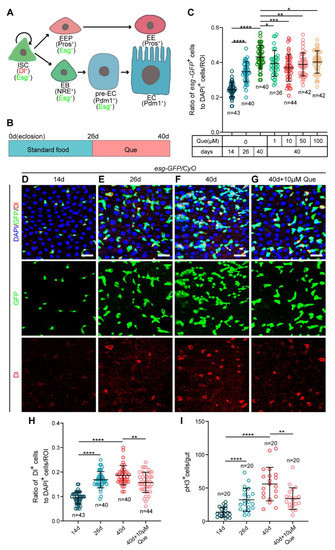
1. IntroductionReactive oxygen species (ROS) represent one of the mainly harmful by-products of cellular metabolism, but are also vital for proper cell functioning as activators of certain signaling pathways [
1]. For instance, the stimulation of non-phagocytic cells with ligands increases intracellular ROS levels [
2], the activation of peptide growth factors via tyrosine kinase [
3,
4], and the cytokine pathways [
5,
6]. While the actual targets of ROS remain largely unexplored, it has been demonstrated that non-lethal concentrations of H2O2 oxidize and thus chemically modify several proteins, including p53, Jun, Fos, and the NF-κB p50 subunit [
7], thereby stimulating (p50) or inhibiting (p53, Jun, Fos) the transcriptional activity.Antioxidants maintain a balance between ROS production and consumption by converting them to harmless reduced species; however, when this balance is disrupted, the elevated ROS levels result in increased DNA, protein, and lipid damage [
7,
8,
9,
10]. This condition, dubbed “oxidative stress”, has been linked to a variety of complex diseases, including various types of cancer [
11,
12,
13,
14], asthma [
15], Crohn’s disease [
16], and multiple sclerosis [
7]. The primary source of cellular ROS represents the leak of electrons from the respiratory or electron transport chain (ETC) located in the mitochondria [
17,
18,
19,
20].The human genome encodes multiple superoxide dismutase (SOD) enzymes that catalyze the transformation of superoxide anions into hydrogen peroxide [
21] to counteract the harmful effects of excessive superoxide anion production. The first line of defense against respiratory chain electron leak forms the manganese superoxide dismutase (MnSOD), located in the mitochondrial matrix, where it scavenges the superoxide anions [
22].The human MnSOD is encoded by the SOD2 gene located on the sixth chromosome in the region 6q25.3 [
23]. In its mature state, the human MnSOD forms a tetrameric enzyme comprised of four identical subunits, each consisting of 198 amino acids and one Mn3+ ion [
24]. The mitochondrial import machinery is required to transport proteins destined for the mitochondria (e.g., MnSOD) across two membranes, the outer and inner mitochondrial membranes [
25]. For proteins to cross the membrane, specific amino acid sequences called mitochondrial targeting sequences (MTSs) are required, which fall into two categories: the N-terminal and internal targeting sequences [
26]. MTSs have been shown to form amphipathic α-helices, which are then recognized by mitochondrial translocases of the outer membrane (TOM) (
Figure 1) [
27,
28,
29]. Upon maturation in mitochondria, the mitochondrial proteins containing N-terminal sequences (e.g., MnSOD) are cleaved of their MTS by the mitochondrial processing peptidase (MPP). It has also been suggested that MTSs adopt context-dependent conformations—α-helical conformation for the recognition by the mitochondrial import machinery, and an extended conformation for the cleavage [
30]. Finally, chaperones fold these proteins into their active form and insert metallocofactors [
31].In 1996, a mutation from cytosine (C) to thymine (T) in the coding region of the SOD2 gene was discovered by two independent studies: Shimoda-Matsubayashi, S. et al. [
32], in the Japanese population, and Rosenblum, J.S. et al. [
33]. This mutation at position 47 of the SOD2 gene results in a substitution of alanine with valine (Ala16Val, also dubbed Ala-9Val) at position −9 of the protein, which corresponds to position 16 in the MTS of MnSOD, as shown in
Figure 2a. In this study, we adhere to numbering II.
