1. IntroductionSword bean (Canavalia gladiata) is an important annual legume consumed as a vegetable and as a medicine, and is endemic to tropical Asia, Africa, and India [
1,
2]. It is currently cultivated across 0.065 million hectares in southwestern China [
3,
4]. All parts of the sword bean are edible, such as the beans, leaves, pods, or roots [
5]. Immature sword bean pods are extensively utilized in Asia as a vegetable [
6]. It is widely cultivated across the world, but especially so in central and southern China, where it is an important food supply [
4]. It can increase soil nutrient levels, improve soil structure, and reduce soil erosion [
7]. Sword bean is the third most important medicinal and food crop in southwestern China.The fungal pathogen Colletotrichum causes anthracnose on legume crops globally [
8,
9,
10]. The genus Colletotrichum includes about 600 species that can destroy many crops [
11], and cause typical symptoms such as sunken necrotic lesions, generally known as anthracnose [
12]. Anthracnose is an important disease of sword bean, and Colletotrichum capsica and Colletotrichum lindemuthianum have specifically been detected in sword beans in India and China [
13,
14].Colletotrichum truncatum is a species commonly reported to cause disease in papaya, lentils, and soybeans [
15,
16,
17,
18]. No previous studies have reported any fungi associated with anthracnose disease on sword beans. Therefore, this study aimed to identify the causal agent of anthracnose disease in sword beans by investigating its morphological and molecular properties, as well as to test its pathogenicity. 2. Materials and Methods 2.1. Plant Collection and Fungal IsolationA total of fifteen diseased stems from five plants (three stems per plant) were excised from the margins of lesions. Stem pieces were sterilized with 75% ethanol for 30 s and 1% NaClO for 75 s, rinsed three times with sterile distilled water, dried three times between sterile filter paper, and plated onto potato dextrose agar (PDA) supplemented with 25 mg/L of penicillin and streptomycin, and incubated at 24 °C for 3 to 21 days [
19]. The isolation frequency for each tissue type was determined by examining colony characteristics on the fourth day. Pure colonies were obtained by transferring 1–2 mm emerging hyphal tips onto fresh PDA plates with no supplements. A diseased specimen, as well as three isolates, was deposited at the Mycological Herbarium of the Lanzhou University (MHLZU) with the numbers MHLZU19328, and YN1932501, YN1932502, YN1932503, and YN1932504, respectively. 2.2. Morphological Characterization
Mycelial plugs (5 mm in diameter) were removed from subcultured hyphae colony edges and dark-incubated at 25 °C on PDA. The PDA colony characteristics were recorded after 10 days, and acervuli, setae, and conidia were observed using a stereomicroscope (Nikon ECLIPSE Ti, Tokyo, Japan) and regular microscope and photographed with a Canon DS126391 camera (Canon, Lanzhou, China). Colony diameters and colors were recorded from PDA-grown cultures.
2.3. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and SequencingPure culture mycelia were scraped into centrifuge tubes using a sterilized spoon. Total genomic DNA was extracted from representative isolates using a Fungal DNA Kit (D3195; OMEGA Biotech Co. Ltd., Norcross, GA, USA) following the manufacturer’s instructions. DNA samples were stored at −20 °C for further study. The rDNA internal transcribed spacer (ITS), a partial actin sequence (ACT), glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and histone3 (HIS3) genes were amplified (PCR amplification and sequencing primers are given in
Table 1). PCR reactions were performed in a 2720 Thermal Cycler (Applied Biosystems, Foster City, CA, USA) in a total volume of 25μLthat contained 1 μL of genomic DNA, 1 μL of forward and reverse primers, 12.5 μL of 2 x High-Fidelity Master Mix, and 9.5 μL of ddH2O. The PCR conditions were as follows: an initial denaturation step was performed at 94 °C for 3 min, followed by 30 cycles at 94 °C for 10 s, then 30 s of annealing (54 °C, 56 °C, 46 °C, 56 °C, and 52 °C for ITS, ACT, GAPDH, and HIS3, respectively), and 72 °C for 10 s. A final extension step was performed at 72 °C for 10 min [
20]. 2.4. Phylogenetic AnalysisThe sequences obtained from the four isolates, as well as other Colletotrichum spp. reference sequences as described by Damm [
26], were downloaded from GenBank (
Table 2).
Single sequences were aligned using ClustalW in MEGA 5.1. The four loci were combined with Sequence Matrix 1.8. The best-fit nucleotide substitution models of each gene were assessed by MrModeltest 2.3 for Bayesian reference (BI) analysis. The best-fit model of each gene was imported into MrBayes v. 3.2.6, and the full dataset was run for 2,000,000 generations, and sampled every 100 generations and four chains. The resulting tree was created in Figtree v1.4.3, Adobe Acrobat DC (Adobe, San Jose, CA, USA), and Microsoft Office PowerPoint 2007 (Microsoft, Redmond, CA, USA).
