
記住我

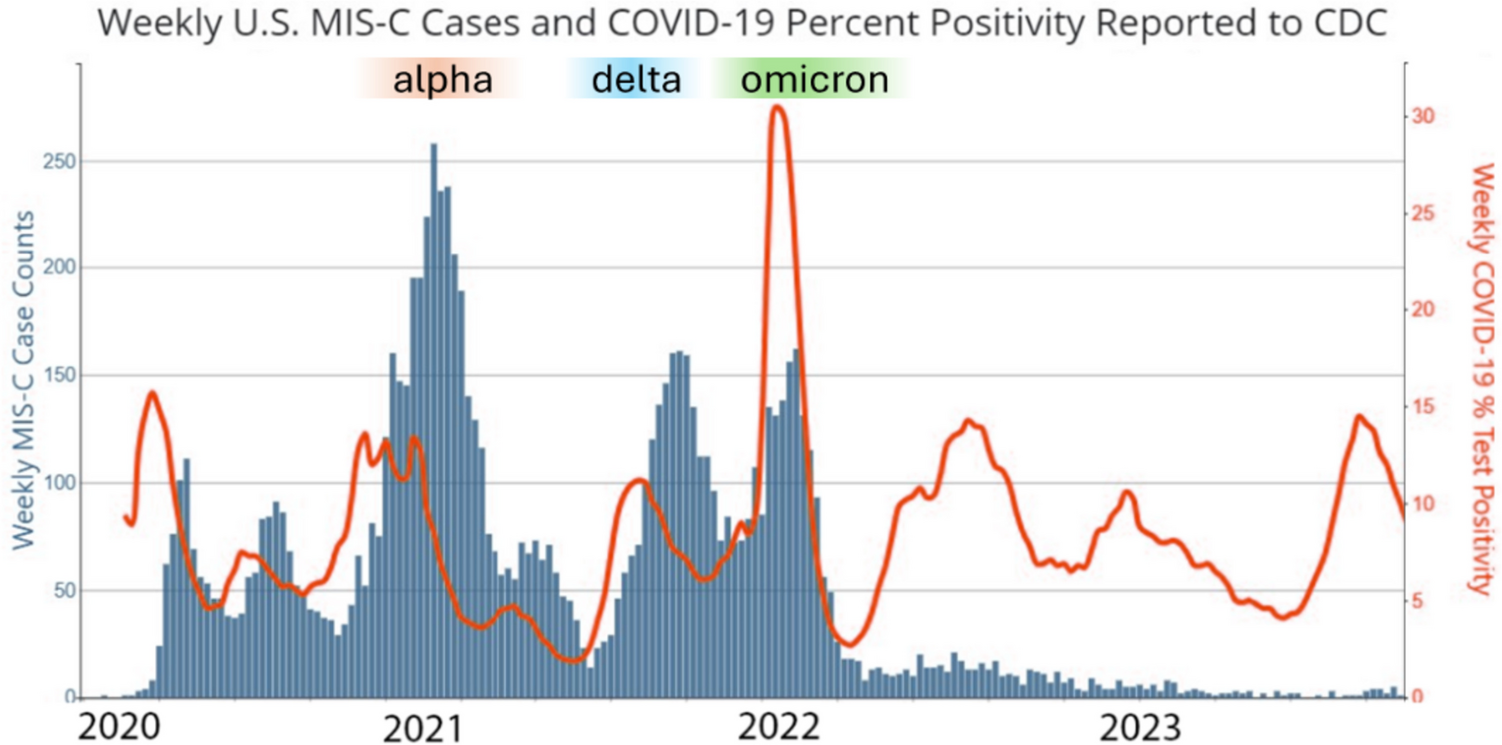

In the last 3 years, immune-metabolic research in allergology had a clear focus on asthma with some studies also reporting findings in food allergy and atopic dermatitis.
AsthmaAsthma is a complex disease comprising different phenotypes that are characterized by airway-inflammation, -remodeling, and -hyperreactivity. Studies have repeatedly shown the presence of metabolic anomalies in asthmatic patients and many metabolic pathways, including glycolysis [36,37,38,39,40], amino acid metabolism [41, 42], fatty acid metabolism [43, 44], and sphingosine metabolism [45,46,47], have been reported to be associated with asthma. Especially obese asthma, which trends towards higher disease severity, is associated with increases in glycolysis and both basal and maximal respiration in airway epithelial cells and platelets [48], as well as overall increased oxidative stress [48, 49].
