2.1. Structural Analyses of the Pol I Enzyme in Saccharomyces cerevisiaeStructural studies of Pol I have detailed the enzyme and the preinitiation complex in the various steps in the transcription cycle, namely, preinitiation, initiation, and elongation ([
38,
39,
40,
41,
42,
43,
44,
45,
46,
47], reviewed in [
3]). The majority of these studies have been conducted in the yeast model Saccharomyces cerevisiae. The nomenclature for yeast and human subunits is different and has evolved over time. We refer the reader to a recent review for the naming convention [
3] and identify here the key subunits with their current species-specific names. In yeast, Pol I is composed of 14 subunits, with a 10-subunit core that resembles a crab claw [
38,
39]. The two largest subunits, A190 (human RPA1) and A135 (human RPA2) form the catalytic core and the DNA binding cleft [
38,
39]. The core encloses a central cleft, where the template DNA binds and the nascent rRNA strand is synthesized. The cleft has two channels, for the entry of substrate nucleoside triphosphates (NTPs) and exit of the RNA product [
38,
39].In addition to the catalytic subunits, the 10-subunit core is formed by several subunits shared with Pol II and Pol III (ABC27, ABC23, ABC14.5, ABC10α, ABC10β) and with Pol III (AC40, AC19), which provide mostly structural support for the core assembly [reviewed in [
3]. The 10-subunit core of the Pol I complex also contains A12.2 (human RPA12), a subunit composed of two Zn-binding β-ribbon domains. The N-terminal ribbon connects A12.2 with A190, A135, and a heterodimer of A49-A34.5 (human RPA49 and RPA34, respectively) subunits [
38,
39]. The C-terminal ribbon extends into the active site and interacts with the NTP entry channel and is involved in RNA cleavage during backtracking/proofreading and termination [
38,
39]. Importantly, the A12.2 C-terminus is only positioned into the active site when Pol I is not in the elongation phase and is displaced during elongation to allow NTP addition [
38,
39,
40,
41].Pol I contains four peripheral subunits unique to this enzyme: A43 (human RPA43), A14, A49, and A34.5. The A43-A14 heterodimer forms the stalk, a structure that protrudes from the core. Preinitiation factor Rrn3 (human RRN3) binds to the A43-A14 stalk and recruits the Rrn3-Pol I complex to the rDNA promoter [
40,
41,
44,
45,
46,
47]. A34.5-A49 are homologous to the Pol II TFIIE/F heterodimer, but in contrast to their auxiliary arrangement in Pol II, are integral to the Pol I complex [
48]. They associate with the Pol I lobe (A135) and cavity and A12.2 N-terminal domain. A49 has a tandem winged helix (tWH) domain that forms a bridge over the Pol I DNA binding cleft, which is mobile and displaced to allow promoter DNA binding. The tWH domain affects the enzyme processivity, while the C-terminus is required for DNA binding in vitro and orients towards the upstream promoter [
40,
41,
43]. A49 helps recruit Rrn3-bound Pol I to the promoter and aids in the release of Rrn3 after promoter clearance [
41]. Furthermore, the A49-A34.5 heterodimer binds to and stabilizes A12.2 and stimulates its RNA cleavage activity [
38,
39]. In addition to transcription initiation, the A49-A34.5 heterodimer has been implicated in transcription elongation [
49]. 2.3. Key Steps in the Pol I Transcription CyclePol I is a highly efficient RNA polymerase that has evolved for the transcription of its long 13 kb transcript. The structure of Pol I, with its in-built RNA cleavage ability and incorporation of subunits homologous to transcription factors, enables it to quickly and efficiently synthesize the enormous amount of rRNA required for ribosome biogenesis [
52,
53]. This is facilitated by its high initiation rate and elongation speed [
54]. Many Pol I complexes are densely packed on the rDNA, with an elongation complex every 140 bp on the actively transcribed genes in yeast [
55]. Transcription occurs through tightly regulated cycles [
9,
31,
54].
