
記住我


Our results have demonstrated that inhibitory and bactericidal concentrations of gold(III) complexes tended to remain constant within the different isolates of the same species. Concerning the origin, the activity of gold(III) complexes seems to be similar among human clinical or veterinary isolates. Thus, except in the case of 2b, 2d and 3, which were more active against human clinical strains, the MICs observed in this study for the veterinary strains are in concordance with previous data obtained by the research group against human clinical isolates from the same species (unpublished data).
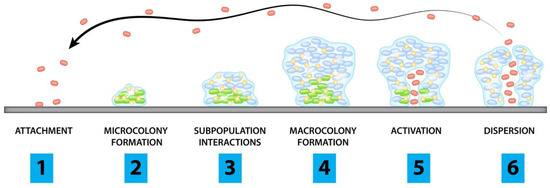
The tested concentrations of the gold(III) complexes have reflected a bacteriostatic effect on bacterial growth of both Gram-negative and Gram-positive bacteria, although we have also observed that the increase of one or two folds in these concentrations leads to a bactericidal effect, except for 2f which presents a greater increase for any of the species. As mentioned above, the overuse of antibiotics has led to the emergence of bacteria that can withstand inhibitory concentrations and can prompt bacteria to persist [23], which is why the emphasis should be focus on development of antibiotics with bactericidal effect.The impact of biofilms on human infections has been studied more in depth than in animal health, having a high importance in veterinary medicine, leading to a negative repercussion in economy and animal production. This biofilm state of life brings to bacteria a high resistance to antibiotics and immune system. The biofilm structure confers resistance via multiple mechanisms: delayed diffusion through the polysaccharide matrix; expression or repression of different genes that confer a range of physiological responses; and growth rate reduction and presence of persister cells [29].Currently, the trend on biofilm treatment in animals is to use combinations of antibacterial and antibiofilm agents, electric currents, nanocarriers and ultrasound [30]. Thus, disruption of the biofilm would facilitate the efficacy of antimicrobial agents and minimize the scope for the development of antibiotic resistance [31]. The investigated gold(III) complexes affected biofilm formation among all Gram-negative strains studied, obtaining the same value or ±1-fold compared to their planktonic activity. However, against Staphylococcus spp. no biofilm inhibition at the tested concentrations was observed. According to our results, antibiofilm activity of several gold(III) complexes has been shown against P. aeruginosa in either pre-adhesion and post-adhesion phases of the biofilm formation [27]. Nevertheless, there are studies of metal-based drugs such as gold-complexed sulfonamide that effectively promoted a remarkable reduction in the bacterial adhesion against S. aureus strains [32].CST is one of the antibiotics that has been used in veterinary medicine for decades to prevent and treat infections caused by Enterobacteriaceae species. This antibiotic has been widely used in animal feed with the consequence that it can reach humans through the food chain [33]. In humans, it is considered a last resort antibiotic due to its neurotoxicity and nephrotoxicity [34]. However, colistin has been recovered to treat those MDR bacterial strains that conventional antibiotics are not able to kill. When monotherapy treatment of both CST and 2b was compared with the combined action of 2b gold(III) complexes with CST, the activities of the single agents were enhanced and a synergistic effect was achieved in the trials. The mechanism of action of CST is based on binding to lipopolysaccharides and phospholipids of the outer cell membrane of Gram-negative bacteria [35]. E. coli GN1044 were resistant to CST, but the combined action of these antimicrobials was able to inhibit bacterial growth by reducing the concentrations of each agent at least three-fold. In fact, the combined action with the gold(III) complex 2b was able to lower the MIC of CST below the breakpoint of ≤2 mg/L, according to EUCAST guidelines [36], same values as those found in bacteria susceptible to this antibiotic. Similar results were found using 1 against CST-resistant clinical strains, decreasing four-fold the MIC of CST and 1 [21]. Studies have shown the synergistic effect of CST in combination with other clinically relevant antibiotics such as ceftazidime against MDR bacteria [37]. Researchers have proposed that CST may exert a subinhibitory permeabilizing effect on the outer membrane of Gram-negative bacteria even in CST-resistant isolates [38]. It appears that the outer membrane compromises the activity of gold(III) complexes, so that when CST permeabilizes the bacterial outer membrane, the activity of gold(III) complexes is greatly enhanced. Indeed, this provides a promising therapeutic approach for animals infected with CST-resistant Gram-negative pathogens. A plausible explanation for the observed synergy between the gold complexes and colistin would be in the initial binding of the latter to anionic lipid A molecules of Gram-negative bacteria [5,6], leading to a rapid permeabilization of the outer cell membrane that allows improved penetration of the gold(III) complexes [39,40]. In the absence of colistin, the permeability barrier conferred by the outer membrane of Gram-negative bacteria impairs the transfer of gold complexes to the cytoplasm [17], so that when CST permeabilizes the bacterial outer membrane, the activity of gold(III) complexes is greatly enhanced. Indeed, this provides a promising therapeutic approach for animals infected with CST-resistant Gram-negative pathogens.Even though the data set is very limited, it is possible to gather some useful information regarding the SAR. In general terms, Au(III) (C^S)-cyclometallated complexes 2 and 3 with a bidentate (S^S) ancillary ligand are more active against both Gram-positive and Gram-negative strains than complex 1, containing two monodentate chloro ancillary ligands. Regarding the structure of the cyclometallated core, there are not salient differences in the activity of dppta complexes 2 when compared to bppta complex 3.