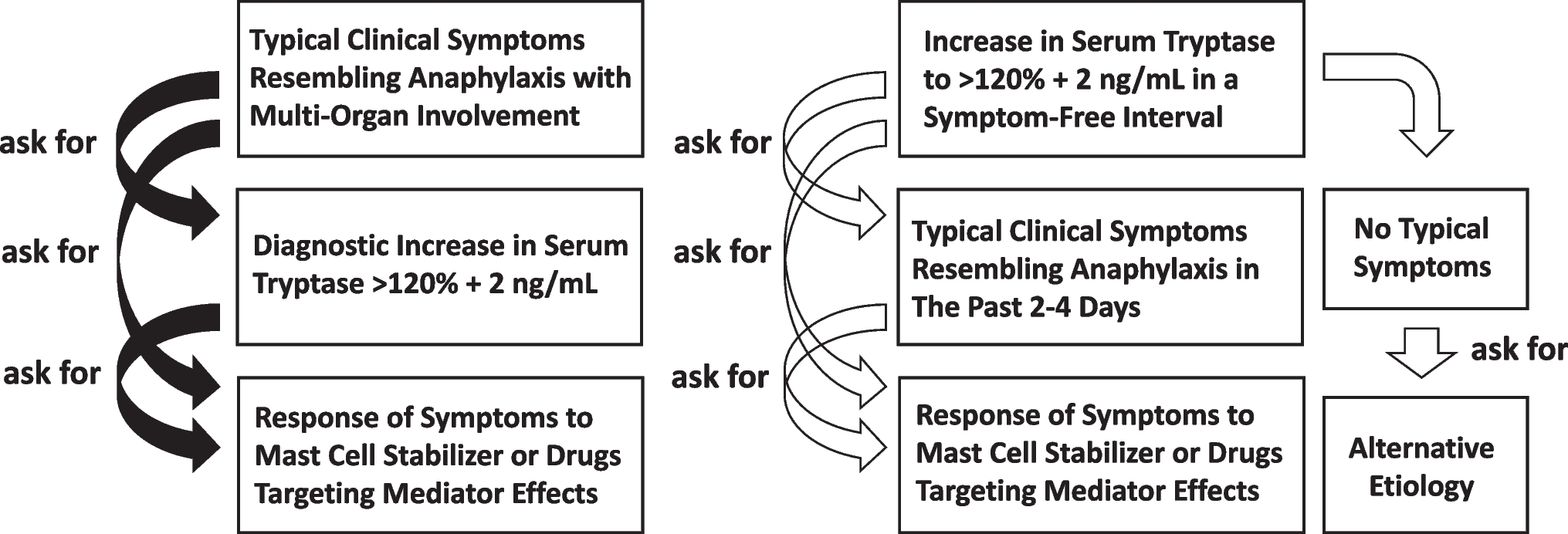
Over the past years, increased rates of glycolysis were reported in bronchial epithelial cells from either obese or asthma patients [48] and glycolytic reprogramming has emerged as a key parameter of allergic airway disease implicated in the development of mucus metaplasia, airway inflammation, and airway hyperreactivity (Fig. 2) [36, 37].
