mg/kg mupadolimab, and half the animals (n=4) were dosed daily with 10 mg/kg hIgG1. Animals were cheek bled on days 1 (pre-treatment), 4, 8, 11, and 15 to assess anti-spike immunoglobulin (A), anti-nucleocapsid immunoglobulin (B) as negative control overtime by ELISA. (C) NSG-SGM3 mice were immunized with an emulsion of influenza H1N1 hemagglutinin plus Freund’s incomplete adjuvant. Anti-influenza antibody level was assessed overtime by MSD platform after treatment with mupadolimab, hIgG1, or PBS. Error bars represent mean±SD. P values are shown whenever results achieved statistical significance. PBS, phosphate buffered saline.
In vivo immunological effects of mupadolimab in patients with advanced cancerMupadolimab is being evaluated as an immunotherapy for cancer in an ongoing phase 1 study (NCT03454451). The dose escalation, monotherapy portion of the study has been completed and followed a traditional 3+3 design, with cohorts of patients receiving mupadolimab administered over 30–60 min by intravenous infusion once every 21 days. Thirty-four patients were assigned sequentially to cohorts to receive doses of 1, 3, 6, 12, 18, or 24 mg/kg mupadolimab. Patient characteristics were: median age 64 years (range 46–79); 77% male; and median prior therapies were 4 (range 1–6). Cancer histologies included bladder, colorectal, head and neck, prostate, non-small cell lung, pancreatic, renal cell, triple negative breast and sarcoma.
The serum pharmacokinetics and occupancy of CD73 sites present on circulating CD73POS lymphocytes were evaluated. Exposure of mupadolimab increased with increasing doses up to 24 mg/kg (online supplemental figure S4A). Substantial occupancy of target sites at day 21 serum trough levels of mupadolimab was achieved with doses of ≥6 mg/kg (online supplemental figure S4B).
We have evaluated the in vivo immunological effects in 22 patients receiving doses up to 12 mg/kg, and observed rapid decreases in circulating CD73POS B cells, 30 min after antibody infusion (figure 6A). Reductions in circulating CD73POS B cells was not due to blocking by the administered antibody as reactivity with another non-cross blocking anti-CD73 antibody, AD2, was similarly reduced. Mupadolimab does not induce B cell death or initiate ADCC (online supplemental figure S1A) so this result is likely due to the redistribution of activated, CD69POS B cells into lymphoid tissues. B cells returned to circulation at levels similar to baseline by day 21 (figure 6A). Returning B cells were enriched in CD27POS IgDNEG class switched memory B cells in the majority of subjects receiving ≥3 mg/kg mupadolimab (figure 6B). Memory B cells are essential for both acute and long-term immunity as they have undergone immunoglobulin rearrangement and somatic hypermutation in order to produce high affinity, antigen-specific antibody on exposure to antigen. BCR repertoire analysis was performed in three patients and revealed the emergence of novel B cell clones that were not present in the peripheral blood prior to treatment (figure 6C). These new B cell clones were limited to 2–40 clones per patient and appeared with frequencies as high as 1:100 B cells, consistent with a robust clonal expansion following an antigen-specific response. The cognate antigens recognized by these new B cell clones are currently under characterization.
 Figure 6
Figure 6 B cell dynamics in patients with advanced cancer treated with mupadolimab. (A) Mupadolimab induces rapid change in peripheral blood B cells, returning to pretreatment baseline by day 21. Results are shown as fold change from baseline for patients receiving up to 12 mg/kg. Results between 0.5 hour and 21 days were statistically significant at 3 mg/kg (*p<0.05) and 12 mg/kg (**p<0.01). (B) An increased frequency of memory B cells (CD27POS IgDNEG) is observed at doses ≥3 mg/kg. Each symbol represents a treated patient, with lines connecting paired samples. (C) Representative examples of BCR repertoire diversification in 3 patients with cancer 21 days after mupadolimab treatment with 12 mg/kg (RCC patient) or 18 mg/kg (patients with colorectal cancer). The Y axis represents clones increased in frequency and the X axis indicates clones that decreased in frequency. Red dots indicate clones that were significantly expanded or exclusively observed following treatment with mupadolimab. BCR, B cell receptor; RCC, renal cell cancer.
