Mammalian mitochondria can contain up to 16 potential H2O2 sources, 12 of which are associated with nutrient catabolism and the electron transport chain (ETC) [1,2]. The 12 sites are subdivided into two categories based on the redox pair that drives H2O2 production [3]. The NAD+/NADH isopotential group is comprised of α-keto acid dehydrogenases (α-ketoglutarate dehydrogenase; KGDH, pyruvate dehydrogenase; PDH, branched chain keto acid dehydrogenase; BCKDH, 2-oxoadipate dehydrogenase; OADH), and complex I when oxidizing NADH [2,4]. The second is the ubiquinone (UQ)/ubiquinol (UQH2) isopotential group, enzymes that rely on the oxidation or reduction of the UQ pool for H2O2 production [2]. This second group contains glycerol-3-phosphate dehydrogenase (G3PDH), proline dehydrogenase (PRODH), electron-transfer flavoprotein:ubiquinone oxidoreductase (ETFQO), complexes II and III, and DHODH, respectively. The reactive oxygen species (ROS) release profiles for each individual enzyme in these groups is complex and dependent on substrate availability and concentration, O2 tension, mitochondrial membrane polarity, supercomplex assemblies, post translational modifications, and many other factors [5]. For example, G3PDH and PRODH can generate modest amounts of H2O2 in liver and cardiac mitochondria [6]. However, most of the H2O2 formed during glycerol-3-phosphate and proline oxidation occurs through reverse electron transfer (RET) and forward electron transfer (FET) to the respiratory complexes [7]. Additionally, some of these sites, like complexes I and III and potentially KGDH and PDH, serve as cell redox signaling platforms.
DHODH is integral for cell proliferation because it catalyzes the oxidation of dihydroorotate to orotate for pyrimidine biosynthesis [8]. This has made it a potential site for cancer treatment and inhibition of tumour progression. Isoforms for DHODH can be subdivided into two categories based on subcellular location and sequence alignment between several different organisms. In mammalian cells, the class 2 isozyme is inserted in the mitochondrial inner membrane with the dihydroorotate binding site facing the intermembrane space [9]. The hydrophobic alpha-helices that insert DHODH into the membrane contain a flavin mononucleotide (FMN) group that funnels electrons from dihydroororate to the UQ binding site [9]. The ubiquinol is then oxidized by complex III of the ETC. Thus, DHODH-mediated orotate generation also drives oxidative phosphorylation. Due to its redox properties, DHODH can also produce ROS. DHODH is a significant H2O2 source in cancer cells and can produce modest amounts of ROS in rat skeletal muscle mitochondria [10]. However, early studies reported that the source of ROS may be outside of DHODH and most of the H2O2 is generated by RET and FET to the complexes [11,12]. This was confirmed in a study published by Hey-Mogensen et al. where it was concluded using a panel of inhibitors that DHODH displays a small rate of ROS production (20–30 pmol H2O2·min−1·mg protein−1 in rat skeletal muscle mitochondria) [10]. By contrast, there is a 10-fold increase in production during RET to the ETC during dihydroorotate oxidation, demonstrating the major source of O2●-/H2O2 is the respiratory chain [10].
Protein S-glutathionylation is a ubiquitous and redox sensitive covalent modification involving the addition and removal of reduced glutathione (GSH) to and from protein cysteine thiols [13]. Glutathionylation occurs in response to the conversion of GSH to GSSG by the glutathione peroxidase-mediated metabolism of H2O2 or elimination of lipid hydroperoxides [14]. Deglutathionylation is driven by restoration of the reductive power of the GSH by NADPH [14]. In mitochondria, protein S-glutathionylation is mostly associated with the inhibition of enzyme function in response to an increase in H2O2 availability, which has been documented to protect from oxidative distress and irreversible protein deactivation [15,16]. The modification is temporary and mitochondrial functions can be restored by an increase in glutathione reductase-dependent GSH availability. It was also shown that glutathionylation prevents further ROS production by complex I, KGDH, and PDH in cardiac, skeletal muscle, and liver tissue from rodents, an effect that could also prevent oxidative distress and may even play a role in regulating the use of H2O2 as a “mitokine” (reviewed in Ref. [17]). Our group is invested in interrogating the impact of glutathionylation on the 12 identified mitochondrial ROS generators. So far, we have shown glutathionylation inhibits ROS production by KGDH, PDH, complex I, and potentially complex III. Here, we show DHODH is a target for glutathionylation, a modification that also decreases mitochondrial ROS production during dihydroorotate oxidation. These findings are highly significant since we identified: 1) the first target for the glutathionylatio-mediated inhibition of H2O2 production in the intermembrane space, 2) a sex dimorphism in this pathway, an observation that builds on our previous discovery that there is sex difference in the regulation of cell ROS availability by glutathionylation.

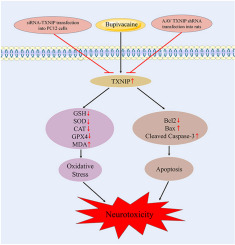
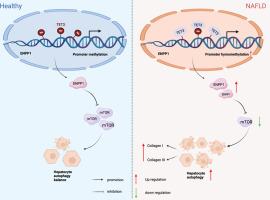
留言 (0)