Figure 2b depicts the comparison between the MTSs of different primates. Among primates, only the substitution located at position 11 from serine to glycine (S → G) is observed, indicating an otherwise high level of interspecies similarity (for human, the V variant is used in sequence comparisons as this is the canonical sequence deposited as UniProt entry P04179). There are no Uniprot entries containing valine at position 16 among primates other than Homo sapiens. Expanding to other organisms shown in
Figure S3, we observed that only three of the twenty-four examined Uniprot entries included valine at position 16 (12.5%) rather than alanine (87.5%); no other amino acid has been observed at position 16. Moreover, we found that the two adjacent amino acid residues (Pro15 and Leu17) are highly conserved across all examined species (
Supplementary material Figure S3).
The MTS is required for the MnSOD to be correctly localized in the mitochondrial matrix. Upon successful localization, the MTS is cleaved off the mature MnSOD and decays, making it a highly unstable species with an unresolved structure. As a result, the human MnSOD possesses 34 inputs in the online RCSB protein data bank, but only two represent MnSOD in its native state (PDB ID: 2ADQ, 5VF9). Despite a large number of entries, the structure of MnSOD’s MTS has not been determined due to the short-lived nature of the MnSOD precursor. Therefore, we performed molecular dynamics (MD) simulations to reveal the nature of the MTS secondary structures.
Shimoda-Matsubayashi, S. et al. [
32] proposed a typical amphiphilic helical structure of the Ala-MTS and its interruption in the Val-MTS variant using the Chou-Fasman method [
34] and the helical-wheel method [
35]. Moreover, an analysis of 23 MTSs revealed that amphiphilic helices in the MTS may indeed be required for the import of immature MnSOD into mitochondria [
29]. Therefore, the alanine variant of MnSOD should be preferred for mitochondrial transport.However, there is a disagreement about the transport properties of the two immature MnSOD variants. On the one hand, Sutton, A. et al. showed that the Val-MnSOD precursor might be partially arrested in the inner mitochondrial membrane (IMM) and produces 30–40% less active MnSOD than the Ala-MnSOD precursor [
36]. A follow-up study indeed demonstrated four-fold higher levels of the mature exogenous protein of the Ala variant compared to the Val variant in transfected HuH7 human hepatoma cells [
37]. Conversely, Bastakia, M. et al. discovered that the TT (thymine-thymine, translating to valine-valine) and CT (cytosine-thymine, translating to alanine-valine) genotypes of the polymorphic MnSOD rs4880 confer a higher enzymatic activity in human erythrocytes than the CC (cytosine-cytosine, translating to alanine-alanine) genotype [
38].Single nucleotide polymorphism (SNP) rs4880 has been associated with various complex diseases. For instance, the Val allele has been associated with a 10-fold increased risk of heart disease in patients with hereditary hemochromatosis [
39], a 2.3-fold increased risk of developing aggressive forms of prostate cancer in patients consuming more iron [
40], and an increased risk of lung cancer in the Turkish population [
41]. On the other hand, the Ala/Ala genotype has been associated with an increased risk of malignant pleural mesothelioma (MPM) [
42]. In addition, the carriers of the Ala allele have been found to have an increased risk of developing acoustic neuroma [
43], a lower survival rate when treated with cyclophosphamide for breast cancer [
44], an increased risk of intestinal metaplasia [
45], a shorter median overall survival after postoperative chemotherapy with platinum and fluorouracil against gastric cancer [
46], increased ROS-induced DNA damage biomarkers [
47], increased oxidative damage from polyaromatic hydrocarbon (PAH) exposure [
48], and a 4-fold increased risk of breast cancer in premenopausal women [
49]. The rs4880 SNP and its associations with diseases are summarized in
Table 1.Both variants are present natively in the human population (SOD2 gene) according to SNPedia [
50,
51,
52]. Based on the results of our study (
Figure 2b), we adhere to the Ala16Val notation throughout this paper for the reason of clarity.Since the experimental structural data on the MnSOD MTS is currently unavailable, we investigated the effect of the Ala16Val SNP on the secondary structure of the MnSOD MTS via extensive MD simulations. We obtained the initial MTS secondary structural information with PsiPred (v4.0) [
53], as well as by using an online server for protein structure prediction (PEP-FOLD3) [
54]. The resulting MTS 3D structures were then docked to the native MnSOD structure (PDB ID: 5VF9) [
55] using Hdock (13 November 2021) [
56] to form the initial MnSOD MTS structure. The resulting structures were solvated, equilibrated, and propagated in time using 200 ns MD simulations (CHARMM36m) [
57]. The ultimate goal was to evaluate the secondary structure dynamics of the Val variant and compare it to the native Ala variant. 4. Conclusions
This study aimed to decipher the unsolved structure of the MTS of the human MnSOD protein. A stable α-helix was observed for the Ala-MTS residues extending from residue 2 to 20. On the contrary, the α-helix of the Val-MTS collapsed shortly after the beginning of the simulation.