Complexes 2 and 3 are all very active against Gram-positive strains and the best results were obtained for complex dimethyldithiocarbamate 2a. In this sense, complex 2a is the less sterically hindered of the whole series, which could contribute to its higher effectiveness, because the lack of bulky substituents facilitates intermolecular interactions between the complexes and molecular components of target cells. Along this hypothesized steric effect is the excellent activity of complex 3 against Gram-positive strains. However, the very good activity of morpholinyl dithiocarbamate 2d is rather surprising considering steric effects alone.
Steric factors also cannot explain the pattern observed for the activity of DTC complexes 2 and 3 against Gram-negative strains. In this case, complexes 2b, 2c and 2f containing cyclic aliphatic ligands display better antibacterial activity than complexes 2a and 3 with smaller ligands. Moreover, the best activity in the series is observed for complex 2f with the largest azepanyl dithiocarbamate ligand. These results point to a different mechanism of antibacterial action for Gram-negative strains; a plausible hypothesis would be the intercalation of multiple molecules of the gold complex within the bilipid layer causing membrane damage, an effect which was previously described for the action of gold complexes on Gram-negative bacteria [41]. This intercalation would be more effective with bigger complexes, explaining the higher activity of the most sterically hindered complex.Regarding the antibiofilm activity, biofilm is a complex matrix of extracellular polymeric substances, and unlike planktonic cells, it has a strongly hydrophobic character [42]. Therefore, biofilm would have more affinity to apolar substances, which would explain the greater effectiveness in the treatment of the biofilm of more apolar complexes 2b, 2c and 2f.Furthermore, gold(III)-complex 1 showed lower in vitro toxicity in the Jurkat E6.1 cell line compared to the DTC complexes 2a–f and 3 panel of molecules tested, which is consistent with the results of previous studies [22]. Specifically, this gold(III) complex showed lower antibacterial activity against Gram-negative cells than 2a, 2b and 2e which had been classified as potential antimicrobial and antibiofilm agents. The slightly increased toxicity over Jurkat E6.1 cells (a tumoral cell line) could be related to the antitumoral activity commonly associated with gold(III) complexes [43]. In contrast to Gram-positive bacteria, the three gold(III) complexes with the lowest cytotoxicity at therapeutic concentrations are classified as active molecules in this group. Some studies suggested that the cytotoxicity of gold(III) complexes is associated with the presence of the gold(III) center [44]. 4. Materials and Methods 4.1. ChemistryThe target (C^S)-cyclometallated Au(III) complexes were synthesized using established procedures with minor modifications. Thus, [Au(dppta)Cl2] complex 1 (dppta = ortho-N,N-diisopropyl-P,P-diphenylphosphinothioic amide) was prepared through tin(IV)-gold(III) transmetalation from the corresponding chlorodimethylstannyl derivative as previously reported [20].[Au(dppta)(dtc)][PF6] complexes 2a–f (dtc = dithiocarbamate, R2NCS2) were synthesized by the overnight reaction of [Au(dppta)Cl2] complex 1 with the corresponding dithiocarbamate sodium or potassium salt [45] in methanol, followed by the addition of aqueous potassium hexafluorophosphate (Method M1, Supplementary Materials).The products were characterized by 1H, 13C and 31P NMR, IR and HRMS. The transformation of the [Au(dppta)Cl2] complex 1 to the [Au(dppta)(dtc)][PF6] complexes was confirmed by the upfield shift in the characteristic signal of the H ortho to the Au (from δH H 8.47 to 7.75 ppm for 2a and 2b; 8.01 ppm for 2c; 7.84–7.91 ppm for 2d and 2e; 7.74–7.80 ppm for 2f) in the 1H NMR (Figures S1, S4, S7 and S10, Supplementary Materials). On the other hand, the 31P NMR spectra of complex 2 display a significant downfield shift from δP 68.2 (1) to 75.5–76.5 ppm (Figures S3, S6, S9 and S12, Supplementary Materials). The 13C NMR resonance for the NCS2 carbon