
記住我





To assess whether LGMD2B causes lipoprotein abnormalities, biobanked serum samples were selected from LGMD2B and control patients. Population characteristics, age and sex distributions, velocity/speed (m/s) measures for the time to run/walk 10-m test (TTRW), and overall ambulatory status (expressed as a %) are listed in Table 1. As functional TTRW scores were only obtained from ambulant patients, and reduced rates of ambulation were observed in LGMD2B cohorts > 30 years of age, any patient listed as nonambulatory or unable to walk 10 m with usual orthotics and walking aids was assigned a velocity or speed value of “0” and included in the analysis, where a reduced walking speed (m/s) is indicative of poorer ambulatory ability.
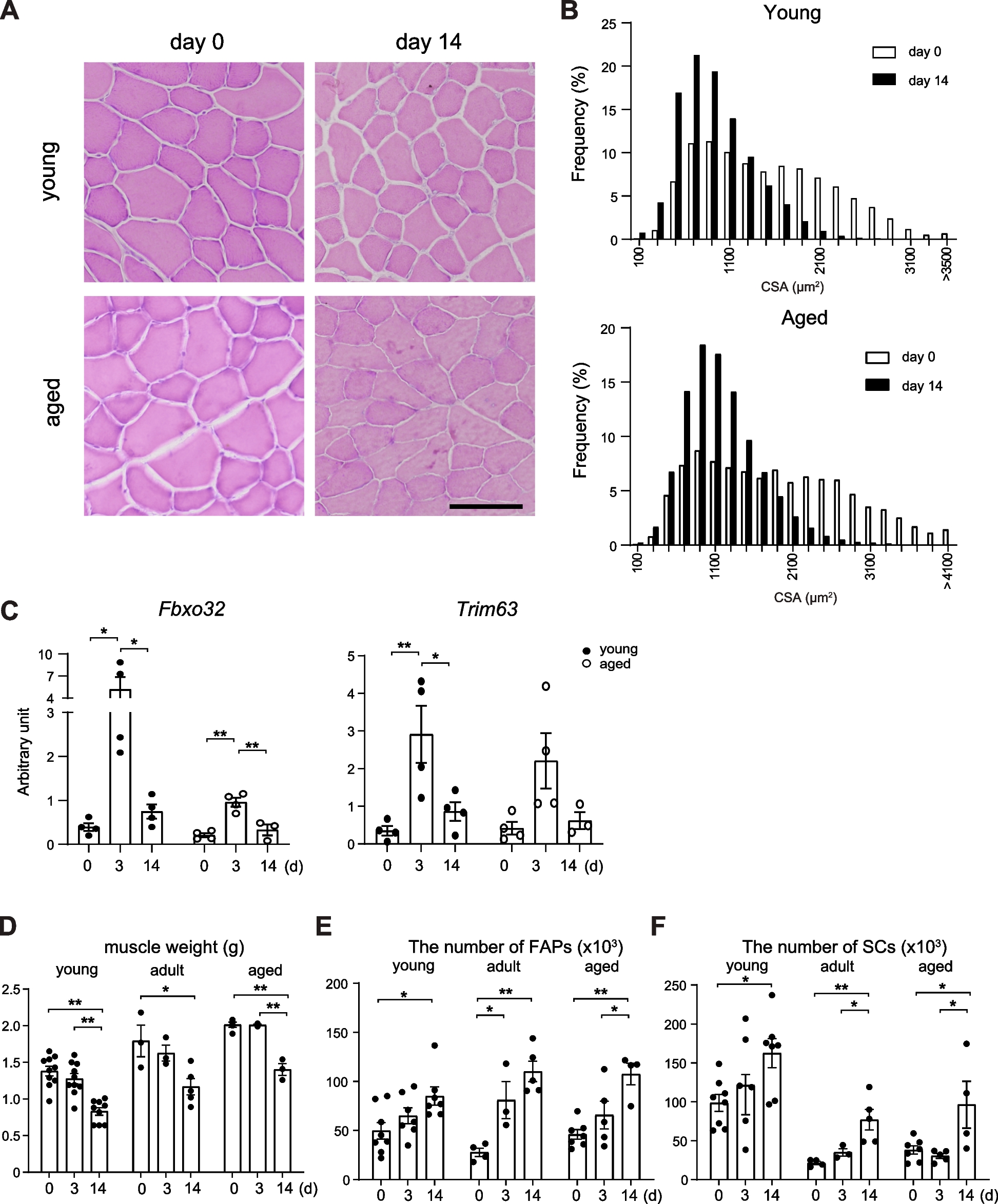
Table 1 Patient characteristics for serum samples of control and LGMD2B patientsAges of LGMD2B patients with biobanked samples were between 22–30 years (young) and 41–51 years (old), with N = 42 and N = 33 samples available in each respective group. Biobanked control serum was also sourced from N = 14 patients aged between 20 and 39 years (young) and N = 16 patients aged between 40 and 55 years (old). The mean ages of patients with available samples were significantly higher in the young control group than the young LGMD2B patients group for both sexes (P ≤ 0.05) but was similar among old cohorts (Table 1). Walking/running speeds in old male and female LGMD2B patients were 54% and 81% lower than that of young patients and were associated with an increased percentage of non-ambulation (Table 1).