Fig. 2
Metabolic adaptions observed in bronchial epithelial cells of asthmatic patients and their contribution to allergic inflammation. Bronchial epithelial cells were repeatedly shown to display increased rates of glycolysis with an increased secretion of lactate, a Warburg Effect, and a disrupted Krebs cycle. Here, the dimeric, non-glycolytic isoform of the enzyme pyruvate kinase isozyme M2 (PKM2, which catalyzes the last step of glycolysis and is an important regulator of overall glycolytic flux) triggered by house dust mite (HDM) exposure, was shown to promote the STAT3-dependent production of pro-inflammatory cytokines. Moreover, the macrophage migration inhibitory factor (MIF), triggered again by HDM exposure, was shown to also promote glycolysis, resulting in barrier disruption and release of pro-inflammatory mediators. Signaling via the long isoform of TSLP binding to the TSLPR/IL-7R-complex was shown to promote allergic inflammation via the JAK/STAT-pathway resulting in the activation of miRNA-233, VHL, and HIF-1α. Also, amino acid metabolism was shown to contribute to the secretion of increased amounts of NO. Inflammatory mediators (NO; ROS; Th2-cytokines and IL-1β) secreted by the bronchial epithelial cells in turn contribute to the activation of other immune cells (e.g., Th2 cells and monocytes, which were also shown to display enhanced rates of glycolysis), the production of IgE antibodies, finally mediating airway inflammation and airway hyperreactivity. For more information see text. Abbreviations: HDM, house dust mite extract; VHL, von Hippel-Lindau; PEP, phosphoenolpyruvate; HIF-1α, hypoxia-inducible factor 1α; JAK, Janus kinase; STAT(3), signal transducer and activator of transcription (3); MIF, macrophage migration inhibitory factor; lTSLP, long isoform of thymic stromal lymphopoietin; TSLPR, TSLP receptor; ROS, reactive oxygen species; NO, nitric oxide
Recently, serum lactate levels were found to be significantly elevated in clinically stable asthmatic patients and chronic obstructive pulmonary disease patients compared to healthy controls. Moreover, CD4+ T cells derived from either asthmatics or a mouse model of ragweed asthma produced higher amounts of lactate upon stimulation. Interestingly, lactate stimulated T cell proliferation at low concentrations, whereas at high concentrations, it inhibited proliferation. Inhibiting aerobic glycolysis using dichloroacetate (which inhibits pyruvate dehydrogenase kinase 1 (PDK1)) inhibited lactate production, T cell proliferation, production of T cell cytokines (IL-5, IFN-γ IL-17), and airway inflammation while at the same time promoting IL-10-secreting regulatory T cells (Tregs) [50].
Niu et al. showed monocytes from asthmatic patients and lungs from ovalbumin-sensitized and challenged mice to display increased lactate production and enhanced activity of glycolytic enzymes (Fig. 2). This enhanced level of glycolysis was accompanied by decreased ATP production and a reduced activity of mitochondrial respiratory chain complexes I and III, suggesting a switch from OxPhos to aerobic glycolysis in asthma. Interestingly, by acting at upstream regulatory elements and regulating co-factor production, hydrogen (H2) was able to reduce airway inflammation by reducing both the activity of glycolytic enzymes and HIF-1α activation while stimulating the activity of OxPhos enzymes [51]. Therefore, hydrogen was able to regulate metabolic programming by reversing the metabolic switch towards aerobic glycolysis.
The establishment and maintenance of asthmatic lung inflammation critically depends on the production of Th2-promoting cytokines such as thymic stromal lymphopoietin (TSLP) [52]. TSLP exists as two distinct isoforms, a long form (lTSLP) and a short form (sTSLP) [53, 54]. Yu et al. found that levels of lTSLP were significantly increased in asthma airway epithelial cells promoting miRNA-233/Von Hippel-Lindau (VHL)/HIF-1α-dependent pro-inflammatory cytokine production and aerobic glycolysis from airway epithelial cells, whereas levels of sTSLP were decreased (Fig. 2). Interestingly, inhibition of glycolysis significantly decreased inflammatory cytokine levels (IL-25 and IL-33), suggesting glycolysis to be involved in asthmatic lung inflammation [55].
In contrast, sTSLP reduced both inflammation of asthmatic airways (reduced airway hyperreactivity, Th2 cytokine production, and IgE production) and aerobic glycolysis in mice by decreasing lactate dehydrogenase A (LDHA) and lactic acid levels in bronchoalveolar lavage fluid (BALF), as well as HIF-1α- and LDHA-protein levels in airway epithelial cells of an asthma mouse model. Mechanistically, both lTSLP and sTSLP induced formation of TSLPR and IL-7R receptor complexes. However, while lTSLP induced phosphorylation of JAK1, JAK2, and STAT5, sTSLP induced little phosphorylation of JAK1 and STAT5. Therefore, it was suggested that sTSLP suppressed inflammation and aerobic glycolysis in asthmatic airway epithelial cells by occupying the binding site of lTSLP in the TSLPR/IL-7R receptor complex [55].