appeared between δC 190.6 and 198.6 ppm (Figures S2, S5, S8 and S11, Supplementary Materials). Bands in the IR spectra between 1528 and 1579 cm−1 are attributed to the delocalized dithiocarbamate NCS2 system, while bands around 837–840 cm−1 are due to the PF6− anion. The complexation of the DTC ligand is further supported by the ion [M]+ observed in mass spectrometry for complex 2. The lack of fragmentation indicates appreciable stability, and is in agreement with other gold(III) complexes containing bidentate ligands [46].Similarly, addition of sodium diethyl dithiocarbamate to a methanolic solution of [Au(bppta)Cl2] complex 4 (bppta = ortho-bis(N,N-diethyl)-P-phenylphosphonothioic amide, [47] afforded, after addition of aqueous potassium hexafluorophosphate, [Au(bppta)(Et2NCS2)][PF6] complex 3 (Method M2, Supplementary Materials, δP 68.2, Figure S15).The transformation of the [Au(bppta)Cl2] complex 4 to the [Au(bppta)(Et2NCS2)][PF6] complex 3 was confirmed by the upfield shift of the signal of the H ortho to the Au [Au(bppta)Cl2] complex 4 from δH 8.24 to 7.48 ppm in the 1H NMR (Figure S13, Supplementary Materials). On the other hand, the 13C NMR spectrum displays the resonance for the NCS2 carbon at δC 195.1 ppm (Figure S14, Supplementary Materials). Further confirmation of the presence of the dithiocarbamate arises from the band observed at 1552 cm−1 in the IR spectra due to the C-N stretching absorption of the NCS2 system. The complexation of the dtc ligand is further supported by the ion of m/z 629.1323 observed in mass spectrometry corresponding to the cation [M]+ (calcd. for C19H33AuN2PS3 m/z 629.1313). The HRMS supports the formation of [Au(bppta)(Et2NCS2)][PF6] complex 3, showing a peak for ion [M]+. Again, the absence of fragmentation is indicative of a good stability.More detailed information about the experimental procedures and full spectroscopic for new complexes are included in the Supplementary Materials (Sections S1 and S3, respectively). Stock solutions were prepared in 100% dimethyl sulfoxide (DMSO, Sigma-Aldrich, Madrid, Spain) at 3000 or 6000 mg/L and stored at −20 °C until use. The final concentration used in the biological experiments was 0.5%, without exhibiting toxic effects. 4.2. Bacterial StrainsA panel of 16 strains of animal origin were used for the study: three Salmonella spp., six E. coli, five Staphylococcus spp. from different species, one S. uberis and one P. aeruginosa. These isolates came from the Institute of Agrifood Research and Technology (IRTA) of Barcelona (Spain). MALDI-TOF mass spectrometry was used to confirm identification and the strains were subsequently stored at −80 °C in skim milk (Becton Dickinson, Barcelona, Spain). E. coli ATCC 25922, P. aeruginosa ATCC 27853, and Staphylococcus aureus ATCC 29213 reference strains were included as controls.
4.3. Minimal Inhibitory Concentration DeterminationMICs were determined using the broth microdilution method, following the CSLI guidelines [48]. The tests were conducted in ISO-Sensitest Broth (Oxoid, Hamsphire, UK), according to recommendations of the International Standard ISO 20776-1. Standard antibiotics were also included in the assays in order to compare their activity and efficacy. In particular, ciprofloxacin, ampicillin, vancomycin, trimethoprim-sulfamethoxazole (Sigma Aldrich, Madrid, Spain), and colistin (MPBiomedicals, Eschwege, Germany) were investigated.Briefly, broth microdilution was performed in polystyrene 96-well round bottomed plates in a final volume of 100 μL. Serial two-fold dilutions of the antimicrobial ranging from 128 mg/L to 0.03 mg/L were used. In addition, wells with bacteria alone and with growth media alone were included as positive and negative controls, respectively. As for the inoculum, after an overnight incubation in agar plates, bacterial colonies were resuspended in 0.9% NaCl to reach a 0.5 McFarland (equivalent to 1.5 × 108 CFU/mL) and diluted to obtain a final concentration in the well of 5 × 105 CFU/mL. Finally, the plates were incubated for 18 h at 37 °C. MIC values were expressed in mg/L. We performed three technical and three biological replicates for each agent.
