
記住我

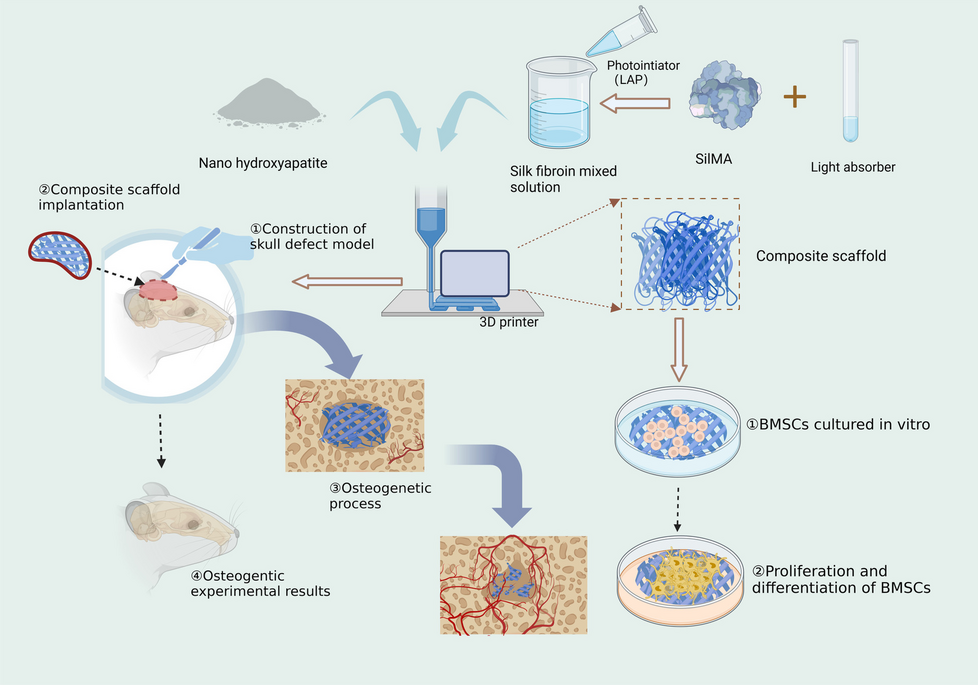
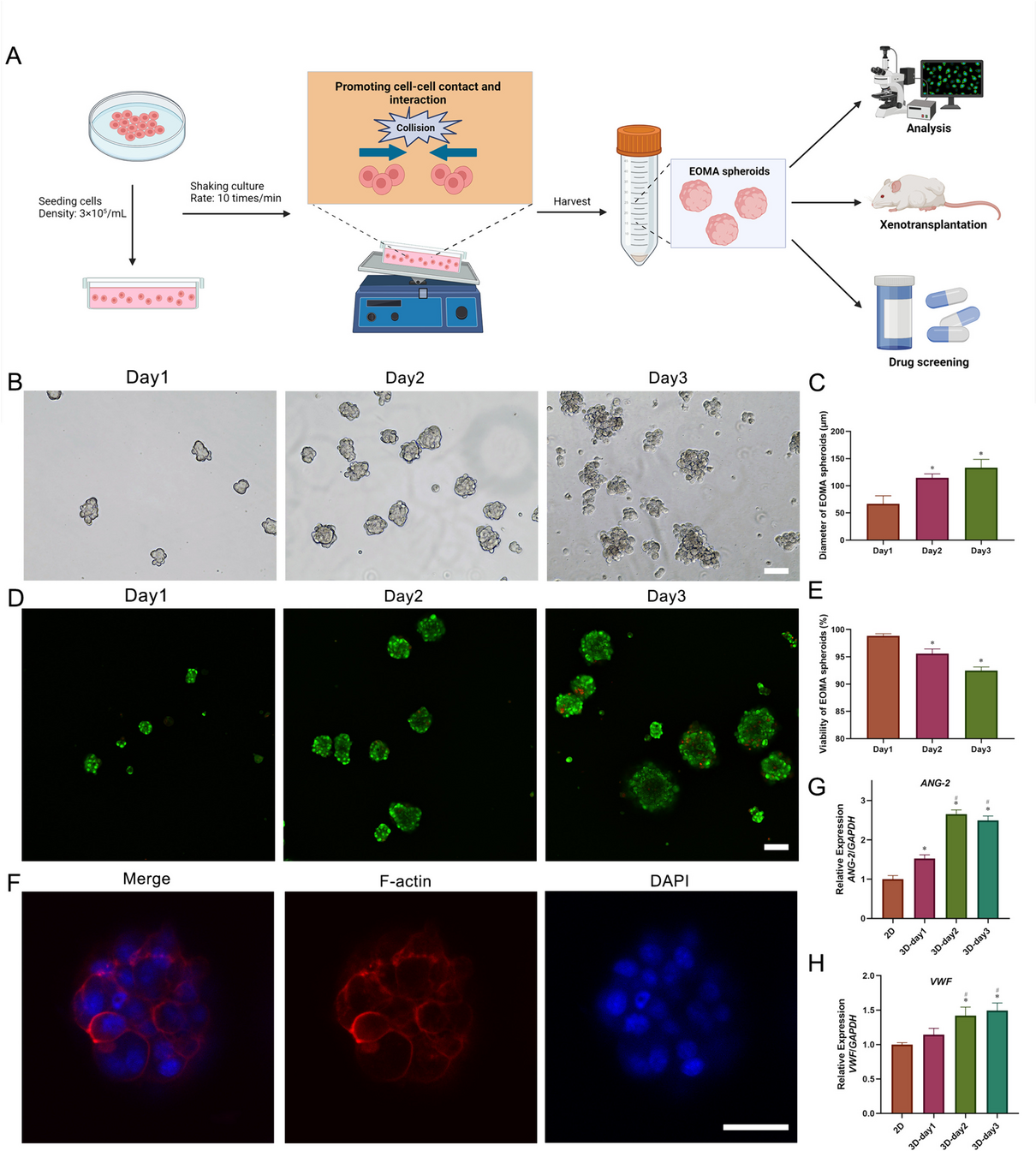
For the first experiment, the complete Poolman medium was divided into five component groups: nucleotides, trace elements, salts, amino acids and vitamins (Table 1). While the other component groups originated from the medium published by Poolman et al. [30], the component group of salts consist of (NH4)2SO4, KH2PO4 and MgSO4∙7H2O. These components are also included in the Gluconobacter complex medium and supply the organism with phosphate, nitrogen and magnesium. Therefore, they are expected to be essential for the media investigated in this work.
The growth of G. oxydans fdh in the complete Poolman medium was compared to growth in the established Gluconobacter complex medium with 60 g/L fructose. To gain a first impression of which component groups are essential for the growth of G. oxydans fdh, five additional cultivations in the complete Poolman medium, each lacking one of the five component groups, were conducted in parallel (Fig. 1).
G. oxydans fdh showed the typical OTR course in the Gluconobacter complex medium [14, 37]. The OTR increased exponentially up to a peak at approx. 32 mmol/L/h after 16 hours, followed by a sharp decline to 5 mmol/L/h, before the OTR decreased slowly over the following hours. The cultivation in complete Poolman medium showed a comparable OTR course, with a shifted OTR maximum of 36 mmol/L/h after 14 hours. Consequently, the growth of G. oxydans fdh is not limited, when cultivated in the rich, synthetic medium, but slightly improved, compared to the cultivation in Gluconobacter complex medium. Cultivations in Poolman medium without salts, amino acids, vitamins and trace elements showed significantly reduced OTRs. In contrast, the cultivation in the Poolman medium without nucleotides showed a similar OTR course as observed for Gluconobacter complex medium. In conclusion, supplementation of nucleotides improves growth in G. oxydans fdh, yet it is not essential. Since pathways for the synthesis of all nucleotides are included in the genome of G. oxydans [18] and nucleotides are not commonly supplemented in synthetic media, the entire component group will not be included in the final minimal medium.