Transforming growth factor beta (TGF-β) was also shown to activate the PI3K-Akt-mTOR pathway in lung myofibroblasts which in turn activates activating transcription factor 4 (ATF4) driving increased expression of the serine-glycine synthesis pathway enzymes that result in excessive deposition of collagen proteins and scarring in idiopathic pulmonary fibrosis. In this context, TGF-β was also shown to increase both glycolysis and mitochondrial oxygen consumption in lung fibroblasts. Here, increased glycolytic flux promoted the conversion of the glycolytic intermediate 3-phosphoglycerate into glycine, the most abundant amino acid found in collagen protein [56].
Macrophage migration inhibitory factor (MIF) was originally described as a T cell-derived protein that could prevent macrophage migration thereby modulating inflammatory responses [39]. Interestingly, MIF overexpression was detected in BALF, serum, and sputum of asthmatic patients where it correlated with the release of pro-inflammatory cytokines TNF-α, IL-1β, and IL-6 [39, 57]. In 16HBE cells, Lan et al. found that house dust mite (HDM) application triggered expression of MIF which in turn promoted epithelial cell barrier disruption and release of Th2-cytokines by accelerating aerobic glycolysis (Fig. 2). Inhibition of MIF in vivo reduced airway hyperactivity, airway inflammation, and IgE production in a chronic mouse asthma model [39].
In line with an increased glycolytic metabolism in asthma, Qian et al. showed primary nasal epithelial cells from asthmatic patients to produce more lactate compared to healthy controls and higher lactate levels in asthmatic sputum samples [36].
In a murine model of allergic asthma, the increased rates of glycolytic flux, glucose consumption, expression of glycolysis genes, and lactate production were found to be dependent on IL-1(α/β). Moreover, enhanced glycolysis was shown to be required for HDM-induced Th2 cytokine release from tracheal epithelial cells. Mechanistically, both the inhibition of IκB kinase ε and the increases in the glycolytic enzyme LDHA were required for HDM-induced glycolysis and pathogenesis of allergic airway disease [36].
Pyruvate kinase M2 (PKM2) is the enzyme catalyzing the last step of glycolysis that also acts as an important regulator of overall glycolytic flux. PKM2 exists in two isoforms, a tetrameric isoform that converts phosphoenolpyruvate to pyruvate and a dimeric isoform with alternative non-glycolytic functions enhancing the transcription of several pro-inflammatory cytokines as a transcriptional coactivator (Fig. 2) [58, 59]. In a recent follow-up study, van de Wetering et al. showed the dimeric form of PKM2 to play a crucial role in the pathogenesis of allergic airways disease by increasing signal transducer and activator of transcription 3 (STAT3)-dependent IL-1β–induced pro-inflammatory signaling in a mouse model of HDM-induced asthma (Fig. 2) [37]. Consequently, TEPP46, a small molecule activator of PKM2, which stabilizes the glycolytic tetrameric PKM2 isoform, decreased IL-1β–mediated airway inflammation as well as expression of pro-inflammatory mediators and attenuated airway eosinophils and mucus metaplasia [37].
In a follow-up study, the same authors showed that glutathione-S-transferase P (GSTP) contributes to IL-1β-induced glycolysis and pro-inflammatory signaling in epithelial cells by S-glutathionylation-mediated disruption of the glycolytic activity of the PKM2 tetramer [60]. In line with their findings from the mouse asthma model, GSTP levels were increased in the sputum of severe asthma patients, and sputum proteomics and transcriptomics also showed strong correlations between GSTP, PKM2, and glycolysis, pointing to a putative PKM2-GSTP-glycolysis signature associated with severe asthma [60].
In aggregate, these results suggest enhanced glycolysis to be critically important for the amplification of allergen-induced pro-inflammatory responses and highlight the importance of the GSTP/PKM2-axis in regulating this process, suggesting GSTP/PKM2 as a novel potential target for the development of IL-1β-associated, glycolysis in asthma [37, 60].
Their results were independently confirmed by Manuel et al., who showed HDM to trigger PKM2-dependent glycolytic reprogramming and airway inflammation in a murine model of obese allergic asthma and airway epithelial cells [38]. These results suggest alternative, non-glycolytic functions of PKM2 to exert pro-inflammatory roles in asthma by glutathione-dependent protein oxidation via a putative IFN-γ–glutaredoxin 1 pathway [38].
