

記住我
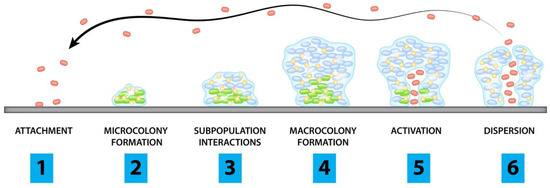
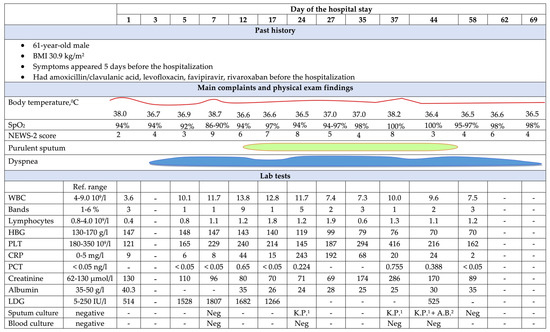

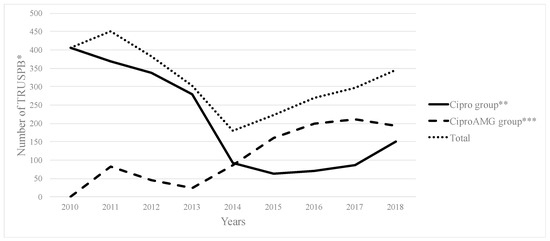
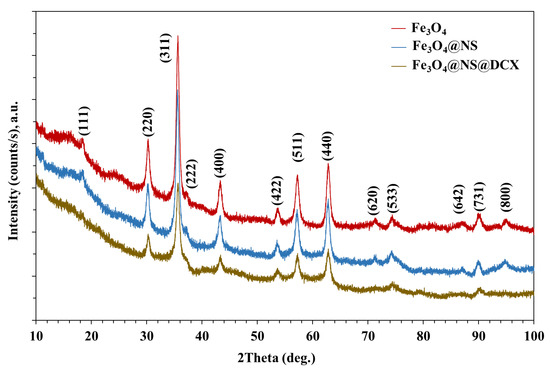

The clinical strain belonged to ST540 as the alleles adk_6, fumC_7, gyrB_57, icd_1, mdh_8, purA_8, and recA_2 exhibited 100% identity to their respective locus, as per MLST typing. Based on Clermon typing, the strain belonged to phylogroup A, a group largely dominated with commensal origins of strains. The E. coli ECU32 belongs to serotype H30-H with 100% identity for fliC in scaffold8|size160413, fimH54 (scaffold11|size140663), and O9-O antigen (>99% identity to wzm and wzt in scaffold34|size42957). These gene clusters were found between hisI and gnd genes, and belong to group 1 K antigens.
2.2. Antimicrobial-Resistant GenesUpon analyzing the sequence of E. coli strain ECU32 for genetic clusters, a 9852-base-pair genomic island was identified that carried the CRISPR-associated proteins, and the 5540-base-pair genomic island (scaffold94xsize3049) showed the presence of ESBL, the class-A blaTEM beta-lactamase (A8A11_02620).
The 32,081-base-pair genomic island in the strain (scaffold7xsize174057) highlighted the presence of members from the EscJ/YscJ/HrcJ family type III secretion system (from A8A11_10065 to A8A11_10125).
The largest genomic island present in strain ECU32, 131,243bp in size (consisting of scaffold78xsize4056 and scaffold83xsize2691), indicated the presence of aminoglycoside-modifying enzymes APH(3′) (A8A11_04060), APH(6)-Id (A8A11_04065),and class 1 integrase, followed by gene cassette dfrA1 (A8A11_21925). The class-C ampC detected in this study exhibited 97% identity to a homolog found in E. coli O157:H7; 95% to Shigella sonnei; 78% to Enterobacteriaceae bacterium; 73% to Shigella flexneri; 71% to Enterobacter, and 70% to Yersinia ruckeri.
