5.1.1. AlkaloidsAlkaloids are noteworthy chemical compositions that act as a major origin for new medicine. Alkaloids contain nitrogen and heterocyclic rings and are found in different plants. Some alkaloids have only recently managed to become established as anti-cancer medications including vinblastine (1), vinorelbine (2), vincristine (3) and vindesine (4). Vinblastine (1), vincristine (3) and vindesine (4) are indol alkaloids isolated from Catharanthus roseus [
133]. Vincristine (3) and vinblastine (1) are usually utilized in conjunction with other chemotherapeutic factors for the therapy of lymphoma, advanced testicular cancer, leukemia, breast cancer, Kaposi sarcoma and lung cancer [
4].Numerous alkaloids presented activities against human hepatocarcinoma cell line (HepG2) and human colon cancer cell line (LS174T) by modulating autophagy. Vinblastine (1), which was reported to inhibit autophagy maturation, was combined with an autophagy stimulator, nanoliposomal C6-ceramide; this mixture enhances autophagy vacuole accumulation and decreases the autophagy maturation of the treated cells [
134]. Another example of the use of these alkaloids is seen with vincristine and CD24, a protein that is bound to membrane lipid raft microdomains through a glycosylphosphatidylinositol anchor. CD24 is associated with poor prognosis in several forms of tumor tissues [
135,
136,
137]. Furthermore, CD24 was reported to cause chemotherapy resistance [
138,
139,
140] and regulate various signaling pathways in diverse types of tumor cells [
141]. Vincristine (3) was reported to suppress the CD24 expression and eliminate autophagy via suppression of the PTEN-AKT-MTOR complex 1 (MTORC1) pathway in WERI-Rb-1 (HTB-169) and Y79 cell lines. Therefore, the sensitivity of retinoblastoma cells to vincristine (3) increases significantly [
142].Camptothecin (5), a natural compound originally derived from the Asian tree Camptotheca acuminate, was synthesized in 1966 [
143]. A low dose of camptothecin (5) stimulates autophagy through the AMPK-TSC2 (TSC complex subunit 2) MTOR pathway in human CRC cell lines, HCT116 and RKO [
144]. Camptothecin (5) and its derivatives have shown anti-cancer function against several types of tumor cells. It was found that camptothecin (5) enhances MYC/c-Myc expression, which in turn upregulates the expression of UPR sensor proteins in human prostate cancer (LNCaP cells), as well as reactive oxygen species (ROS) generation, which induces autophagy through the Ca2+-AMPK and MAPK/JNK-AP-1 signaling pathways [
145]. Human non-small cell lung cancer (NSCLC) H1299 cells can be sensitized to camptothecin (5) by modulating autophagy. Camptothecin (5) increases autophagosome formation in H1299 cells, in a dose-related mode. The induced autophagy exerts a protective role in camptothecin-treated cells with regard to DNA damage and apoptosis. It was suggested that a combination of camptothecin (5) with an autophagy suppressor may be considered an effective approach for lung cancer treatment [
146].Matrine (6), an alkaloid isolated from Sophora flavescens Ait, has been reported to function as an autophagy inhibitor by blocking autophagic degradation and modulating the maturation procedure of lysosomal proteases in the gastric cancer (GC) SGC-7901 cell line [
147].Berberine (7) is an isoquinoline quaternary alkaloid isolated from many well-known medicinal plants, such as Rhizoma coptidis [
148]. Some synthesized derivatives of berberine (7) with improved efficacy and bioavailability (NAX053, NAX056, NAX057, NAX080 and NAX081) have been reported to increase LC3 lipidation (LC3-II) and consequently are able to stimulate autophagy in human colon carcinoma (HCT116) cells, expressing WT TP53 and in TP53-mutated drug-resistant (SW613-B3) cell lines [