Initiation. The 140-160-base pair rRNA gene promoter contains two key elements, a core element and the UCE [
7,
31]. Binding of the mammalian upstream binding factor (UBF) to the UCE and core element mimics a nucleosomal fold. UBF recruits and activates selectivity factor 1 (SL-1) [
7,
56]. While UBF is present throughout the bodies of actively transcribed coding genes, SL-1 is only present on the gene promoters. SL-1 consists of a TATA-binding protein (TBP) and five additional factors: TAFI110, TAFI48, TAFI63, TAFI12, and TAFI41 [
7,
31]. RRN3 interacts tightly with the RPA43 stalk of Pol I [
51]. Similarly, the association of RRN3 with RPA43 prevents the enzyme dimerization and maintains Pol I in its monomeric form [
44,
45,
46,
47,
51]. SL-1 subunits TAFI63 and TAFI110 interact with RRN3 on the RRN3-Pol I complex and recruit Pol I to the promoter. Together, these proteins form the PIC. This process is relatively conserved in yeast, as yeast has Rrn3, TBP, and a core complex homologous to SL-1 [
44,
45,
46,
47]. However, yeast also have an upstream activating factor (UAF), which aids in initiation but has no mammalian homologue [
54].The successful assembly of the PIC bends the promoter to an angle, remarkably different from that of the Pol II promoter, favorable for transcription initiation. Pol I stutters on the first few nucleotides, during which the DNA duplex melts to favor the formation of the transcription bubble and promoter escape. This process is further assisted by A49 and A12.2, which support the binding of Pol I to the promoter and the formation of the open elongation complex, respectively [
40,
43]. Pol I re-configures from its open complex (bound to DNA) into its elongation complex (synthesizing RNA), followed by dissociation of RRN3 [
40,
43]. With these features in place, Pol I is positioned for not only the high initiation rates but also the rapid transition into the elongation phase.
Elongation. In the active site, two magnesium cations in the catalytic aspartate triad coordinate an NTP condensation reaction. During this reaction, one nucleoside monophosphate (NMP) is added to the nascent RNA strand as pyrophosphate is released. Pol I then translocates down the DNA template by one nucleotide [
54]. This translocation is aided by other elements in the active site, such as the bridge helix, rudder, fork loop(s), trigger loop, and wall [
3]. As each NMP is added to the 3′ end of the growing RNA chain, one NMP from the 5′ end goes into the RNA exit tunnel [
57]. Pol I nucleotide addition rates are faster than those of Pol II, but this comes with the cost of a less stable enzyme and higher error rates [
58]. The mechanic nucleotide addition has been compared to a “ratchet” that drives the forward reaction. However, the Pol I enzyme is not indifferent to its substrate—the sequence context of the substrate, especially the GC-richness, and the co-transcriptional folding of the RNA product both affect the enzyme elongation rate [
59,
60,
61,
62].
Pausing and Backtracking. Given the ratchet motion during elongation, the enzyme is prone to backtracking. Furthermore, if the polymerase encounters an obstacle, such as DNA damage or an incorrect nucleotide, it will pause. The polymerase will then backtrack. Since the “gating tyrosine” in the active site remains “open”, the polymerase can backtrack as much as it needs to. The C-terminus of A12.2 inserts into the active site and cleaves the RNA [
63,
64]. This cleavage activity provides more efficient backtrack recovery for Pol I compared to Pol II. While the C-terminus of A12.2 has the cleavage activity, it is supported in this task by the N-terminal domains, as well as by the A34.5–A49 heterodimer [
65]. Inefficient RNA cleavage further leads to proofreading errors [
65], and the fidelity of Pol I transcription decreases by 10-fold in the absence of A12.2 [
66].
Termination. Transcription termination elements are positioned on two separate sites on the rDNA gene repeat: at the 3′ end of the transcribed region and upstream of the transcription start site. Transcription termination factor I (TTF-I) binds to the termination element at the 3′ end of the transcribed region, bends the DNA, and triggers Pol I to pause [
7]. However, given the RNA cleavage activity of the enzyme and co-transcriptional processing, TTF-1 is not essential for the cleavage step but aids in preventing transcription from continuing into the IGS. A12.2 is required for the Pol I release from the DNA template [
54]. Given that the promoter and termination sites are closely spatially positioned, the Pol I-RRN3 complex can be recruited back to the same or another UBF and SL-1-bound rDNA promoter to re-engage with the transcription cycle [
67,
68].