The quinolone resistance gene qnrS1 was found in scaffold57xsize16952 (A8A11_12295) with an adjacent ISKra4-like element (A8A11_12305), while the sulfonamide-resistant dihydropteroate synthase sul2 was found in scaffold78xsize4056 (A8A11_04055). The quinolone resistance determinant exhibited identities to homologs found in Vibrio mytili CAIM 528 (95%); Photobacterium ganghwense strain DSM 22954 (92%); Vibrionales bacterium SWAT-3 (83%); and Photobacterium halotolerans strain MELD1 (66%). The sequence analysis of QnrS1 indicated the conserved pentapeptide sequences, and homology modeling highlighted the differences in loops A and B regions, as shown in Figure S1.Homologs of well characterized efflux pumps, AcrAB, EmrAB, MdtABC, EmrE, AcrD, and TetA (scaffold82xsize4187; A8A11_05610), were present in the genome; moreover, the marR gene harbored sense mutations at the Y137H and G103S residues. The tetracycline efflux pump exhibited varied identities to homologs found in Aeromonas simiae CIP 107798 (98%), Pseudomonas fluorescens HK44 (97%), E. coli O83:H1 str. NRG 857C (97%), and Clostridium nexile DSM 1787 (78%).
Using the BacWGSTdb server, the resistance genes were assessed amongst ST540 strains from different countries, hosts, and isolation sources, and the ECU32 strain exhibited the presence of diverse resistance genes (Figure 2A). 2.3. Virulence and Its Associated GenesComparative genome sequence analysis revealed that E. coli strain ECU32, in scaffold21xsize74584, carried ORFs for both the ferric enterobactin ABC transporter system (fepA, fepB, fepC, fepD, fepG, fes), and enterobactin (siderophore) biosynthesis exporter system (entB, entC, entD, entE, entF, entS). Important virulent factors that were identified included the type 1 fimbriae adhesin fimH (scaffold11xsize140663-1, A8A11_14300), hemolysin E hlyE (scaffold22xsize79984, A8A11_04330), invasin of brain endothelial cells, ibeB (scaffold60xsize15487, A8A11_08140), gspC, gspD, gspE, (scaffold35xsize41361), and intimin-like adhesion fdeC (scaffold1xsize239491 with 93% to EC958_0448).
Virulence genes were analyzed and compared with different ST540 strains and pathotypes. The analysis demonstrated the prevalence of virulence genes that are involved in adherence, autotransporter, invasion, non-LEE-encoded TTSS effectors, secretion system, toxin, and others (Figure 2B,C). 2.4. Phenotypic Characterization of Multidrug-Resistant E. coliThe Kirby Bauer assay revealed the E. coli strain ECU32 to be multidrug resistant (AMP, AMK, CLI, CST, ERY, KAN, LZD, MET, OXA, PEN, RIF, STR). The minimum inhibitory concentrations of this Indian isolate for different antibiotics, as evaluated by the agar dilution method, were ceftazidime 0.5 µg/mL, ticarcillin >1024 µg/mL, neomycin 4 µg/mL, norfloxacin 16 µg/mL, doxycycline 64 µg/mL, tetracycline 64 µg/mL, nalidixic acid >1024 µg/mL, ampicillin >1024 µg/mL, kanamycin 16 µg/mL, erythromycin >1024 µg/mL, trimethoprim 64 µg/mL, and chloramphenicol 4 µg/mL.
E. coli, a common flora in the human gastrointestinal tract, has adaptive survival response strategies under different stress conditions. The growth profile of E. coli ECU32 was assessed in the presence of low to high pH concentrations. The strain retained the ability to grow in pH 5.0 to pH 8.0; meanwhile, in pH 10, the strain displayed less (~0.5-fold) growth compared to pH 5.0 (Figure 3A). Concentration-dependent growth of the strain in the presence of the antibiotic norfloxacin was observed. The strain was able to grow in doses upto 64 µg/mL, and exhibited nine-fold lower growth compared to its control (Figure 3B).However, the E. coli ECU32 strain was able to survive in the presence of both oxidative and nitrostative stress-inducing agents, such as hydrogen peroxide, sodium nitroprusside, and sodium nitrite, respectively (Figure 3C–E). The survival of E. coli ECU32 under osmotic stress conditions was determined in the presence of different sodium chloride concentrations. The strain exhibited more than 80% survival upto 0.75 M, and 40% in 1 M of sodium chloride (Figure 3F). Altogether, these observations emphasize the adaptability of E. coli under any intracellular stress conditions, and its survival inside the host to cause disease severity.The E. coli ECU32 strain has an ability to cope with different intracellular stress responses. Furthermore, the ability of the strains to survive in the presence of a various range of antimicrobial agents was examined. Analyses revealed a more than 50% survival rate was observed for the strain at 1024 µg/mL for ampicillin, 4 µg/mL for neomycin, and 0.5 µg/mL for tetracycline. Furthermore, the strain was found to have retained the ability to survive in various antimicrobial compounds, such as acriflavine, acridine orange at 256 µg/mL, and saffranine and deoxycholate at 1024 µg/mL (Figure 3G).In order to detect efflux activity in the E. coli strain, MICs for the following efflux-based substrates were initially analyzed: acridine orange 256 µg/mL, acriflavine 64 µg/mL rhodamine >1024 µg/mL, saffranine >1024 µg/mL, deoxycholate >1024 µg/mL, and SDS >1024 µg/mL. Hence, a growth inhibition assay was performed with the E. coli strain ECU32, using ampicillin 256 µg/mL, in the presence of known efflux pump inhibitor PAβN, which exhibited an approximatelyfive-fold reduced growth relative to its control. Overall, the assays demonstrated that the strain has the ability to survive under different assails, including antimicrobial compounds. Furthermore, the sequence analysis and phenotypic efflux assays indicated the role of active efflux as the primary mechanism used to confer antimicrobial resistance. In order to substantiate these observations, genomic analysis of the strain further confirmed the presence of well-characterized efflux families, depicting the importance and possible involvement of these pumps in antimicrobial resistance and multiple cellular functions.