An increase in OTR up to 15 mmol/L/h after 18 hours can be observed during the cultivation without trace elements. Trace elements are essential for microbial growth and should, therefore, be supplemented in synthetic media [38]. They are commonly included in the form of trace element solutions, without detailed consideration of the single components [39,40,41]. The conversion of fructose to 5KF is stoichiometrically linked to oxygen consumption and, thus, visible in the OTR [14]. In the experiment shown in Fig. 1, respirational activity can still be observed, which is due to the transfer of small amounts of trace elements from the pre-culture and the highly active FDH [14, 15]. However, the reduced concentration of trace elements still impaired growth. Consequently, the trace element solutions of the complete Poolman medium remained unchanged and were used for further experiments.
Fig. 1
Cultivation of G. oxydans 621H ΔhsdR pBBR1p264-FDH-ST in a μRAMOS device with 60 g/L fructose and different media. Depicted is the oxygen transfer rate (OTR). Cultivations were performed at 30 °C, 1000 rpm, VL = 500 μL in a 48-well round well microtitre plate at a shaking diameter of 3 mm, initial pH value: 6. Media: Complete Poolman medium as reference (black), Gluconobacter complex medium containing 5 g/L yeast extract, 2.5 g/L MgSO4 ∙ 7H2O, 1 g/L (NH4)2SO4 and 1 g/L KH2PO4 (dark blue), Poolman without salts (light blue), without amino acids (green), without vitamins (orange), without nucleotides (pink), without trace elements (purple). Mean values of at least 4 replicates are shown. The shadows around the curves indicate the standard deviation. For clarity, only every fifth measuring point is marked as a symbol
The cultivations lacking salts, amino acids or vitamins showed only slight increases in OTR and significantly longer cultivation times. Hence, all component groups except nucleotides include at least one component crucial for G. oxydans fdh.
Nitrogen, phosphate and magnesium have important functions in an organism’s metabolism and are included in most synthetic media [39, 40]. Nitrogen is the fourth most common element in the biomass of G. oxydans [42] and is essential for its optimal growth [43, 44]. Phosphate plays a crucial role in energy metabolism [45] and the pentose phosphate pathway [46], which is of particular importance for oxidising sugars in Gluconobacter strains [22]. Magnesium is important for cell proliferation in most organisms by stimulating DNA and protein synthesis [47, 48]. The influence of different nitrogen sources will be further discussed later in this work. KH2PO4 and MgSO4∙7H2O will be used in unchanged concentration in the complete Poolman medium.
The complete Poolman medium contains 18 different amino acids as well as 12 vitamins. Since the cultivations using the Poolman medium without amino acids or vitamins showed very limited growth, at least one amino acid and vitamin are essential for the growth of G. oxydans fdh. As a result, five amino acids groups and four vitamin groups (Table 2) were established, according to the classification by Müller et al. [29]. It is based on metabolic pathways described by Akashi and Gojobori [29, 42]. Single amino acids and vitamins were further investigated in the following.
Table 2 Group of amino acids and vitaminsInvestigation of the influence of amino acids on the growth of G. oxydans fdhThe influence of amino acids on the growth and metabolism of Gluconobacter species has been intensively studied for decades. Since 1953, supplementation of amino acids has been reported to support growth in various Gluconobacter species [23, 25, 43]. Sainz et al. [28] showed that the optimal composition and concentration of amino acids deviate between different Gluconobacter species. Gosselé et al. [24] stated that no single amino acid is essential for the growth of most Gluconobacter strains, as growth was not decreased by the omission of single amino acids in their investigations. Consequently, a detailed study of the influence of amino acids on the growth of G. oxydans fdh in the Poolman medium was carried out in this work. In a series of experiments, the five amino acid sub-groups were individually omitted from the complete Poolman medium. When an influence of the omission on growth was observed, the single amino acids of the investigated group were added again. The resulting cultivations were compared to the cultivation without the concerning sub-group, to evaluate the influence of the single components. The composition of the amino acid sub-groups can be found in Table 2.