4.4. Minimal Bactericidal Concentration DeterminationThe MBCs are known as the lowest concentration of an antimicrobial agent that will prevent the growth of 99.9% of bacteria [49]. This is done by subculturing the broth dilutions used for MIC determination (i.e., wells above the MIC) on fresh agar plates [50]. Serial two-fold dilutions of the antimicrobial ranging from 128 mg/L to 0.03 mg/L were used. Thus, after microdilution assays, 10 μL aliquots of all wells showing no visible pellets or turbidity were seeded onto Luria-Bertani (LB) plates and incubated for 24 h at 37 °C. Results were expressed in mg/L and MBC values were defined as the last well at which no regrowth was observed. 4.5. Biofilm FormationBiofilm formation standardization was carried out only for microorganisms without previous experimental protocols in the laboratory, specifically for Salmonella spp. strains. The main aim was to evaluate the best experimental conditions and requirements for detecting and inducing large amounts of biofilm in these strains. The possibilities studied were different for both species and were chosen after reviewing the literature.
For Salmonella spp., this study was carried out with three different strains. We tested three culture media: TSB diluted 1/20, and Luria Bertani broth (LB). Additionally, each medium was screened supplemented with or without 0.25% glucose (Merck Millipore, Bedford, MA, US). The last condition tried was a pre-fixation step with 100 μL of methanol (VWR Chemicals, Barcelona, Spain) during 15 min before quantification, as a way to attach the film [51,52,53,54].Prior to analysis, tested strains were cultivated on solid medium and transferred into liquid cultures. These bacterial suspensions were made with LB broth (Condalab, Barcelona, Spain) for Gram-negative strains and TSB (Condalab, Spain) for Gram-positive strains. Incubation was carried out overnight at 37 °C with shaking. Overnight cultures in LB were diluted into the corresponding media and the turbidity was adjusted to match 1.5 × 108 CFU/mL (McFarland 0.5) [52]. Plates were filled with 75 μL of the suitable medium and 75 μL of bacterial suspension. Negative control wells containing only broth were included. The inoculated plate was covered with a lid and incubated aerobically 48 h at 37 °C, with the exception of E. coli which was incubated at 30 °C. The planktonic suspension was carefully removed. Then, 150 μL of 1x phosphate-buffered saline (PBS) (Merck Chemicals, Bedford, MA, USA) was added for washing. The content was again poured off in order to remove all non-adherent bacteria and plates were dried for 15–20 min at 65 °C. Afterwards, microtiter plates were stained 10 min with 150 μL of 1% crystal violet (CV) (Comercial Bellés, Tarragona, Spain) per well. Excess dye was vigorously shaken and washed with 150 μL of 1× PBS. Later, empty plates were dried 20–25 min at 65 °C. The attached biofilm stained with crystal violet was dissolved in 100 μL of 33% glacial acetic acid (Panreac, Castellar del Vallés, Spain).Results were obtained by measuring the OD of each well at 580 nm using an Epoch™ Microplate Spectrophotometer (BioTek, Winooski, VT, USA). Average OD values were calculated for all tested bacteria and negative controls. Interpretation of the results was achieved by using a cut-off value of 0.2 (ODc) in order to separate the strains in different categories: OD ≤ ODc = no biofilm producer; ODc 55]. 4.6. Antibiofilm ActivityThe biofilm inhibitory activity of the gold(III) complexes was determined against the biofilm-forming strains included in the study. Serial two-fold dilutions of each gold(III)-complex, ranged from 64 to 0.5 mg/L, were performed in flat-bottomed 96-well microtiter plates with the corresponding culture media to enhance biofilm formation (TSB diluted 1/20 for Salmonella spp., M63 supplemented with 0.25% glucose for E. coli, LB supplemented with 0.25% glucose for P. aeruginosa, TSB with 0.25% glucose for Staphylococcus spp., and TSB with 5% horse-lysed blood for S. uberis). Bacterial suspensions were prepared from overnight cultures, and the turbidity was adjusted to a 0.5 McFarland in the corresponding media for each species and consequently diluted to reach an inoculum of 5 × 106 CFU/well. For P. aeruginosa, biofilms were formed by immersing pegs of a modified polystyrene microtiter lid (Nunc TSP System, Nunc, Rockslide, Denmark). After 48 h incubation at 37 °C in static, biofilms on microtiter plates or pegs were carefully rinsed three times with 1× PBS to discard planktonic cells and exposed at 65 °C until completely dry. Biofilms were then fully covered with a 1% CV stain solution for 10 min at room temperature [52]. The CV was then removed, washed with 1× PBS to eliminate the excess of dye and heat-dried for 60 min. Biofilm formation was quantified by eluting the CV fixed to the biofilm in 33% glacial acetic acid and absorbance of each well was measured at 580 nm using a microplate spectrophotometer (EPOCH, BioTek, Winooski, VT, USA).The minimum biofilm inhibitory concentration (MBIC) was defined as the minimal concentration of the compound that led to a three-fold decrease in absorbance when compared to the growth control values. All the experiments were carried out in triplicate.