149].Isorhynchophylline (8) is a tetracyclic oxindole alkaloid isolated from Uncaria rhynchophylla. A study showed that this alkaloid results in an enhancement of autophagy markers including LC3-II and stimulates autophagy in an MTOR-independent but BECN1-dependent manner in neuronal cells including SH-SY5Y, PC12 cells and N2a, as well as primary cortical neurons. Moreover, isorhynchophylline (8) can trigger autophagy in vivo in the fat bodies of Drosophila L3 larvae [
150].Tetrandrine (9) is a bisbenzylisoquinoline alkaloid isolated from the stem of the creeper Stephania tetrandra S. Moore [
151]. The effect of tetrandrine (9) on human hepatocellular carcinoma (HCC) metastasis was evaluated both in vivo and in vitro. The obtained data showed that this compound stimulates autophagy in human liver cells, which modulates cancer cell metastasis by inhibiting WNT-CTNNB1/β-catenin pathway activity and decreasing MTA1 (metastasis-associated 1) expression [
152].Hernandezine (10) is an isoquinoline alkaloid isolated from Thalictrum glandulosissimum. Hernandezine (10) demonstrates potent cytotoxic effects toward different cancer cell types (A549, HeLa, PC3, MCF-7, Hep3B, HepG2 and H1299), while it shows low cytotoxicity toward normal liver hepatocytes. This compound activates AMPK directly and consequently induces ACD in apoptosis-resistant or drug-resistant cancer. Moreover, the resulting alkaloid-induced cellular toxicity is related to the upregulation of ATG7-dependent autophagy [
153].Bisleuconothine A (11) is a bisindole alkaloid isolated from Leuconotis griffithii. This compound increases LC3 lipidation in lung cancer A549 and in breast cancer MCF-7 cells, which might be due to the induction of autophagy via the AKT-MTOR signaling pathway. Bisleuconothine A (11) induces formation but inhibits the degradation of autophagosomes [
154].The utmost common cancer of the oral cavity is oral squamous cell carcinoma (OSCC), which is a serious global well-being issue. The major constituents of Murraya koenigii (L.) Sprengel are carbazole (12) alkaloids. The anti-cancer activities and mechanism of action of methanol and dichloromethane extract of the leaves of M. koenigii were investigated in OSCC cell line, CLS-354. The obtained results indicated that both isomahanine and carbazole alkaloids-mahanine cause increased accumulation of SSQTM1/p62, with a coordinated expression of LC3B-II and cleaved CASP3 (caspase 3), implying inhibition of autophagy flux. It was suggested that suppression of autophagy flux is related to carbazole-induced apoptosis [
155].Capsaicin (8-methyl-N-vanillyl-6-nonenamide) (13) is a functional element of chili peppers. Capsaicin (13) triggers mutated TP53 protein degradation through autophagy. Treatment with capsaicin (13) results in a reduction in mutated TP53 levels and reactivated WT TP53 protein in mutant TP53-carrying cells and the TP53 reactivation contributes to capsaicin-induced autophagy [
156].Alkaloids that are mentioned in this section are presented in
Figure 6. Moreover, the schematic role of these natural compounds on different proteins involved in the autophagy pathway is demonstrated in
Figure 7. 5.1.2. TerpenoidsIn 2005, the effect of some saponins isolated from soybeans was evaluated on the inhibition of colon tumor cell growth. According to the obtained results, a refined soybean B-group saponin extract caused the accumulation of human colon adenocarcinoma HCT-15 cells in the S phase along with a substantial increase in LC3-II levels and numerous autophagy vacuoles, showing autophagy induction [
157]. Neoalbaconol (14), a terpenoid isolated from the fungus Albatrellus confluens, targets PDPK1/PDK1 (3-phosphoinositide-dependent protein kinase 1) and inhibits the subsequent PI3K-AKT signaling in the nasopharyngeal carcinoma (NPC) nude mouse model, resulting in activation of autophagy [
158].Guttiferone K (15) is isolated from Garcinia yunnanensis Hu. This compound results in stress-induced cell death through an AKT-MTOR-dependent autophagy pathway in cervical cancer HeLa cells by MAPK/JNK activation and AKT-MTOR inhibition, causing LC3-II accumulation and SQSTM1 degradation [