Besides increased glycolysis, fatty acid metabolism was repeatedly described to be altered in asthmatic patients [61, 62]. Over the last years, several studies compared asthmatics to healthy controls using either metabolomics or transcriptomics approaches.
Zhu et al. performed an unbiased metabolomics approach with BALF samples of 12 patients undergoing an acute inflammatory response that was experimentally induced by bronchial allergen challenge [63]. Interestingly, unbiased associations of 549 metabolites changed by allergen challenge identified two distinct metabolic profiles in a group of allergic asthmatics that were otherwise indistinguishable under basal conditions [63]. Here, among other pathways relevant to asthma, especially an upregulation of saturated fatty acid synthesis and mitochondrial beta-oxidation of saturated fatty acids was observed upon allergen challenge [63].
Johnson et al. quantified lipid mediators in the nasal airway epithelium in three non-atopic controls, four mild to moderate asthmatics, and four severe asthmatics by LC–MS [64]. In line with an increase in fatty acid metabolism in asthmatics, they found lower levels of pro-resolving 15-HEPE, 19,20-DiHDPA, RvD5, 14-HDHA, 17-HDHA, and 13-HOTrE. In contrast, levels of mainly pro-inflammatory prostaglandin E2 were increased in asthmatic samples, suggesting the levels of lipid mediators in nasal epithelium to distinguish asthma patients form healthy controls [64].
Eosinophils are critical for the local production of cys-leukotrienes in asthma [65]. In line with the higher production of the pro-inflammatory lipid mediator prostaglandin E2 observed by Johnson et al., Miyata et al. reported nasal polyp-derived eosinophils from patients with eosinophilic rhinosinusitis to present a characteristic fatty acid metabolism with selectively higher production of leukotriene D4 [66].
Confirming higher rates of fatty acid metabolism in asthmatics, bronchial smooth muscle (BSM) from asthmatic patients was characterized by a switch towards increased fatty acid consumption and increased rates of mitochondrial respiration [67]. In line with these results, the authors observed a significant decrease in arachidonic-, stearic-, and linoleic acid content in asthmatic BSM cells compared to non-asthmatic control cells. Interestingly, in this study, no differences in glucose uptake were observed between asthmatic and non-asthmatic BSM cells [67]. Esteves et al. also found increased levels of two molecules involved in the internalization of fatty acids, carnitine palmitoyl transferase-2 (CPT2) and LDL-receptor, in asthmatic BSM cells [67]. Here, blocking of either CPT2 or LDL-receptor strongly reduced BSM cell proliferation, confirming the reliance of asthmatic, hyperproliferative BSM cells on increased FAO and suggesting fatty acid metabolism as a novel treatment target.
Ravi et al. performed unbiased transcriptome analyses of bronchial epithelial cells (BECs) from either mild, moderate, or severe asthma patients to investigate metabolic adaptations in asthma [68]. Patients with severe asthma displayed a reduction in OxPhos genes, while for patients with mild asthma, this reduction was less pronounced and more heterogeneous [68]. In contrast to this, both genes related to fatty acid metabolism and certain lipid species (PCs, LPCs, lysophosphatidylethanolamines, and BMP) were significantly upregulated in BECs from all asthma patients independently of asthma severity, clearly differentiating asthma patients from healthy controls and demonstrating a persistently altered fatty acid metabolism in asthmatic bronchial epithelium [68]. This metabolic shift possibly affecting purine metabolism, amino acid biosynthesis, and glycolysis was reversed in patients who underwent asthma treatment with bronchial thermoplasty, resulting in a gene expression profile more comparable to healthy controls [68].
In a mechanistic study, Lechner et al. described macrophages derived either from asthma patients, HDM-sensitized mice, or macrophages trained with HDM extract in vitro to produce high amounts of CCL17 and cysLTs, both key mediators involved in asthma while not generally increasing the production of pro-inflammatory cytokines [69]. This allergen-driven trained immunity program depended on formyl peptide receptor 2 (FPR2)- and TNF-signaling and resulted in metabolic reprogramming and lysine demethylase (KDM) 1A–mediated histone demethylation (particularly H3K4 and H3K9) and the potential removal of repressive marks to enhance Th2 responses [69]. Therefore, this trained immunity response may contribute to both chronification and exacerbation of allergic asthma and could be a promising target for preventing asthma.