The strain was examined for its resistance level towards hospital-based disinfectants, and it exhibited tolerance as follows: benzalkonium chloride 12.8 µg/mL, chlorhexidine 0.1 µg/mL (Figure 3H(i,ii)), indicating that the urinary strain was broad-spectrum antimicrobial, as well as biocide-resistant. Additionally, the E. coli strain ECU32 displayed an ability to form biofilms (ratio 570/600 nm = 0.252), as it harbored genes that are required for adhesion and virulence.Upon transforming the plasmid preparation from the MDR strain into E. coli JM109, transformants were confirmed on ampicillin (>256 µg/mL) and ciprofloxacin (0.5 µg/mL) plates; subsequent PCR detection indicated the presence of quinolone resistance determinant in the transformant, and further sequencing confirmed the role of qnrS-dependent plasmid-mediated resistance in E. coli strain ECU32.
It is worthwhile to state here that ST540 has usually been reported in E. coli that was isolated from birds and chickens. Identifying the serotype ST540 from a biological sample strictly emphasizes the periodic monitoring of emerging E. coli strains at the One-Health Interface. The E. coli strains TOP2386 (accession no: AORB01), TOP2396_1 (accession no: AOQQ01), TOP2515 (accession no: AOQT01), TOP2522_1 (accession no: AOQU01), and TUM3433 (accession no: BGLY01), isolated from human samples in USA and Japan, belong to the same ST, as that of E. coli strain ECU32. As per the BacWGSTdb server, E. coli strain ECU32 (accession no: LZGD01) with ST540 is being reported for the first time in India.
Figure 3. Biological characterization of the E. coli ECU32 strain. The growth profile of the E. coli ECU32 strain was monitored in the presence of different pHs (A) and concentrations of norfloxacin (B). The growth of the ECU32 strain was monitored in the presence of varied concentrations of oxidative stress-inducing agent—hydrogen peroxide (C) and nitrostative stress-inducing agents—sodium nitrite and sodium nitroprusside (D,E). The osmotic stress response survival was determined in the presence of sodium chloride concentrations (F). The survival ability of the E. coli ECU32 strain was determined under different concentrations of antibiotics [ampillicin, neomycin, tetracycline], bile [deoxycholate], structurally unrelated compounds and dyes [acriflavine, acridine orange, saffranine] (G), and biocides [benzalkonium chloride, chlorhexidine, and triclosan] (H(i,ii)). The mean values of the different independent experiments were used for plotting graphs, using GraphPad Prism.
Figure 3. Biological characterization of the E. coli ECU32 strain. The growth profile of the E. coli ECU32 strain was monitored in the presence of different pHs (A) and concentrations of norfloxacin (B). The growth of the ECU32 strain was monitored in the presence of varied concentrations of oxidative stress-inducing agent—hydrogen peroxide (C) and nitrostative stress-inducing agents—sodium nitrite and sodium nitroprusside (D,E). The osmotic stress response survival was determined in the presence of sodium chloride concentrations (F). The survival ability of the E. coli ECU32 strain was determined under different concentrations of antibiotics [ampillicin, neomycin, tetracycline], bile [deoxycholate], structurally unrelated compounds and dyes [acriflavine, acridine orange, saffranine] (G), and biocides [benzalkonium chloride, chlorhexidine, and triclosan] (H(i,ii)). The mean values of the different independent experiments were used for plotting graphs, using GraphPad Prism.
留言 (0)