159].Oridonin (16), an active diterpenoid isolated from Rabdosia rubescens [
160], induces autophagy via deactivation of p-AMPK and downregulation of SLC2A1/GLUT1 (solute carrier family 2 member 1) in the CRC cell line, SW480 [
161].Paclitaxel (17) is a taxane class diterpenoid, which was first discovered in the Pacific yew tree (Taxus brevifolia Nutt). Paclitaxel (17), with the trademark of Taxol, has been identified as a promising anti-cancer drug [
162]. The anti-cancer effects of paclitaxel (17) have been extensively studied [
163,
164]. The accumulation of ATG genes (ATG7, ATG12, ATG5), BECN1 and MAP1LC3B has been found in drug-resistant HeLa cells (HeLa-R) treated with paclitaxel (17). HeLa-R cells may undergo a shift in cellular metabolism from the tricarboxylic acid cycle to glycolysis; this activates HIF1A in HeLa-R cell lines. Suppression with koningic acid or glycolysis by 2-deoxy-D-glucose and downregulation of HIF1A by specific siRNA can reduce autophagy and increase the sensitivity of HeLa-R cells to paclitaxel (17) [
165]. In 2016, it was found that paclitaxel (17) activates BECN1 in 2 NSCLC lines, A549 and Calu-3 in a dose-dependent manner. The microRNA MIR216B levels are significantly downregulated in paclitaxel-treated cells. In fact, paclitaxel (17) modulates MIR216B, which targets BECN1 upregulation to increase autophagy in NSCLC [
166]. Research shows that paclitaxel (17) can suppress the proliferation of cervical cancer SiHa cells in a dose-related mode. However, following treatment with the long non-coding RNA RP11-381N20.2 substantial changes are found in the expression of key regulators of cell death pathways and autophagy is reduced significantly [
167].In 2011, it was proposed that gelomulide K (18), a diterpene, can induce PARP1 (poly(ADP-ribose) polymerase 1) hyperactivation, AIFM/AIF nuclear translocation and cytoprotective autophagy in cancer cells. Gelomulide K (18) can synergize with paclitaxel (17); together, it shows low toxicity in normal cells. This compound induces autophagy through an ROS-mediated downstream signaling pathway. Moreover, the significance of the 8,14-epoxy group in the structure of gelomulide K (18) was highlighted for induction of cell death and autophagy [
168].Tanshinone IIA (19), an abietane diterpenoid, isolated from Salvia miltiorrhizae, was reported to trigger autophagy in KBM-5 leukemia cells through AMPK activation and suppression of MTOR and RPS6KB/p70S6K. Moreover, it significantly increases the expression of LC3-II and activates the MAPK/ERK signaling pathway, such as RAF, MAPK/ERK and RPS6KA1/p90 RSK in a dose- and time-related mode [
169].A sesquiterpene lactone named helenalin (20) was reported to trigger autophagy via inhibition of RELA/NF-κB p65 expression in the RKO colon carcinoma, MCF-7 breast adenocarcinoma and A2780 human ovarian cancer cell lines. According to the acquired results, helenalin (20) treatment induces autophagy markers (MAP1LC3B and ATG12) and caspase activation, showing caspase activity is vital for ACD triggered by helenalin (20) [
170].Elevated MAPK/JNK signaling association with autophagy induction has been demonstrated [
171]. Ursolic acid (21), a natural triterpenoid, was reported to modulate autophagy by stimulating the accrual of LC3B-II and SQSTM1/p62 with the engagement of the MAPK/JNK pathway in apoptosis-resistant CRC cells [
172].The results from an experiment on HCT116 colon cancer cells showed that ginsenoside compound K (22) stimulates autophagy mediated by the ROS-JNK1 pathway. Ginsenoside compound K (22) therapy of tumor cells causes sensitizing TNFRSF10A/TRAIL/APO2L1 (TNF receptor superfamily member 10a) and TNFRSF10A-triggered apoptosis through autophagy-related and -independent (TP53-DDIT3 pathway) TNFRSF10B upregulation [
173].Frondoside A (23), extracted from the sea cucumber Cucumaria frondosa, is a triterpenoid saponin with a sugar-steroid structure [
