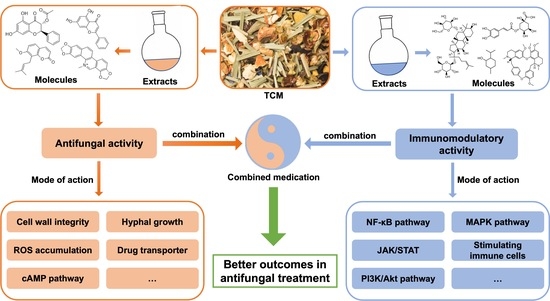
1. IntroductionMicrosporum canis is an important zoophilic dermatophyte in domestic cats and dogs; however, it also causes dermatophytosis in humans. Human infections are acquired from domestic animals through direct contact with clinically or subclinically infected animals [
1]. M. canis is a major species of keratinophilic and keratinolytic filamentous fungi that cause superficial fungal infections worldwide [
2], particularly in Europe, the eastern Mediterranean, and South America [
3,
4,
5]. M. canis infections in cats, dogs, and other domestic animals such as rabbits generally manifest as multifocal alopecia, scaling, and circular lesions [
6,
7]. Both stray and domesticated cats are considered common reservoirs for M. canis [
8,
9,
10] and anyone in direct contact with these animals, including owners and veterinarians, is at risk of becoming infected [
9]. Treatments include oral and topical preparations of antifungal agents such as amphotericin B, griseofulvin, terbinafine, itraconazole, flucytosine, and fluconazole, administered alone or in combination. These agents cause adverse reactions, such as nephrotoxicity, hepatotoxicity, and neurotoxicity and have been associated with treatment failure secondary to drug resistance [
11,
12]. One antifungal-resistant strain of M. canis is resistant to terbinafine and has been reported in some feline patients [
13]. Factors mitigating drug resistance include a lack of patient compliance, poor tissue penetration, and variable medication bioavailability [
11,
14]. Biofilms produced by M. canis are an additional concern [
15,
16]. Biofilm refers to the matrix that surrounds the microbial population and acts as a physical barrier, protecting the microorganisms in it. Biofilms are typically associated with increased resistance to antifungal agents [
15].Plants, including their extracts, contain phytochemical compounds that are used as antimicrobials and antibiofilm agents [
17]. Identifying novel antifungal targets and compounds derived from natural plants will help develop novel antifungal strategies and improve existing ones [
15]. Several previous (and ongoing) studies have focused on the use of plant extracts as alternative treatments for fungal infections [
12]. Plant extracts that have antifungal activity include lemon grass (Cymbopogon citrates), lantana (Lantana camara), nerium (Nerium oleander), basil (Ocimum basilicum), and olive leaves (Olea europaea) [
18]. The ethanolic extracts of Echinophora platyloba inhibit Candida albicans ATCC 10231 [
19]. The aqueous and ethanolic extracts and the essential oil of Thymus capitatus exhibit antifungal activity against C. albicans and M. canis [
20]. Extracts of Ocimum gratissimum leaves display antifungal activity against M. canis, Microsporum gypseum, Trichophyton rubrum, and Trichophyton mentagrophytes [
21]. Methanolic leaf extracts of Eucalyptus camaldulensis were investigated for suspected in vitro antifungal activities against M. canis, M. gypseum, T. rubrum, Trichophyton schoenleinii, T. mentagrophytes, and Epedermophyton floccosum [
22].Coleus amboinicus Lour. (synonym: Plectranthus amboinicus [Lour.] Spreng), whose common name is Indian borage, is widely cultivated in tropical Africa, Asia, and Australia. This perennial herb belonging to the family Lamiaceae contains several phytochemicals, including monoterpenoids, diterpenoids, triterpenoids, sesquiterpenoids, phenolics, flavonoids, and esters [
23,
24]. The geographic location of C. amboinicus cultivation influences the phytochemical composition of plant extracts [
25]. The stem, leaf, and root extracts of C. amboinicus contain high concentrations of polyphenolics such as caffeic acid, rosmarinic acid, apigenin, chrysoeriol, 5-O-methyl-luteolin, and 5,8-dihydroxy-7,2′,3′,5′-tetramethoxyflavone, which is a novel flavonoid [
26,
27,
28]. Several studies have reported the wide range of pharmacological properties of C. amboinicus extracts and essential oil, including antioxidant activity [
25], antidandruff action [
29], antiproliferative effects on cancer cells [
30], analgesic and anti-inflammatory activities [
31], antirheumatoid arthritis [
32], anti-inflammatory effect following bone injury [
33], antibacterial activity against methicillin-resistant Staphylococcus aureus [
34], mosquitocidal and water sedimentation properties [
35], and insect repellent [
36] and insecticidal effects [
37].Although C. amboinicus exhibits antifungal activity against several fungal species [
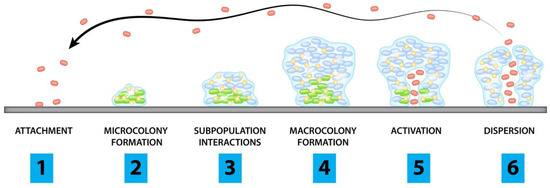