LGMD2B patients present with serum HDL-C abnormalitiesWhen serum lipoprotein levels were stratified by sex, both male and female LGMD2B patients displayed statistically similar levels of CHOL, LDL-C, and nonHDL-C compared to their respective controls (Fig. 1A–C). Interestingly, we observed a 21% and 19% reduction in LGMD2B serum HDL-C levels in both male and female LGMD2B patients, respectively (Fig. 1D; P < 0.01). Analysis of CHOL/HDL-C ratios, a typical indicator of cardiovascular disease, demonstrated that 73% of male and 50% of female LGMD2B patients had abnormal CHOL/HDL-C ratios (Fig. 1E). CHOL/HDL-C ratios were also 12% higher in males (ns) and 25% higher in females than their respective controls (P < 0.05; Fig. 1E). TG levels in LGMD2B patients of both sexes were not statistically different from controls (Fig. 1F). When stratified for age (young vs old), LGMD2B patients displayed similar levels of serum CHOL, LDL-C, and nonHDL-C compared to age-matched control samples (Fig. 2A–C). Significant decreases in mean serum HDL-C, however, were observed in young LGMD2B females (by 27%), as well as in older LGMD2B patients regardless of sex (27% male and 15% female, respectively; Fig. 2D). Similarly, CHOL/HDL-C ratios were significantly increased in old (21%; P < 0.05) LGMD2B patients compared to controls, yet both young and old LGMD2B patients showed an unusually high prevalence of ratio abnormalities (55% young and 70% old) (Fig. 2E). TG levels were similar across all cohorts (Fig. 2F).
Fig. 1
Serum lipoprotein and triglyceride distribution in adult control and LGMD2B patients aged ≥ 20 years and stratified by sex. A–F Scatter plots of control and LGMD2B lipoprotein serum lipoprotein (CHOL, LDL-C, nonHDL-C, HDL-C, and CHOL/HDL-C ratio) and TGs showing values that fall within and outside of normal adult levels. Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between control and LGMD2B means; *P < 0.05; *P < 0.01; ***P < 0.001. Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between male and female means; #P < 0.05; ##P < 0.01; ###P < 0.001. Gray zone denotes normal adult range for each specific parameter. The number of patients falling outside of normal range (abnormal values) is listed as a percentage. Mean ± SEM. Male control (N = 15). female control (N = 15). Male LGMD2B (N = 37). Female LGMD2B (N = 38)
Fig. 2
Serum lipoprotein and triglyceride distribution in adult control and LGMD2B patients aged ≥ 20 years and stratified by age. A–F Scatter plots of control and LGMD2B lipoprotein serum lipoprotein (CHOL, LDL-C, nonHDL-C, HDL-C, and CHOL/HDL-C ratio) and TGs showing values that fall within and outside of normal adult levels. A, B, C, and E Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between control and LGMD2B means; *P < 0.05; **P < 0.01; ***P < 0.001. D and F Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between control and LGMD2B means; #P < 0.05; ##P < 0.01; ###P < 0.001. Gray zone denotes normal adult range for each specific parameter. The number of patients falling outside of normal range (abnormal values) is listed as a percentage. Mean ± SEM. A, B, C, and E Young control (N = 14); old control (N = 16); young LGMD2B (N = 42); old LGMD2B (N = 33). D and F Young male control (N = 8); young female control (N = 6); old male control (N = 7); old female control (N = 9); young male LGMD2B (N = 22); young female LGMD2B (N = 20); old male LGMD2B (N = 15); old female LGMD2B (N = 18)
LGMD2B causes subclinical liver enzyme elevationsTo eliminate liver damage as the source of low-serum HDL-C levels in LGMD2B patients, analyses of serum gamma-glutamyl transferase (GGT), the most sensitive enzymatic indicator of liver disease, were performed in the abovementioned samples (Fig. 3). In the context of MD, regular markers of liver abnormality including alanine and aspartate transaminases (ALT and AST) are typically deemed inappropriate, as transaminases are also released by muscle cells in response to damage [26]. When stratified for both age and sex, GGT levels were increased in young male (by 110%; ns), young female (by 122%; ns), old male (by 44%; ns), and old female (123%; P < 0.05) LGMD2B cohorts compared to healthy controls (Fig. 3A). Overall, GGT levels in LGMD2B patients were 59% higher than that of healthy controls (P < 0.05), although only 13% of all LGMD2B patients exhibited values that were considered to be clinically abnormal (Fig. 3B). Moreover, no significant correlation between serum GGT and HDL-C levels was observed (Fig. 3C).