The results for groups 1–3 are displayed in Fig. 2, as omitting groups 4 and 5 did not negatively influence the growth of G. oxydans fdh (Fig. S1). Omitting group 1 from the complete Poolman medium led to a slightly delayed increase in OTR and a decreased peak height, compared to the complete Poolman medium.
Fig. 2
Investigation of auxotrophic deficiencies of G. oxydans 621H ΔhsdR pBBR1p264-FDH-ST in Poolman medium regarding the amino acids of group 1 (Asp, Ile, Met, Thr), group 2 (Arg, Pro, His, Glu) and group 3 (Cys, Gly, Ser). Depicted is the oxygen transfer rate (OTR). Cultivation of G. oxydans 621H ΔhsdR pBBR1p264-FDH-ST in a μRAMOS device with 60 g/L fructose, lacking amino acids from group 1 (aspartate, isoleucine, methionine, threonine), group 2 (arginine, glutamine, histidine, proline) and group 3 (cysteine, serine, glycine). Three independent cultivations (A, B and C) were performed at 30 °C, 1000 rpm, VL = 500 μL in a 48-well round well microtitre plate at a shaking diameter of 3 mm, initial pH value: 6. Media: Complete Poolman medium as reference (black) and A without group 1 (light blue), without group 1 with aspartate (purple), without group 1 with isoleucine (green), without group 1 with methionine (orange), without group 1 with threonine (pink), B without group 2 (pink) and without group 2 with glutamate (purple), C without group 3 (green) and without group 3 with glycine (orange). Mean values of at least 6 replicates are shown. The shadows around the curves indicate the standard deviation. For clarity, only every fifth measuring point is marked as a symbol
Figure 2 A). The addition of aspartate or methionine showed no positive influence on the respiration of G. oxydans fdh. Adding isoleucine or threonine, however, reduced the time until the OTR peak is reached by 2 hours. This effect was slightly stronger for the addition of isoleucine. Since respiration remains similar to the complete Poolman medium, it can be concluded that no amino acid in group 1 is absolutely essential for the growth of G. oxydans fdh. This is not surprising, as the components of group 1 did not receive much attention in research on Gluconobacter strains so far [21, 22]. Aspartate has been reported to be an important nutrient for acetic acid bacteria [44], while some Gluconobacter strains are able to deaminate threonine [24]. However, since isoleucine supplementation led to accelerated growth, isoleucine is included in the medium for further experiments.
Leaving out group 2 from the Complete Poolman medium reduced the respirational activity significantly (Fig. 2 B). The maximal OTR of 20 mmol/L/h was reached after 17 hours, compared to the OTR of the complete Poolman medium, which increased up to 45 mmol/L/h after 13 hours. The addition of glutamate increased the OTR peak height and reduced the time of the OTR peak to 32 mmol/L/h after 16 hours. Especially glutamate has been found to improve the growth of different acetic acid bacteria [22, 45], while Sainz et al. [28] described histidine as a sufficient sole nitrogen source for G. oxydans 621H. Like all amino acids, glutamate can be synthesised de novo by G. oxydans [18]. In the biosynthesis of other amino acids, it is an important donor of amino groups as well as a precursor. Moreover, it acts as an acceptor molecule for inorganic nitrogen in the ammonium assimilation of the cell [46]. In conclusion, glutamate is crucial for efficient growth and is, thus, included in different media for the cultivation of G. oxydans [22, 23, 47]. It is not surprising that supplementation of glutamate improves the cultivation of G. oxydans fdh, as evidenced by the observed increase in respirational activity (Fig. 2 B). Adding the other single amino acids of group 2 led to a similar effect (Fig. S1A). Glutamate, however, improved the cultivation the most and is, therefore, chosen to remain in the medium, to reach growth similar to the Gluconobacter complex medium with as few components as possible.