174], which inhibits autophagy in human urothelial carcinoma cell lines. Accumulation of LC3-I/II and SQSTM1 and LC3-positive organelles (autophagosomes), time- and dose-dependently, indicates inhibition of autophagy [
175].Oblongifolin C (24) is a natural compound isolated from Garcinia yunnanensis Hu. It is an effective autophagy flux inhibitor. Contact with oblongifolin C (24) leads to an elevated quantity of autophagosomes and defective degradation of SQSTM1 [
176].The chemical structures of all autophagy-modulator terpenoids are mentioned in
Figure 8. The schematic role of the mentioned terpenoids on the proteins involved in the autophagy pathway is shown in
Figure 9. 5.1.3. Flavonoids and Phenolic CompoundsQuercetin (25), a flavonoid molecule discovered in vegetables, grains, leaves and fruits has exhibited anti-cancer effects on different cancer cells [
177,
178,
179]. Quercetin (25) causes preferential degradation of oncogenic RAS and leads to autophagy in HRAS/Ha-RAS-transformed human colon cells, DLD-1 and HT-29. Treatment with 3-methyladenine (3-MA), an autophagy inhibitor, inhibits quercetin-induced cell death. Moreover, the formation of EGFP-LC3 is detected in quercetin-treated cells. Moreover, zVAD-FMK, a general caspase inhibitor, fails to suppress vacuolization, demonstrating that the induced autophagy is caspase-independent [
180]. Moreover, quercetin (25) induces autophagy in GC cells. The results indicate the engagement of PI3K-AKT-MTOR and HIF1A signaling in quercetin-triggered autophagy development. In the presence of quercetin (25), formation of acidic vesicular organelles (AVOs), autophagic vacuoles, conversion of LC3-I to LC3-II, activation of autophagy genes and recruitment of LC3-II to phagophores are detected. Accordingly, quercetin (25) triggers autophagy progression in GC cells, such as AGS, BGC-823, SGC-7901 and MKN-28 [
181]. Quercetin (25) suppresses the proteasome function in breast cancer cells, resulting in blocking MTOR activity by a decrease in the MTOR substrates EIF4EBP1/4E-BP1 and RPS6KB/p70S6 kinase phosphorylation, consequently triggering autophagy [
182]. Emblica officinalis extract reduces cell proliferation in ovarian cancer OVCAR3 cells, dose- and time-dependently, by significantly increasing the expression of the autophagy proteins BECN1 and LC3B-II in vitro. Quercetin (25) as one of the components of the extract also increases the expression of these proteins and consequently reduces cell proliferation in ovarian cancer cells under in vitro conditions [
183].Apigenin (26), a common dietetic flavonoid available in vegetables and fruits, shows promising biological effects against different cancer cells [
184,
185,
186]. This compound initiates autophagy through the suppression of MTOR and RPS6KB/p70S6K of the JAK-STAT pathway in leukemia TF1 cells [
187].Genistein (27), a naturally derived isoflavonoid discovered in soy products [
188], can trigger cell death through autophagy in ovarian cancer cells via a caspase-independent pathway in a starvation-like signaling response. Treatment with 25–100 µM genistein (27) results in reduced AKT phosphorylation [
189]. In 2010, another investigation showed that this isoflavonoid as a tyrosine kinase inhibitor activates autophagy in ovarian cancer cells through the PDE4A/PDE4A4 (phosphodiesterase 4A) and SQSTM1 pathways. Genistein (27) inhibits both PRKC/PKC and MAPK/ERK inhibitors via suppression of PDE4A mass formation, which results in the induction of autophagy [
190].Salvigenin (28), a natural polyphenolic compound, increases the levels of autophagy factors, including the LC3-II:LC3-I ratio, ATG12 and ATG7 in human neuroblastoma SH-SY5Y cells [
191].Chrysin-organotin is a synthetic compound based on a natural flavonoid named chrysin (29). Research has shown that treatment with chrysin (29) and chrysin-organotin/Chry-Sn in breast cancer MCF-7 cells results in an enhancement of LC3-II levels, giving rise to the induction of autophagy in the treated cells [