Fig. 3
Comparative analysis of male and female serum gamma-glutamyl transferase (GGT) levels and overall distribution in adult control and LGMD2B patients ≥ 20 years of age. A Scatter plot of control and LGMD2B serum GGT stratified for both age and sex. Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between control and LGMD2B means; *P < 0.05. B Scatter plot of combined control and LGMD2B serum GGT. Unpaired t-test was used to compare means; *P < 0.05. Gray zone denotes normal clinical adult range. C Correlations between serum GGT and HDL-C levels in control and LGMD2B patients. Pearson correlation R2 and P-values are listed were applicable. Mean ± SEM. A Young male control (N = 7); young female control (N = 5); old male control (N = 7); old female control (N = 8); young male LGMD2B (N = 20); young female LGMD2B (N = 19); old male LGMD2B (N = 13); old female LGMD2B (N = 18). B–C Control (N = 27); LGMD2B (N = 70)
Simvastatin did not attenuate murine LGMD2B-associated disease pathology despite showing biological activitySince statin inhibition of muscle wasting in dystrophin-deficient mdx models of DMD remains controversial [15, 16], we tested simvastatin in the Dysf-null model of LGMD2B between 2 and 11 months, under both normolipidemic chow and hyperlipidemic HFD conditions. In Dysf mice, oral administration of simvastatin prevented HFD-induced weight gain (Fig. 4A), owing to both a reduction in epidydimal fat accumulation and liver enlargement (Supp. Fig. 2), yet simvastatin had no effect on ambulatory function and step length readouts (Fig. 4B). Notably, simvastatin did not change CHOL, HDL-C, or LDL-C levels in chow-fed mice and had a minor effect on TGs (Fig. 4C–F). In HFD-fed animals, Dysf mice showed lower levels of CHOL, HDL-C, LDL-C, and TGs than their WT controls (Fig. 4C–F), and simvastatin induced a 46% and 43% reduction in plasma CHOL and HDL-C but left TG levels unaffected (Fig. 4C–E). Direct comparisons between vehicle treated WT and Dysf-null lipid levels have been included in Supp. Fig. 2A–F. No discernable effects of simvastatin were observed on body, fat pad, or liver weights in chow-fed mice or WT controls on either diet (Supp. Fig. 3A–B).
Fig. 4
Simvastatin mitigates HFD-induced weight gain and plasma lipoprotein accretion in Dysf-null mice but fails to prevent ambulatory dysfunction. A Monthly body weight measures (2, 4, 6, 8, and 11 months) in Chow and HFD-fed, vehicle, and simvastatin (SIM)-treated WT and Dysf mice. Two-way ANOVA with Tukey’s post hoc tests were used for direct mean comparisons; #P < 0.05 and compares WT SIM and Dysf SIM at the age group selected. **P < 0.01 and ***P < 0.001 independently compare both WT SIM to Dysf SIM and Dysf vehicle to Dysf SIM across the age groups selected. B End-point (11 m) step length images from Chow and HFD-fed Dysf mice treated with either vehicle or simvastatin and their quantification for all groups. Two-way ANOVA with Sidak’s post hoc tests were used for direct mean comparisons between; ••P < 0.01 and within; no significance detected. C–E End-point total plasma cholesterol (CHOL), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C) and triglycerides (TG) from Chow and HFD-fed, WT and Dysf-null mice treated with either vehicle or simvastatin. Two-way ANOVA with Sidak’s post hoc tests were used for direct mean comparisons between, •••P < 0.001 and ••••P < 0.0001, and within, *P < 0.05 and ****P < 0.0001. Mean ± SEM. A–BN = 6–9. C–FN = 8–13
Despite profoundly affecting body composition and key metabolic parameters, which confirmed biological activity, simvastatin failed to prevent LGMD2B-associated muscle wasting in quadriceps and triceps muscles under either dietary condition (Table 2). Moreover, in quadriceps and triceps muscle sections stained with Masson’s trichrome, simvastatin was unable to mitigate LGMD2B-associated fat infiltration, muscle damage, and collagen deposition in chow and HFD-fed Dysf mice (Figs. 5A–B and 6A–B). No detrimental effects of simvastatin on muscle pathology were observed in age- and diet-matched WT mice (Figs. 5A–B and 6A–B).