38], little is known of its antifungal effects on dermatophytes and the biofilms they produce. Therefore, we evaluated the antifungal effects of C. amboinicus essential oil (CEO) and C. amboinicus ethanolic extracts (CEE) against planktonic cells and biofilm formation of M. canis clinically isolated from a feline dermatophyte. 3. DiscussionTo the best of our knowledge, this is the first report of the antifungal activity of C. amboinicus extracts against planktonic cells and the biofilm formation of M. canis. M. canis biofilms are composed of a multidirectionally expanded network of hyphae linked together by a polysaccharide extracellular matrix [
16]. Biofilm reduces the penetrability of antifungal agents, thus contributing to treatment failure and recurrent infection [
39,
40]. The inhibitory effect of antifungal agents on biofilm formation was observed at concentrations higher than those required to inhibit the growth of planktonic cells [
41]. Fungal biofilm formation is a key factor in fungal virulence, persistence, and invasion as well as recurrent fungal infections and conventional antifungal resistance [
40,
42]. The time-dependent adherence of arthroconidia was observed, starting at 2 h and up to 6 h after inoculation. M. canis produced keratinolytic enzymes and secreted endo and exoproteases during adhesion; this process was likely inhibited by chymostatin, a serine protease inhibitor [
43]. After biofilm formation for 72 h, a polysaccharide extracellular matrix that links fungal hyphae was observed [
16,
44]. The extracellular matrices of poor, moderate, and strong biofilm-producing M. canis appear to be related to mechanisms of antifungal resistance; however, further investigations are needed to confirm this. Flucytosine or fluconazole treatment at every 6–24 h could not completely destroy the biofilms of Candida spp. Poor drug penetration might not be a major mechanism of antifungal resistance for Candida biofilms [
45]. During the early stage of C. albicans biofilm formation, genes encoding efflux pumps are upregulated, thereby mediating antifungal resistance [
46]. Developing new compounds or alternative inhibitors to treat biofilm-related drug resistant fungal infections is essential to veterinary and human medicine [
40,
42,
47]. In this study, fluconazole (4–16 µg/mL) had no effect on the mature biofilms of M. canis isolates. This result was similar to those reported by Bila et al., who found that fluconazole only inhibited the metabolic activity of early-stage biofilms of T. mentagrophytes at 32 mg/L but did not exhibit antibiofilm activity on mature biofilms, even at the highest concentration (512 mg/mL) [
48].Our study also demonstrated that C. amboinicus can inhibit planktonic cell growth and biofilm formation of feline zoonotic M. canis. CEO and CEE significantly inhibited the planktonic cell growth of M. canis at 128 µg/mL (32–256 µg/mL). Considering the MIC90 values, CEO was found to have a higher potency more potent than CEE against all M. canis isolates (MIC90 of 128 µg/mL vs. 256 µg/mL). C. amboinicus has been reported to exhibit antifungal activity against several fungi, including Aspergillus clavatus, Aspergillus niger, Cladosporium cladosporioides, Chaetomium globosum, Myrothecium verrucaria, Penicillium citrinum, Trichoderma viride, and Mucor sp. [
38,
49]. It also inhibits the biofilm formation of other pathogenic microorganisms, such as Streptococcus mutans, Streptococcus pyogenes, and S. aureus [
50,
51,
52]. C. amboinicus is rich in monoterpenes, including carvacrol, thymol, eugenol, chavicol, and ethyl salicylate [
37,
49,
53]. The 18 compounds identified in CEO in this study represent 99.84% of the total essential oil and include carvacrol (56.65%), p-cymene (10.89%), and γ-terpinene (9.33%); these three compounds alone comprise 76.87 % of the total essential oil. The concentrations of carvacrol, β-caryophyllene, and thymol were found to be 3.4 ± 0.2, 0.35 ± 0.16, and 0.013 ± 0.08 mg/mL, respectively. This result differs from that reported by da Costa et al., who found thymol to be the major constituent (64.3%), followed by p-cymene (10.3%), γ-terpinene (9.9%), and β-caryophyllene (2.8%) [
54]. Previous studies have reported that the phytochemical composition of CEO is significantly influenced by the cultivation location, processes, and methods of essential oil extraction [
25,
37]; for example, steam distillation produced higher levels of carvacrol in C. amboinicus essential oil than those produced via the hydrodistillation method [
37]. CEO at MIC had excellent effects against all clinical isolates. The high potency of CEO may be attributed to the hydrophobic property of essential oil, which adversely affects every step of biofilm formation, including adhesion, growth, maturation, and dissemination. The antibiofilm mechanisms of essential oil include reducing bacterial adhesions, preventing fresh biofilm formation, and destroying existing biofilm [