192].Another natural flavonoid named baicalin (30) is present in several medicinal plants including Scutellaria baicalensis Georgi [
193]. Repolarization of tumor-associated macrophages towards the M1 phenotype characterizes the immunocompetent microenvironment in favor of tumor regression. Baicalin (30) treatment in hepatocellular carcinoma induces this repolarization, which is implemented by autophagy induction through activation of the RELB-p52 pathway and degradation of TRAF2 (TNF receptor associated factor 2) [
194].Juglanin (31) is a flavonol compound isolated from Polygonum aviculare, which triggers autophagy and apoptosis in breast cancer cells via MAPK/JNK activation and ROS production. A MAPK/JNK inhibitor attenuates juglanin (31) and significantly induces apoptosis and autophagy, whereas an ROS scavenger can reverse them [
195].Resveratrol (32) is a stilbenoid that is found in significant concentrations in berries, red wine, grapes and some other herbal sources [
196]. A connection between autophagy and resveratrol (32) has been shown in the treatment of several cancer cells. It has been demonstrated that this compound triggers autophagy in ovarian tumor cells. Morphology and ultra-structural changes and even a high amount of the anti-apoptotic proteins BCL2L1/Bcl-xL and BCL2 indicate that resveratrol (32) stimulates cell death in ovarian cancer A2780 and CaOV3 cells via a different mechanism, compared to apoptosis [
197]. Resveratrol (32) significantly suppresses breast cancer cell development in breast cancer stem-like cells separated from SUM159 and MCF-7 by repressing the WNT-CTNNB1/β-catenin signaling pathway. Moreover, LC3-II, BECN1 and ATG7 are significantly upregulated by resveratrol (32) in breast cancer cells in a dose-related mode [
198]. The resveratrol (32) impact was investigated in C33A (TP53 mutant), HeLa and CaLo (HPV18+), and CaSki and SiHa (HPV16+) cervical cancer cell lines. An elevated lysosomal permeability (autophagy) was observed in HeLa, C33A and CaLo cell lines in a concentration-independent manner [
199]. Resveratrol (32) also induces autophagy in melanoma B16 cells via ceramide accumulation and suppression of the AKT-MTOR pathway [
200]. Moreover, resveratrol (32) triggers autophagy-mediated cell death in prostate tumor cells (PC3 and DU-145) through regulation of a store-operated calcium entry (SOCE) mechanism, including downregulating STIM1 (stromal interaction molecule 1) expression, which triggers ER stress and therefore activates AMPK and inhibits the AKT-MTOR pathway [
201]. The ability of resveratrol (32) to trigger a starvation-like signaling reaction through decreasing phosphorylated AKT and MTOR to start autophagy was reported in ovarian tumor cells [
202]. However, the AKT-MTOR pathway is not engaged in resveratrol-triggered autophagy in resveratrol-treated U373 glioma cells. MAPK14/p38 or MAPK1/ERK2-MAPK3/ERK1 inhibitors decrease the autophagy pathway, suggesting that resveratrol-induced autophagy is positively controlled by MAPK14/p38 and the MAPK1-MAPK3 pathways [
203]. Resveratrol (32) was concluded to trigger ACD in chronic myelogenous leukemia cells through both AMPK activation and MAPK/JNK-mediated SQSTM1 overexpression [
204].Another study reported that resveratrol (32) activates a novel type of autophagy in breast tumor cells. Resveratrol (32) can inhibit the activation of AKT and MTOR in the MCF-7 cell line. Resveratrol (32)-induced ACD is impeded by CASP3 expression, except for its enzymatic function. Overexpression of BCL2 cannot reverse the non-canonical autophagy induced by resveratrol (32) in these breast tumor cells and resveratrol (32)-induced autophagy acts as a caspase-independent cell death mechanism in breast tumor cells [