It is well established that an uninterrupted α-helix in the MTS of mitochondrial proteins facilitates their transport into mitochondria [
29], implying that the Ala-MTS should be more effective for transporting the MnSOD precursor into mitochondria. This hypothesis is consistent with the study of Sutton, A. et al. [
36], who showed that the Ala-MnSOD precursor produces up to 40% more of the active MnSOD homotetramer than the Val-MnSOD precursor. However, in that study, the functional activity was determined by examining intensity bands on activity gels, a method that has been criticized for possible accuracy limitations [
75].Another study by Sutton et al. showed that in human hepatoma cells HuH7 transfected with vectors encoding for the human Ala- or Val-MnSOD precursor, the Ala variant produced four-fold more mature exogenous protein and MnSOD activity compared to the Val variant [
37]. Moreover, the results of our study are also partially consistent with the results of the study by Shimoda-Matsubayashi et al. [
32], who proposed an α-helix between residues 10 and 17 of the Ala-MTS and a β-sheet for the Val-MTS. Another study, using I-Tasser [
76] and ab initio modeling [
77], suggested that valine disrupts the α-helix.On the other hand, Bastakia, M. et al. [
38] showed that the genotypes with the T allele (Val-MTS) confer higher enzymatic activity in human erythrocytes than the C allele (Ala-MTS). Two additional studies found increased levels of ROS-induced DNA damage biomarkers for the Ala/Val and Ala/Ala genotypes [
47], and a greater oxidative damage from PAH exposure than Val/Val [
48].Our observations on the valine role in helical structures are supported by Mayer et al., Okamoto et al., and Chakrabartty et al. [
78,
79,
80]. Moreover, Gregoret et al. and Tobias et al. demonstrated that alanine preserves the α-helix in contrast to valine, mainly due to a favorable helix-turn equilibrium observed in MD simulations of peptides [
81,
82]. A similar trend of alanine vs. valine in helix stability was also observed using homo-oligomeric decapeptidides subjected to a Monte Carlo simulated annealing [
83]. As demonstrated by the experimental studies that employed circular dichroism, valine’s role in a helical structure is a result of the β-branching of the side chains [
84,
85,
86,
87], whereas hydrophobicity plays a negligible role [
84,
85]. These results are consistent with the observations from Lyu, P.C. and Jacchieri et al. [
88,
89]. They reported that branched side chains of isoleucine or valine are helix destabilizing, while linear side chain amino acids (e.g., alanine, 2-aminobutyric acid, norvaline, norleucine) are classified as helix stabilizing.Another study that analyzed amino acid distributions in short, medium, and long helices with neighbor-dependent sequence analysis [
90] found that Pro preceding Ala is 50% more common in short helices (four to seven residues) than in proteins. Notably, the MnSOD MTS contains a proline (Pro15) right before the variable residue 16, thus resulting in the Pro15-Ala16 sequence for the Ala-MTS and the Pro15-Val16 sequence for the Val-MTS. However, according to their classification, the MnSOD MTS falls under the long helices category (14 to 22 residues), for which no statistically significant trend between Pro and Ala or Val could be established. Nonetheless, the propensity values are still higher for Pro-Ala (0.52) than for Pro-Val (0.39), even in long helices [
90]. Following Ala16 or Val16 in the MTS is leucine (Leu17). The study found a high propensity for the Ala-Leu sequence in helices (1.81) and a non-statistically significant propensity for the Val-Leu sequence (1.02) [
90], which further supports our findings.
According to the presented computational data, the Ala-MTS prefers a uniform α-helix between residues 2 and 20, while the Val-MTS exhibits helical stability only between residues 13 and 17 for 200 ns. In conjunction with the established principles, the present results, therefore, suggest that the Ala-MTS variant is more easily transported into the mitochondria, resulting in a higher concentration of homotetrameric MnSOD than in the case of the Val-MTS.
The results show that the alanine variant retains the α-helix while the α-helix of the valine MTS variant collapses.



留言 (0)