4.7. Antimicrobial Synergy StudyTwo-dimensional checkerboard arrays were used to determine the efficacy of the combination of the selected gold(III) complex, 2b, together with colistin (CST), in contrast to their individual activities. This experiment was carried out only with the CST-resistant strain E. coli GN1044, with the aim of observing differences in their resistance profile. Gram positive strains were not included since they are intrinsically resistant to polymyxins.
The setup of each assay plate evaluated two-fold dilutions of selected gold(III) with two-fold dilutions of CST, considering a concentration range that included, in the middle of the gradient, the MIC of each molecule against the tested strain. Inoculum size, culture media and incubation conditions were the same as described above for the MIC microdilution method. Positive and negative controls were also included, and the final volume of the assay was 200 μL per well. The fractional inhibitory concentration index (FICI) was calculated considering the following formula: FICI = FIC A + FIC B, where FIC A is the MIC of drug A in combination/MIC of drug A alone, and FIC B is the MIC of drug B in combination/MIC of drug B alone. Synergy was defined by a FICI ≤ 0.5. Interactions with FICI values ranging from 0.5 to 4 were classified as additive/indifferent. Antagonism was defined by a FICI ≥ 4 [55]. 4.8. In Vitro Cytotoxicity AssayThe cytotoxicity of gold(III) complexes was determined using a colorimetric kit that detects cellular metabolic activities (XTT Cell Proliferation Assay Kit, Canvax Biotech, Córdoba, Spain). The experiment is based on the extracellular reduction of XTT (sodium 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)carbonyl]-2H-tetrazolium) by NADH to a highly colored formazan dye. This only occurs in metabolically active cells as it depends on mitochondrial respiration. Therefore, the amount of formazan produced is proportional to the viable cells in the sample. The experiment was also carried out with DMSO as a control, to see if the solvent caused toxicity. We used Jurkat E6.1 commercial cell line. The cells were maintained in RPMI-1470 supplemented with 10% fetal bovine serum (FBS), penicillin and streptomycin at 37 °C and 5% CO2 atmosphere.
Study was conducted in sterile 96-well microplates and cells were spread to a density of 105 in 100 μL/well. Serial concentrations of the tested compounds were also included in the rows, starting with a range of 64 mg/L. After 24 h incubation with the conditions mentioned before, we added 50 μL of XTT to each well and incubated for an additional 4 h. Measurement of the results were made on the Epoch™ spectrophotometer plate reader at a wavelength of 450–500 nm and 630–690 nm. The first measurement gives the signal absorbance of the samples, and the second gives the background signal.
4.9. Statistical AnalysisData analysis was performed with the statistical software R commander 4.1.0. Regarding the assessment of biofilm formation, determination of the appropriate protocol was carried out using the analysis of variance (ANOVA) test in order to check for significant differences between the conditions. However, our data did not comply with the homoscedasticity assumption (i.e., no equal variances), so heteroscedastic ANOVA were used. This test employs Welch’s correction and allows comparison of normal samples with different variances. After establishing differences between means, we used the Bonferroni posthoc test for pairwise comparisons. Graphics were performed using GraphPad Prism 9 software.
留言 (0)