205]. Moreover, in some studies, resveratrol (32) was reported to induce premature senescence in squamous cell carcinoma (SCC) A431, which is related to a blockage in autolysosome formation, as evaluated by the lack of colocalization of essential markers of lysosomes and autophagosomes LAMP2 (lysosomal associated membrane protein 2) and LC3, respectively. RICTOR (RPTOR independent companion of MTOR complex 2) is a crucial element of the MTORC2 complex. Resveratrol (32) is able to downregulate the amount of RICTOR, which in turn decreases RHOA GTPase activity, alters the actin cytoskeleton network, elevates the senescence-associated GLB1/β-gal function and attenuates the autophagy process, which may be the mechanism of tumor inhibition related to early senescence [
206].The phosphatidylinositol-3-phosphate (PtdIns3P) effectors, WIPI1 (WD repeat domain, phosphoinositide interacting 1) and WIPI2 (WD repeat domain, phosphoinositide interacting 2) act downstream during the start of autophagosome creation and following resveratrol (32) treatment. The involvement of WIPI1 with the phagophore membrane was confirmed by the localization of WIPI1 at the ER and the plasma membrane, upon triggering autophagy [
207]. An investigation was conducted to assess the autophagy-inducing properties of resveratrol (32) in four human tumor cell lines (G361, U2OS, MCF-7 and HeLa). The results indicate that WIPI1 is specifically bound to PtdIns3P and becomes a membrane-associated protein of the phagophore. Moreover, WIPI1 increases the LC3-II level, upon nutrient starvation and functions upstream of both ATG5 and ATG7. It was suggested that resveratrol (32)-mediated autophagy occurs through the non-canonical pathway and is BECN1-independent but ATG7- and ATG5-dependent [
208]. Resveratrol (32) was reported to increase the protein expression of ATG5, ATG7, ATG9 and ATG12 in a human hepatitis C-triggered Huh-7 hepatoma cell line inducing ACD [
209].Glioblastoma is one of the increasingly prevalent main brain cancers [
210]. Resveratrol (32) induces the formation of autophagosomes in three human glioblastoma cell lines (U-251, U-87 MG and U-138 MG) via upregulation of autophagy proteins ATG5, BECN1 and LC3-II [
211]. SIRT1 (sirtuin 1) is a NAD-dependent deacetylase that has a key function in regulating autophagy; it forms a complex with some autophagy machinery components ATG5, ATG7 and Atg8-family proteins and directly deacetylates these components in a NAD-dependent fashion [
212]. Resveratrol (32) is able to regulate SIRT1 and PARP and consequently regulates induced autophagy in lung cells with cigarette smoke-mediated oxidative stress [
213]. SERPINB4/SCCA1 (serpin family B member 4), an endogenous CTSL (cathepsin L) inhibitor, is widely expressed in various uterine cervical cells. Autophagy and the CTSL-SERPINB4/SCCA1 lysosomal pathway are engaged in resveratrol-induced cytotoxicity in cervical tumor cells. Moreover, resveratrol (32) increases the level of LC3-II and autophagosomes in these cells [
214].Curcumin (33) is a natural curcuminoid with high anti-cancer efficacy. Curcuminoids are the major polyphenolic-derived compounds found in the rhizome of many Curcuma species, which have been applied for centuries in conventional Indian and Chinese medicine. The primary study on the anti-cancer activities of curcumin (33) was published in 1987 [
215]. The antineoplastic activity of curcumin (33) has been comprehensively investigated in several cancers, including melanoma, breast [
216], colon and prostate cancers [
217]. There are numerous studies demonstrating the beneficial effect of curcumin (33) in modulating autophagy. Curcumin (33) suppresses the AKT-MTOR-RPS6KB/p70S6K pathway and stimulates the MAPK1-MAPK3 pathway, thereby inducing autophagy [
218]. Curcumin (33) exerts anti-proliferative effects on two human malignant glioma cell lines (U373-MG and U87-MG) by arresting the cell cycle in the G2/M phase and promoting autophagy by inhibiting the AKT-MTOR-RPS6KB/p70S6K pathway and stimulating the MAPK1-MAPK3 pathway [