2.5. Pathogenicity Assay
The pathogenicity of C. truncatum was determined by a spray treatment. A total of 100 sword bean seeds were obtained from sword bean plants during the 2020 harvest season, surface-sterilized with 75% ethanol for 30 s and 1% NaClO for 75 s, rinsed three times with sterile distilled water, and transferred into sterilized Petri dishes that contained a double filter paper layer. A total of 5 mL sterile distilled water was added to the seeds, whereafter they were incubated in the dark at 25 °C for 7 days. The fungal strains were cultivated in the dark at 25 °C for 14 days, and the colony surfaces were gently scraped with a glass spreader. The conidial suspension was adjusted to a concentration of 1.0 × 106 conidia/mL sterile water using a hemocytometer. Fifty sword bean plants were transferred into 10 pots (with five plants per pot) that contained 500 g of sterilized soil. The stems of 50 healthy plants were sprayed with a conidial suspension that contained 1.0 × 106 conidia/mL and 0.01% Tween 80. A total of 50 healthy plants were used as controls and received a sterile water spray. All plants were kept in a greenhouse (days: 22 °C, 18 h light; nights: 18 °C, 6 h dark) and were covered with clear polyethylene bags for three days to maintain a high humidity level.
4. Discussion
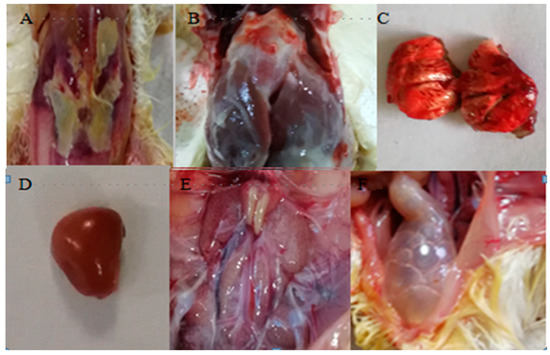
In this study, sword bean was described as a new host for C. truncatum in the Yunnan Province, China. Sunken necrotic lesions were typical symptoms observed on sword bean stems in the field. Koch’s postulates were satisfied since this fungus was (1) isolated from diseased sword bean plants, (2) the successfully inoculated sword bean plants produced the same symptoms, and (3) the fungus was subsequently re-isolated. A combined analysis of the morphological characteristics and multiple gene sequence data further confirmed that all four Colletotrichum isolates were C. truncatum.
Sword bean anthracnose symptoms caused by C. truncatum, as observed in the field, were similar to those caused by C. lentis. The stem lesions were initially dark and linear, but later enlarged and often coalesced with adjacent lesions until large stem areas were involved [
20]. However, C. truncatum anthracnose seemed to cause more and more severe lesions on stems than on leaf laminas, whereas C. lentis caused severe lesions on both stems and leaves. Four other plant species grew in the vicinity, namely Canavalia gladiata, Brachiaria eruciformis, Indigofera amblyantha, and Stylosanthes guianensis, and previous studies have reported that C. truncatum can infect Indigofera plants. It is possible that the infection source for C. truncatum on sword bean might be closely related to the presence of Indigofera amblyantha plants, but this must be verified. Furthermore, the climatic conditions of the area, as well as field planting patterns, may also play a role.Identification based on morphology is a primary step towards classifying fungal pathogens at the genus level [
30]. Corda described the genus Colletotrichum in 1831, which was historically based on morphological characteristics such as fusiformed, curved, hyaline conidia with acute ends, and brown, opaque, subulate setae with acute tips [
31]. The morphology of C. truncatum found in this study was similar to that of previous reports [
26]. However, Colletotrichum species identification based only on morphology is not highly accurate since few morphological characters can distinguish between the species, and the teleomorphic stages are rarely formed [
32]. Moreover, morphological species characteristics can change when environmental conditions change, particularly the small morphological differences among C. destructivum, C. linicola, and C. truncatum [
33]. The combination of molecular diagnostic tools, along with morphological techniques, is therefore the best approach for studying Colletotrichum species complexes [
34].Doyle and Gaut [
35] described that single genes are usually insufficient to resolve interspecific relationships in Colletotrichum, which is why phylogram trees derived from single genes always yield limited information at lower taxonomic ranks. Using single-gene ITS cannot clearly distinguish between C. lentis and C. truncatum [
29]. Previous studies have indicated that using a combined dataset analysis of datasets to generate multigene phylogenetic trees could provide a higher overall support than any of the single-locus phylogenies [
36,
37]. In this study, phylogenetic analyses based on the combination of ITS, ACT, HIS3, and GAPDH sequences clearly distinguished C. truncatum (YN1932501, YN1932502, YN1932503, and YN1932504) from the other closely related Colletotrichum species.C. truncatum has a broad host range and can reportedly infect numerous plant species, such as Chinese flowering cabbage, soybean, lentil, common vetch, alfalfa, chili, solanaceous crops, and hemp plants [
17,
29,
38,
39,
40]. To the best of our knowledge, no previous record exists of C. truncatum causing disease in sword bean. This is, therefore, the first report of sword bean anthracnose caused by C. truncatum.


留言 (0)