Administration of mupadolimab was well tolerated. Mild to moderate infusion reactions (primarily chills and rigors) seen with the first infusion of mupadolimab at doses of ≥3 mg/kg were mitigated by pretreatment with diphenhydramine and acetaminophen. Treatment-related grade 3 adverse events seen in one patient each included anemia, lymphopenia, transient elevation of aspartate aminotransferase and alkaline phosphatase, and hyponatremia (online supplemental table 1). A maximum tolerated dose was not reached. No changes in serum quantitative immunoglobulins were seen. No objective responses by RECIST criteria were observed, however, tumor regression was seen in patients with non-small cell lung, head and neck, and prostate cancer (figure 7). No significant dose response correlations were observed. Interestingly, patients who exhibited the largest changes in circulating B cells appeared more likely to have reduction in tumor volumes (data not shown). The disease control rate in 34 evaluable patients (confirmed stable disease≥3 months) was 23% with median duration of 6 months. The duration of treatment ranged from 1 to 20 months.
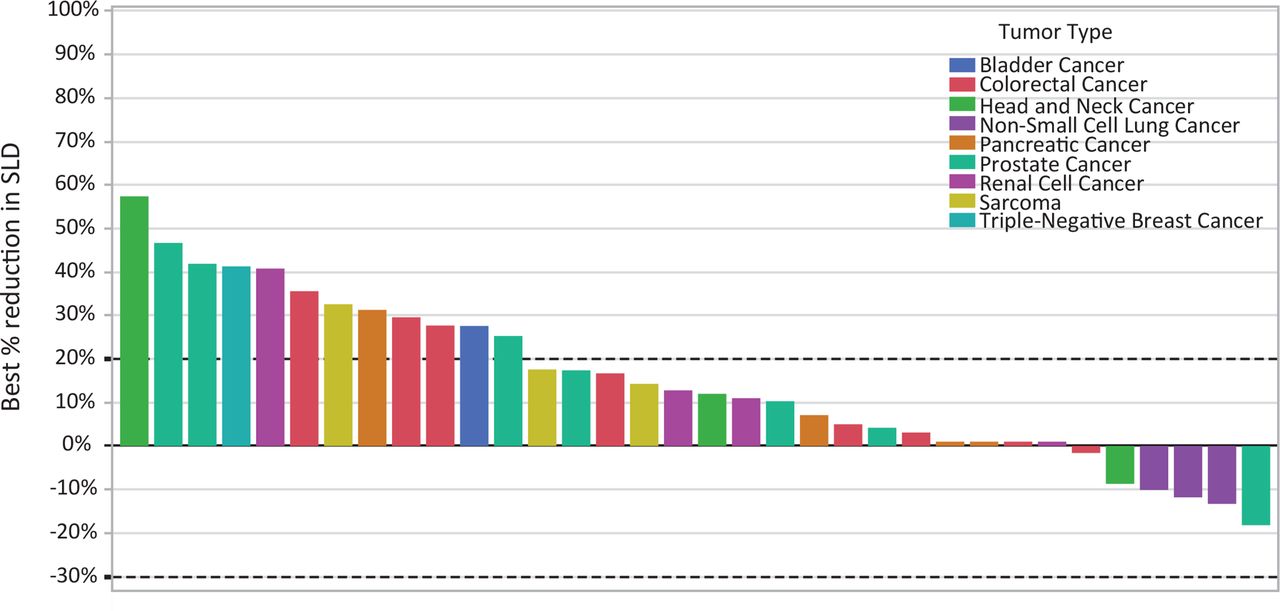 Figure 7
Figure 7 Mupadolimab antitumor activity. The waterfall plot shows the best change in the sum of the longest dimensions (SLD) for each patient.