Table 2 Effect of HFD-feeding and simvastatin treatment on relative muscle weights in C57BL/6 and Dysf-null mice aged 11 monthsFig. 5
Simvastatin did not attenuate LGMD2B-associated quadriceps or triceps histopathology in normolipidemic Chow-fed Dysf-null mice. A Representative images of and B quantification of percentage fat, damaged and healthy myofiber areas, and the percentage of collaged deposition in whole quadriceps and tricep muscles from Chow-fed WT and Dysf mice treated with either vehicle or SIM. Scale 1 mm for whole muscle images and 200 μm for insets. Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between WT and Dysf mice; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. Mean ± SEM. A–BN = 7–8
Fig. 6
Simvastatin did not attenuate LGMD2B-associated quadriceps or triceps histopathology in hyperlipidemic HFD-fed Dysf-null mice. A Representative images of and B quantification of percentage fat, damaged and healthy myofiber areas, and the percentage of collaged deposition in whole quadriceps and tricep muscles from HFD-fed WT and Dysf mice treated with either vehicle or SIM. Scale 1 mm for whole muscle images and 200 μm for insets. Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between WT and Dysf mice; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. Mean ± SEM. A–BN = 7–9
Dysf deficiency causes overexpression of cholesterol metabolism regulators in skeletal muscle but not the liverTo further investigate the role of cholesterol in LGMD2B muscle pathology, we measured muscle protein levels of HMGCR and LDLR, the main regulators of nonHDL-C cholesterol metabolism and effectors of statin bioactivity. Western blot analysis demonstrated that HMGCR and LDLR protein expression was increased in gastrocnemius muscles of chow and HFD-fed Dysf mice compared to diet-matched WT controls, and simvastatin treatment failed to normalize intramuscular levels of either protein (Fig. 7B). These data were further corroborated using immunofluorescence tissue staining for both HMGCR and Filipin, the latter of which detects cholesterol-rich domains as well as intracellular levels of unesterified-free cholesterol [15]. Filipin staining was significantly elevated in Dysf-null quadriceps muscle sections compared to WT under both chow (by 380%; P < 0.001) and HFD-fed conditions (by 775%; P < 0.001) (Fig. 8A–B). Similarly, levels of HMGCR were elevated by 87% (P < 0.05) and 94% (ns) in the quadriceps muscles of Chow and HFD-fed Dysf-null mice compared to WT controls, respectively (Fig. 8A–B). Intramuscular levels of either protein were again not significantly reduced following simvastatin treatment (Fig. 8A–B). In the liver, the main site of whole-body cholesterol regulation, we observed that in contrast to skeletal muscle, neither HMGCR nor LDLR were affected by genotype, diet, or simvastatin treatment (Fig. 9A–B). Combined, these data suggest that loss of dysferlin interferes with normal muscle expression of mevalonate/HMGCR and LDLR, two key regulators of cholesterol metabolism previously shown to be both elevated in mdx muscle tissues and reduced with simvastatin [15].