Polyunsaturated fatty acids (PUFAs) were repeatedly shown to have health-promoting effects [70, 71]. Stearoyl-coenzyme A desaturase (SCD) is the rate-limiting enzyme in the synthesis of PUFAs [72]. Impaired fatty acid metabolism was suggested to contribute to asthma pathology [43, 73, 74]. Rodriguez-Perez et al. observed significantly reduced serum fatty acid levels (palmitoleic acid, arachidonic acid, and docosahexaenoic acid) and a suppressed polyunsaturated:saturated fatty acid ratio in serum of non-obese asthma patients, suggesting reduced desaturase activity in non-obese asthma patients with severe disease [62]. Their results were confirmed in human bronchial epithelial cells and both mouse OVA and HDM models where animals also showed both reduced desaturase activity and reduced SCD expression in the lung [62]). Here, inhibition of SCD promoted airway hyper-responsiveness in mice and reduced anti-viral defense in bronchial epithelial cells [62]. Taken together, their results suggested that SCD gene expression and therefore, the levels of potentially anti-inflammatory PUFAs may be regulated by inflammatory responses in the lung. Therefore, SCD may be a promising target for the therapy of asthma.
Besides glucose and fatty acid metabolism, amino acid metabolism was also reported to be altered in asthma. Arginine metabolism in particular was identified as a key driver of Th2 airway inflammation [75] and to affect the development of asthma [75,76,77]. Arginine is a semi-essential amino acid that is metabolized to generate NO by several enzymes, including the cytosolic inducible nitric oxide synthase (iNOS), the mitochondrial arginase-2 (ARG2), and ARG1 which mostly expressed in the cytosol of hepatic cells [78, 79]. ARG2 gene variants are among the first and most consistently found single nucleotide polymorphisms (SNPs) associated with asthma in genome-wide association studies (GWAS) [80,81,82].
In line with these results, expressions of iNOS and ARG1/2 were found to be upregulated in asthmatic patients, leading to higher levels of exhaled NO and more severe symptoms (Fig. 2) [75,76,77]. Interestingly, patients with high levels of exhaled NO were reported to also have increases in airway lactate levels, suggesting a link between enhanced levels of glycolysis, arginine metabolism, and exhaled NO in some asthma phenotypes [77].
In this context, Asosingh et al. showed arginine metabolism to be a critical modulator of the severity of inflammation and remodeling in eosinophilic and neutrophilic asthma [83]. In their asthma cohort, ARG2 variants associated with lower arginase activity, combined with high levels of fractional exhaled nitric oxide identified a more severe asthma phenotype [83]. In line with these results, ARG2 knock-out mice had the highest levels of airway inflammation, airway remodeling, and lung inflammatory cytokines, suggesting a central role for arginine metabolism in the control of lung inflammation. In line with their results, deletion of ARG2 was shown to increase HIF- and STAT6-signaling resulting in increased eosinophilic Th2 inflammation and Th2 cytokine production [75].
In an independent study, Zhou et al. identified decreased levels of L-citrulline and L-arginine in the serum of 30 young patients with chronic persistent asthma [84], suggesting the balance in arginine metabolism to be important for the pathogenesis and progression of chronic asthma. They suggested that therapeutic supplementation of L-citrulline could restore airway NO production and therefore reduce inflammation [84].
Using ultra-high performance liquid chromatography–mass spectrometry (UHPLC-MS), Liu et al. characterized metabolic profiles in induced sputum samples from both healthy controls and asthmatic patients [85]. They found different inflammatory asthma phenotypes to have specific metabolic profiles with 77 differential metabolic signatures which belonged to five underlying pathways (histidine metabolism, glycerophospholipid metabolism, nicotinate and nicotinamide metabolism, and linoleic acid metabolism as well as phenylalanine-, tyrosine-, and tryptophan-biosynthesis) [85]. Here, levels of adenosine 5′-monophosphate, allantoin, and nicotinamide in sputum samples predicted rate ratios of severe asthma exacerbations [85].
Recently, Bravo-Solarte et al., using bronchial brush samples, described genomic evidence of misregulated glutamine metabolism in airway epithelial cells from asthma patients [
留言 (0)