DiscussionMupadolimab is a humanized FcγR binding-deficient anti-CD73 monoclonal antibody that directly activates CD73POS B cells, thereby inducing their trafficking to lymphoid tissues and promoting antibody production and differentiation into memory B cells. Mupadolimab induces markers of B cell activation (CD69), maturation (CD138, CD38, CD27, IgG), and antigen presentation (CD86, CD83, MHC-II) in vitro along with a corresponding morphological transformation into antibody secreting plasmablasts. Studies in patients with cancer demonstrated that administration of mupadolimab leads to the diversification of the BCR repertoire by stimulating the emergence and expansion of novel B cell clones. Experiments conducted in NSG-SGM3 mice with mupadolimab showed it could enhance antigen specific antibody responses. SARS-CoV-2 and influenza hemagglutinin antigens were evaluated in this model as they represent important, relevant viral antigens. We found that antibody responses to these antigens were enhanced raising the possibility that mupadolimab could be used as an immune adjuvant with vaccines or to enhance humoral immunity to tumor antigens.
These findings indicate that mupadolimab can activate CD73POS B cells to elicit antigen specific humoral and memory responses that display functional hallmarks associated with protective immunity. These effects are independent of CD73 enzymatic activity as addition of adenosine analogs or blockade of CD73 enzymatic activity alone has no effect on B cell activation in vitro. In vitro studies indicated that cross linking of CD73 is involved in B cell activation as monomeric Fab fragments showed markedly reduced activation measured by expression of CD69. BTK inhibition blocked expression of CD69 in cells stimulated with mupadolimab. These findings, together with the demonstration of phosphorylation of ERK, suggest that mupadolimab activation occurs through the BCR signaling pathway.
CD73 was originally characterized as a costimulatory molecule for T cells, but our results demonstrate that mupadolimab predominantly activates B cells.5 6 Human CD73 is expressed on IgDPOSIgMDIM/NEG naïve B cells, and CD27POS memory B cells expressing IgG or IgA.9 Mupadolimab induces the expression of CD69, an activation marker that negatively regulates S1PR1 function, resulting in the prolonged retention of activated B cells in lymphoid organs and thymus.28 This increased lymphoid residence time provides time to complete B cell activation and interact with CD4POS T follicular helper cells to shape downstream immune responses. We also find that mupadolimab induces the secretion of cytokines involved in B cell differentiation including CCL22 (figure 4E). Liu et al have published that CCL22 promotes efficient B cell maturation in germinal centers by increasing the interaction of B cells with T follicular helper cells.29 While B cell activation with mupadolimab is independent of adenosine blockade in vitro, the CD73 enzymatic blockade may be complementary in vivo as adenosine has been shown to restrict lymphocyte migration into lymph nodes in preclinical animal models.30 Additional studies will clarify if this temporary sequestration in lymph tissues is the dominant mechanism by which mupadolimab diversifies the BCR repertoire and promotes the development memory B and T cells.
The findings that mupadolimab induces immune activation together with blockade of the production of immunosuppressive adenosine led us to evaluate monotherapy with this antibody in a phase 1 dose-escalation clinical trial in patients with advanced refractory cancers. Initial in vivo results from this trial are consistent with our in vitro observations. We observed dramatic effects on circulating B cells with prompt reduction of CD73POS cells and slow return to baseline as circulating mupadolimab levels fall. The returning B cells, were primarily CD73NEG. We hypothesize that the CD73POS cells were retained in lymphoid tissues due to expression of CD69. Effects on circulating B cells were seen with relatively low doses of mupadolimab (1 mg/kg), however, this dose achieves plasma concentrations in excess of 1 µg/mL; a level shown to cause maximal expression of CD69 (figure 3C). Mupadolimab was well tolerated at doses up to 24 mg/kg given on a 21-day schedule. B cell activation did not result in changes to serum quantitative immunoglobulins. In other studies with mupadolimab in patients with COVID-19, no alterations of serum antibody titers to common viral antigens were observed.31 32 These results demonstrate that mupadolimab does not induce polyclonal B cell activation and implies a requirement for concomitant antigen exposure in order for mupadolimab to induce an antigen-specific response. In three patients with cancer studied with BCR repertoire analysis, there was evidence of expansion of new B cell clones in the blood following treatment, in some cases reaching frequencies as high as 1% of B cell clones. The antigen specificity of these clones is unknown, although it is interesting to speculate that they could be directed against tumor antigens. It is unlikely that such changes in clonality occur spontaneously, though concomitant exposure to other external antigens is a possibility.
Mupadolimab was well tolerated at all doses tested. Tumor reduction with single agent therapy was observed in several patients and other patients had disease control for significant durations. This phase 1 tr
留言 (0)