The cultivation without the addition of group 3 showed a significantly delayed increase in OTR, compared to the complete Poolman medium, reaching 30 mmol/L/h after 26 hours (Fig. 2 C). When glycine was added, the OTR progressed nearly identical to the complete medium with only a slight delay in reaching the maximum OTR. An almost identical observation was made for serine supplementation (Fig. S1B). Wethmar et al. [21] reported that serine was one of two amino acids required for the growth of G. oxydans in their work. In contrast, Tachiki et al. [48] described that glycine inhibited the glutamate-glutamine metabolism in G. suboxydans. According to the genome sequence of G. oxydans by Prust et al. [18], serine acts as a precursor for glycine production. Yoshitake et al. [49] described the microbial conversion of glycine to serine. In this context, the nearly identical results of serine and glycine addition to the medium are very plausible, and one of both components should remain in the medium. Since the cultivation with glycine showed a slightly shorter cultivation time (Fig. S1B), glycine was chosen as the remaining media component of group 3 for further experiments.
The amino acids in group 4 have been reported to play a role in the activity of single enzymes of G. oxydans [50, 51], while valine from group 5 has been reported to have an inhibitory effect on the growth of Acetobacter suboxydans [52]. Because of their limited influence on the growth of G. oxydans fdh, groups 4 and 5 were decided to be discarded from the medium (Fig. S1C). After investigation of all amino acids, only isoleucine, glutamate and glycine remain in the medium for further experiments. Since the nitrogen content of the media is strongly affected by the omission of the majority of amino acids, the influence of different nitrogen sources is investigated in the next step.
Investigation of nitrogen supply for the cultivation of G. oxydans fdhAfter omitting the majority of amino acids from the complete Poolman medium, only 37.8% of the original elemental nitrogen is left. Therefore, it has to be investigated, if the nitrogen requirements of G. oxydans fdh are provided in the reduced version of the medium. For that purpose, five cultivations were conducted with different concentrations of (NH4)2SO4 and amino acids to investigate the changes in nitrogen concentration and nitrogen source (Fig. 3). As a reference cultivation, the complete Poolman medium with a concentration of 912 mgN/L was used. The second cultivation was conducted without supplementation of (NH4)2SO4 with all amino acids in the original concentration (700 mgN/L). In the third cultivation, (NH4)2SO4 and the three crucial amino acids isoleucine, glutamate and glycine were supplemented in the original concentrations (345 mgN/L). In cultivation four, a 1.5-fold (NH4)2SO4 concentration was used, while the glutamate concentration was tripled for the fifth cultivation (440 mgN/L each).
Fig. 3
Cultivation of G. oxydans 621H ΔhsdR pBBR1p264-FDH-ST in a μRAMOS device with 60 g/L fructose with different ammonium and amino acid concentrations. Depicted is the oxygen transfer rate (OTR). Cultivations were performed at 30 °C, 1000 rpm, VL = 500 μL in a 48-well round well microtitre plate at a shaking diameter of 3 mm, initial pH value: 6. AA: amino acids, crucial AA: glutamate, glycine and isoleucine. Media (with elemental nitrogen concentration): Complete Poolman medium as reference (black, 912 mgN/L) and without (NH4)2SO4 with all amino acids (pink, 700 mgN/L), with (NH4)2SO4 with crucial amino acids (purple, 345 mgN/L), with 1.5-fold (NH4)2SO4 with crucial amino acids (green, 443 mg/L N), with (NH4)2SO4 with crucial amino acids with tripled glutamate concentration (orange, 440 mgN/L). Mean values of at least 4 replicates are shown. The shadows around the curves indicate the standard deviation. For clarity, only every fifth measuring point is marked as a symbol
The cultivation in the complete Poolman medium showed the same OTR course as in previous experiments. The omission of (NH4)2SO4 from the medium did not influence the respiration, while reducing the amino acids to isoleucine, glutamate and glycine shifted the peak to 25 mmol/L/h at 18 hours. A 1.5-fold increase in (NH4)2SO4 concentration did not change the OTR. However, increasing the glutamate concentration shifted the OTR peak back to 35 mmol/L/h at 15 hours.