Pol I Complex Stability. Pol I processivity is facilitated by enzyme stability. However, the mechanistic understanding of which factors govern mammalian Pol I assembly, stability, and localization is incomplete. In yeast, the interaction between A190 and A135 is stable [
69], and in mammals, RPA1 and RPA2 have long half-lives (>20 h). We previously showed that the stability of RPA1 (yeast A190) is dependent on its binding partner, RPA2 (yeast A135), as silencing of RPA2 reduced RPA1 protein expression and caused its nucleoplasmic translocation [
70].In yeast, A49 stabilizes the expression of A34.5 and is mediated by a protease-sensitive linker domain. Conversely, depletion of A34.5 also destabilizes A49, suggesting that this complex is inherently unstable if either subunit is missing [
49]. UV light causes bulky pyrimidine dimer lesions that stall transcription elongation complexes. At cryo-EM resolution these lesions cause RPA1 structural rearrangements that block the enzyme translocation step [
71]. In contrast, the A49 domains remain in an open complex configuration, suggesting that the polymerase keeps scanning for initiation even when facing these extensive blocks. A yet unanswered question is whether the A34.5-A49 dimer dissociates from the core complex during the transcription cycle or when facing elongation blocks [
72]. Furthermore, kinetic studies on the nucleotide addition rate have suggested that A12.2 acts as an intrinsic destabilizer of Pol I elongation complex in vitro [
73].External factors, such as zinc availability and temperature, also mediate Pol I stability. Zinc depletion has been shown to induce vacuolar proteolysis of Pol I in yeast [
74]. Cold temperatures induce the ubiquitination and destabilization of Rpa190, the yeast homolog of mammalian RPA1 [
75]. Since the deubiquitylating enzyme Ubp10 affects Rpa190 stability [
75], it is possible that Rpa190 is marked for degradation through the ubiquitin proteasome system. This possibility is intriguing, since the largest subunits of both Pol II and Pol III undergo proteasome-mediated degradation when facing transcription challenges. This is discussed in more detail below [
76,
77,
78,
79,
80,
81].

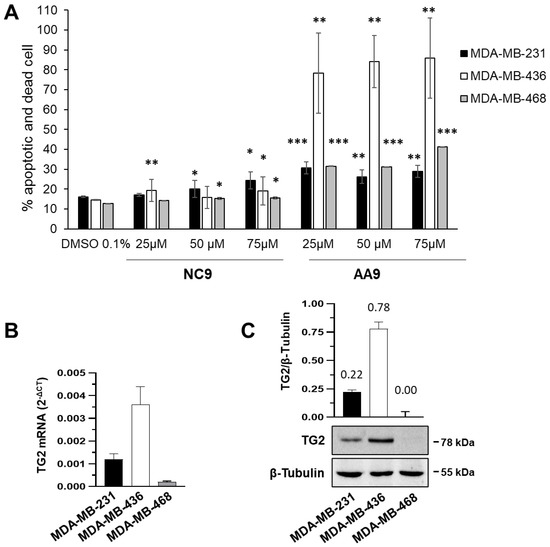
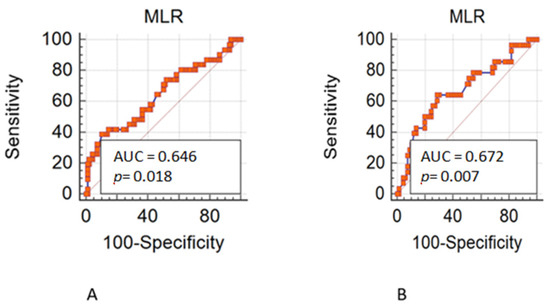
留言 (0)