219]. In 2011, the participation of ROS in curcumin-triggered ACD was investigated. The data indicate that curcumin (33) leads to ROS production, which results in ACD in human colon cancer HCT116 cells. However, curcumin-triggered autophagy is not implicated in the ROS-dependent stimulation of MAPK1-MAPK3 and the MAPK14/p38 signaling pathway. Thus, a combination of curcumin (33) and agents that result in ROS production might enhance the cytotoxic effect of this compound [
220].Additionally, this compound exhibits anti-cancer activity against OSCC through induction of autophagy, which was confirmed by detecting a significant increase in LC3-I conversion to LC3-II in curcumin-treated cells. In this research, a substantial level of AVOs can be identified in spite of a brief duration of exposure (6 h) to curcumin (33) [
221]. This compound also induces autophagy in the human lung adenocarcinoma cell line, A549, as confirmed by the analysis of LC3 and SQSTM1. In addition, it enhances the phosphorylation of AMPK and ACAC (acetyl-CoA carboxylase) in A549 cells, although it can also cause blocking and disruption in the AMPK signaling pathway [
222]. Curcumin (33) induces autophagy in SK-UT-1 and human uterine leiomyosarcoma SKN cell lines through stimulation of the MAPK1-MAPK3 pathway; accordingly, suppression of the MAPK1-MAPK3 pathway decreases curcumin-induced autophagy [
223].Lysosomes have a vital function in the maturation and degradation of autophagosomes. A major molecular mechanism in the control of lysosomal action involves nuclear TFEB (transcription factor EB), which is the immediate downstream focus of MTOR. TFEB activity is regulated by the MAPK1/ERK2 pathway [
224,
225]. Data obtained from one study revealed that curcumin (33) triggers autophagy in human colon cancer HCT116 cells by activation of lysosomal function via suppression of the AKT-MTOR signaling pathway and induces TFEB transcriptional activity by direct binding to TFEB [
226]. The effect of curcumin (33) was investigated on H2O2-triggered apoptosis and autophagy in EA.hy926 cells. In this case, curcumin (33) induces autophagy and LC3-II expression and suppresses the phosphorylation of AKT-MTOR, indicating its protective effect against oxidative stress damage by triggering autophagy via AKT-MTOR pathway inhibition [
227]. Curcumin (33) also triggers autophagy and suppresses proliferation by downregulating the AKT-MTOR signaling pathway in human melanoma A375 and C8161 cell lines [
228]. Curcumin (33) potentiates autophagy in pancreatic cancer PANC1 and BxPC3 cells, as confirmed by the promotion of autophagosome formation, upregulation of LC3-II and downregulation of the MTOR protein expression [
229].Finally, inhibition of apoptosis following curcumin (33) treatment in osteosarcoma MG63 cells increases autophagy due to upregulation of the MAPK/JNK signaling pathway [
171]. In addition, the impact of curcumin (33) on the proliferation, apoptosis and autophagy of GC cells (SGC-7901 and BGC-823) shows that this compound inhibits PI3K via downregulating the PI3K-AKT-MTOR pathway and activates the TP53 signaling pathway by upregulating TP53 and CDKN1A/p21, both of which regulate autophagy [
230].Some derivatives of curcumin (33) also possess anti-cancer activities. A derivative of curcumin named bis-dihydroxy-curcumin (36) increases autophagosome formation and autophagy flux in human colon cancer HCT116 cells. An enhancement of ER stress and poly-ubiquitinated proteins is detected upstream of autophagy initiation and results in cell death. Thus, this compound induces autophagy through an ER-stress pathway [
231]. Moreover, tetrahydrocurcumin (37) is one of the primary catabolic products of curcumin (33). This compound suppresses the PI3K-AKT-MTOR-RPS6KB/p70S6K, GSK3B
(glycogen synthase kinase 3 beta) and MAPK14/p38 pathways and activates the MAPK1-MAPK3, MAPK8/JNK1-MAPK9/JNK2 pathways. These changes cause the initiation of autophagy in human leukemia HL-60 cells [
232].Honokiol (34), a biphenolic compound isolated from Magnolia officinalis, acts as a potent inhibitor of melanoma B16-F10 cells by targeting melanoma stem cell signaling pathways by either suppressing NOTCH1 (notch receptor 1) signaling or increasing autophagosome formation, suggesting that honokiol (34) can also induce autophagy in melanoma cells via attenuating AKT-MTOR [