55,
56].Carvacrol appears to be a major contributor to the antifungal properties of CEO. Carvacrol, p-cymene, and γ-terpinene are monoterpenes that exhibit various biological activities, including antioxidative, anti-inflammatory, anxiolytic, antineoplastic, and antimicrobial effects [
57]. The antimicrobial effects of carvacrol are effective against various microorganisms, including bacteria such as S. aureus and Pseudomonas aeruginosa and fungi such as C. albicans, Candida glabrata, and Candida parapsilosis [
58]. In a previous study, Lavandula multifida L. essential oil containing carvacrol as the main constituent was effective against dermatophytes (MIC: 0.16 μL/mL) and Cryptococcus neoformans (MIC: 0.32 μL/mL) [
59]. Carvacrol also exhibits antifungal activity against Aspergillus spp. (MIC: 100 µg/mL) and Cladosporium spp. (MIC: 100 µg/mL) [
60,
61], possibly by binding to sterols in the fungi. The sterols residing on planktonic cell membranes are essential for their survival [
62], and their hydrophilic properties allow carvacrol to penetrate the polysaccharide layer of their biofilm matrix, thereby destabilizing the biofilm [
63]. A recent study reported that p-cymene exhibited no antifungal activity against A. niger (MIC: >300 µL/mL) and Rhizopus oryzae (MIC: >1024 µg/mL) [
64], whereas γ-terpinene has been shown to exhibit antifungal activity against Sporothrix schenckii (MIC = 62.5–500 µg/mL) and Sporothrix brasiliensis (125–500 µg/mL) [
65].In the present study, CEE effectively eradicated the biofilm formation of weak, moderate, and strong biofilm producers at 2× MIC. Total phenolic and flavonoid contents were positively correlated with the antimicrobial activity of the plant extracts [
66]. We found higher total phenolic and flavonoid levels in CEE than those reported in previous studies. For example, the ethanolic extract of C. amboinicus leaves obtained from Vietnam had a total phenolic and total flavonoid content of 26.84 ± 0.91 µg GAE/mg sample and 12.14 ± 0.42 µg QE/mg sample, respectively [
67]. A methanolic extract of the C. amboinicus stem obtained from India had a total phenolic content of 49.91 mg GAE/g sample and total flavonoid content of 26.6 mg rutin equivalent/g sample [
68]. Flavonoids inhibit nucleic acid biosynthesis and spore germination in plant pathogens [
69,
70]. High phenolic and flavonoid levels may thus be related to the significant antifungal effects of CEE. Importantly, CEE contained remarkable levels of rosmarinic acid (1.251 mg/g sample), apigenin (1.175 mg/g sample), and caffeic acid (0.732 mg/g sample) in this study. Rosmarinic and caffeic acid compounds have significant antifungal effects against Fusarium oxysporum [
71]. The antifungal mechanism of rosmarinic acid is poorly understood but is believed to be related to the RTPase enzyme [
72]. Apigenin at a concentration of 5 µg/mL exhibited antifungal activity against C. albicans, C. parapsilosis, Malassezia furfur, T. rubrum, and T. beigelii by inhibiting biofilm formation and efflux-mediated pumps of fungi. It also induced cell death by interfering with membrane function and increasing cell permeability [
73,
74]. Mice infected with T. mentagrophytes recovered after treatment with apigenin ointment administered at concentrations of 2.5 and 5 mg/g on the 12th and 16th days, respectively [
75]. Caffeic acid phenethyl ester, a major active component of propolis (Apis trigona), has been shown to exert concentration-dependent effects on planktonic cells and biofilm formation of different Candida species [
76] and synergistically enhance the antifungal activity of fluconazole against resistant clinical isolates of C. albicans [
77]. Another study reported that the fungicidal activity of caffeic acid against T. rubrum was observed at 86.59 μM; this activity was mediated via plasma membrane damage and reduced ergosterol production, where caffeic acid reduced isocitrate lyase activity and downregulated critical genes (ERG1, ERG6, and ERG11) required for ergosterol synthesis [
78].
Although CEO and CEE had different chemical constituents, both exhibited excellent and comparable inhibitory activities against all fungal isolates obtained from feline dermatophyte samples. Our findings suggest that both CEO and CEE act as natural antifungal agents against planktonic cells and biofilm-producing M. canis. Future investigations of the relationship between plant-based compounds, such as carvacrol in CEO and apigenin in CEE, their mechanisms of action, and classification based on biofilm production may contribute to a better understanding and guide the development of safe and effective antifungal agents derived from natural sources.



留言 (0)