The omission of (NH4)2SO4 from the medium in cultivation two did not impact the growth of G. oxydans fdh, as the same respirational activity was achieved as with the complete Poolman medium. Since (NH4)2SO4 only acts as a nitrogen source, it is not essential for growth, when a sufficient amount of nitrogen is supplied via amino acids. The remaining amount of nitrogen of 700 mgN/L exceeds the necessary amount for the growth of Gluconobacter strains reported by Sainz et al. [28] (< 300 mgN/L) and Hahn et al. [23] (38 mgN/L). Moreover, amino acids can act as the sole nitrogen source for various Gluconobacter strains [23, 24]. In cultivation three, the growth of G. oxydans fdh was negatively impacted, as evidenced by the shift in the OTR. The nitrogen from the three crucial amino acids and (NH4)2SO4 amounts to 345 mgN/L, which exceeds the reported necessary concentrations [23, 28]. Since increasing the nitrogen concentration with additional supplementation of (NH4)2SO4 does not improve the growth of G. oxydans fdh, a nitrogen limitation is not the cause for the impaired growth. Increasing the glutamate concentration, however, improved the growth of G. oxydans fdh, resulting in a higher OTR peak and shorter cultivation time. Because of the importance of glutamate as a precursor for many other amino acids and the cell’s nitrogen supply [46], the demand for glutamate is increased, when only the crucial amino acids are supplemented to the medium. Consequently, the impaired growth is caused by the limited availability of glutamate. Increasing the glutamate concentration in the final medium more than three times, as well as increasing the concentrations of isoleucine and glycine two times, showed no positive influence on the cultivation of G. oxydans fdh (Fig. S2). As a result, the glutamate concentration is tripled in the final minimal medium. (NH4)2SO4 remains in the medium at the original concentration of 1 g/L as an additional nitrogen source.
Determination of essential vitamins for growth of G. oxydans fdhIn this study, 12 vitamins were examined, divided into groups like the amino acids (Table 2). Group 6 consists of the three vitamins nicotinic acid, pantothenic acid and p-aminobenzoic acid, which have already been studied by Underkofler et al. [26] and Gosselé et al. [53]. Group 7 consisted of pyridoxamine and pyridoxine, group 8 of ascorbic acid, biotin, folic acid, orotic acid and riboflavin, and group 9 of thiamine and vitamin B12. First, all groups were removed individually from the complete Poolman medium, investigating their impact on the OTR kinetics. Cultivations without groups 7 and 9 led to a shift of the OTR maximum by about 1 hour, while group 8 showed no influence (Fig. S3). It can be concluded that the components of groups 7 to 9 are not essential Fig. 4 shows the investigation of group 6. When this group is omitted from the complete Poolman medium, a strongly reduced OTR course can be observed. It can be concluded that group 6 contains essential components for the growth of G. oxydans fdh. The slight OTR increase of the experiment without group 6 was due to the transfer of media components from pre-culture and the highly active fructose dehydrogenase, as described before [14, 15]. The individual examination of the components of group 6 showed the same decreased OTR course when nicotinic acid and pantothenic acid were removed separately from the complete Poolman medium, indicating both vitamins are essential. Underkofler et al. [26] described pantothenic acid, p-aminobenzoic acid and nicotinic acid as essential vitamins for the growth of Acetobacter suboxydans. These vitamins, among others, have been used in the cultivation of G. oxydans in recent decades [21, 23, 25, 27, 54, 55]. Gosselé et al. [53] examined 95 Gluconobacter strains with regard to pantothenic acid and nicotinic acid. The results showed that 58%
留言 (0)