233].Magnolol (35), a lignin compound, induces autophagy via the PI3K-AKT signaling pathway in human GC cells SGC-7901 at elevated amounts, but it is not engaged in cell death [
234].Rottlerin (38) is a polyphenol isolated from Mallotus phillippinensis (the monkey-faced tree, Kamala) [
235]. Rottlerin (38) triggers autophagy in human fibrosarcoma HT1080 cells through a PRKCD/PKCδ-independent pathway. The LC3-II protein level also increases following the administration of rottlerin (38); early induction of autophagy acts as a survival mechanism versus late apoptosis [
236]. Another pathway through which autophagy is regulated via rottlerin (38) involves PRKCD-TGM2 (transglutaminase 2) and downregulation of PRKCD results in ACD [
237]. Moreover, this phytochemical inhibits MTORC1 signaling through its negative controller TSC2 in breast tumor cells in a high nutritional environment. Rottlerin (38) causes the assembly of EGFP-LC3-II and free EGFP, along with proteolytic fragments of EGFP [
238]. Moreover, rottlerin (38) induces ACD in apoptosis-resistant breast cancer MCF-7 cells, as confirmed by an elevated LC3-II:LC3-I ratio and degraded SQSTM1 protein. The autophagic death appears to be triggered by non-canonical signaling cascades because it is independent of BCL2, BECN1, AKT and MAPK/ERK pathways [
239].Another investigation using breast cancer stem cells revealed that rottlerin (38) treatment increases the expression of ATG12, BECN1 and LC3 proteins. Rottlerin (38)-triggered autophagy is mediated through stimulation of the AMPK pathway [
240]. Rottlerin (38) treatment results in BCL2- and BECN1-independent autophagic death in apoptosis-resistant breast cancer MCF-7 cells. Interestingly, it was reported that rottlerin (38)-triggered autophagy is mediated by suppression of MTORC1; however, any role for AMPK was excluded and it was suggested that rottlerin (38) interacts directly with MTORC1 and induces autophagy through a novel AMPK- and MTORC1 phosphorylation-independent mechanism [
241]. Rottlerin (38) triggers autophagy via the PI3K-AKT-MTOR signaling pathway in prostate tumor stem cells; as a result, the formation of autophagosomes, an increase in LC3-II and induction of BECN1, ATG7, ATG5 and ATG12 are observed [
242].Gossypol (39) is a natural phenol isolated from the cotton plant (genus Gossypium) [
243]. An enhancement in LC3-II levels and autophagosome numbers in mutant TP53 pancreatic adenocarcinoma PANC-1 cells are observed in the presence of a combination of gossypol (39) and BRD4770, an inhibitor of histone methyltransferase EHMT2/G9a (euchromatic histone lysine methyltransferase 2). This combination acts in a BNIP3 (BCL2 interacting protein 3)-dependent manner. It seemed that gossypol (39) enhances BRD4770 cytotoxicity significantly in TP53-mutant cells and induces autophagy-related cell death. The upregulation of BNIP3 upon EHMT2 inactivation might be responsible for the synergistic ACD effect. Thus, EHMT2 inhibition may solve drug resistance issues in certain cancer cells [
244].Salvianolic Acid B (40) is a phenolic acid isolated from the dried root and rhizome of Salvia miltiorrhiza that triggers autophagy via the AKT-MTOR pathway suppression in CRC cell lines, HCT116 and HT-29 [
245].Isoliquiritigenin (41), a natural flavonoid derived from the root of licorice (Glycyrrhiza uralensis), triggers autophagy in drug-resistant breast tumor MCF-7 cells through MIR25 inhibition, thus functioning as a regulator of chemoresistance-associated autophagy by promptly elevating ULK1 expression [
246].Summaries of the flavonoids and phenolic compounds that act as autophagy modulators are shown in
Figure 10 and
Figure 11. The schematic role of these compounds on the proteins involved in the autophagy pathway is shown in
Figure 12.

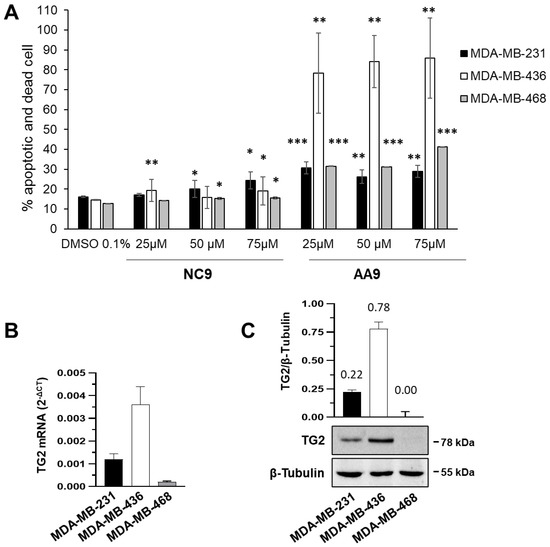
留言 (0)