Fig. 7
Simvastatin failed to downregulate HMGCR and LDLR signaling in skeletal muscles in Chow or HFD-fed Dysf-null mice. A Representative images and immunoblot quantification of HMGCR and LDLR in gastrocnemius muscles of Chow-fed, WT and Dysf mice treated with either vehicle or simvastatin. Two-way ANOVA with Sidak’s post hoc tests were used for direct mean comparisons; *P < 0.05, **P < 0.01, and ****P < 0.0001. B Representative images and immunoblot quantification of HMGCR and LDLR in gastrocnemius muscles of Chow-fed, WT and Dysf mice treated with either vehicle or simvastatin. Two-way ANOVA with Sidak’s post hoc tests were used for direct mean comparisons; *P < 0.05, **P < 0.01, and ****P < 0.0001. HMGCR and LDLR were standardized to the loading control GAPDH. Y-axes represent arbitrary units (AU). Full blots were imaged separately and thus have differing exposures. A common sample was loading onto each gel to normalize for detection efficiencies across membranes. Mean ± SEM. N = 5 for each cohorts
Fig. 8
Simvastatin failed to normalize intramuscular levels of Filipin and HMGCR in the quadriceps of Chow or HFD-fed Dysf-null mice. A Representative images of Filipin and HMGCR in both Chow and HFD-fed, WT and Dysf mice treated with either vehicle or simvastatin. B Quantification of the average fluorescent intensity of Filipin and HMGCR. Relative intensity values were generated by normalizing to WT (Chow) levels to account for detection efficiencies. Scale 50 μm in both main images and insets. Insets: no Filipin control (DRAQ7 only) and no HMGCR control DAPI only). Two-way ANOVA with Sidak’s post hoc tests were used for direct comparisons between WT and Dysf mice; *P < 0.05; **P < 0.01; ***P < 0.001. Mean ± SEM. N = 4–7 per cohort
Fig. 9
No effect of simvastatin on the liver expression of HMGCR or LDLR. A Representative images and immunoblot quantification of HMGCR and LDLR in the livers of Chow-fed, WT and Dysf mice treated with either vehicle or simvastatin. Two-way ANOVA with Sidak’s post hoc tests were used for direct mean comparisons; no significance. B Representative images and immunoblot quantification of HMGCR and LDLR in the livers of Chow-fed, WT, and Dysf mice treated with either vehicle or simvastatin. Two-way ANOVA with Sidak’s post hoc tests were used for direct mean comparisons; no significance. HMGCR and LDLR were standardized to the loading control GAPDH. Y-axes represent arbitrary units (AU). Full blots were imaged separately and thus have differing exposures. A common sample was loading onto each gel to normalize for detection efficiencies across membranes. Mean ± SEM. N = 3 for each cohort
Simvastatin treatment had no effect on mTORC1 or AMPK pathway activation in muscle lysatesmTORC1 signalling is a key regulator of protein synthesis in skeletal muscle. The activation of mTORC1, either downstream of AKT or directly by nutrients, can promote protein synthesis by phosphorylating two major targets: S6K1 and 4E-BP1, which can also be assessed downstream by rpS6(Ser235/236) expression [27]. Since simvastatin has been shown to inhibit the phosphorylation of rpS6 (Ser235/236; p-rpS6) and mTORC1 signalling in skeletal muscle and contribute to statin-induced myopathy [28], we therefore measured rates of p-rpS6 activation in skeletal muscle lysates of WT and Dysf-null mice treated with simvastatin. Western blot analysis demonstrated that simvastatin had no effect on levels of p-rpS6(Ser235/236) levels when standardized to total rpS6 (t-rpS6) (Supp. Fig. 4A and B). Levels of total rpS6 levels (t-rpS6) standardized to GAPDH in Dysf-null tissues were also unaffected by simvastatin treatment, despite displaying increased t-rpS6 protein standardized to GAPDH when compared to WT muscle (Supp. Fig. 4A and B). While AMPK phosphorylation is an inhibitory regulator of the mTORC1 pathway [29], in other tissues (e.g., liver), the phosphorylation of AMPKα(Thr172) can also directly downregulate HMGCR activity and thus inhibit intracellular rates of cholesterol synthesis [30], although it should be noted that studies describing AMPK-mediated inhibition of cholesterol synthesis are limited. Similarly to mTORC1 activation, levels of p-AMPKα(Thr172) standardized to t-AMPKα were not significantly affected by simvastatin treatment in WT and Dysf-null mice fed either a chow or HFD (Supp. Fig. 4A and C). While total amounts of AMPKα standardized to the loading control GAPDH were significantly elevated in both chow and HFD-fed Dysf-null muscles compared to WT (by 113% and 200%, respectively), expression levels were not sufficient to antagonize intramuscular cholesterol synthesis (nor affect mTORC1 basal activation) (Supp. Fig. 4A and C). Furthermore, levels of t-AMPKα were not affected by simvastatin treatment (Supp. Fig. 4A and C).
留言 (0)