
記住我
The phosphatase and tensin homolog deleted on chromosome 10 (PTEN) gene, a potent tumor suppressor, is frequently mutated in human malignancies.9,54,55 The protein it expresses is a dual phosphatase with both protein and lipid phosphatase activities. Since the first discovery and identification in 1997,30 PTEN has been studied extensively in cancers, including solid tumors and lymphoid malignancies. Biochemically, PTEN plays 2 distinct roles depending on whether it is in the cytoplasm or nucleus.37,55 In the cytoplasm, PTEN antagonizes phosphoinositide 3-kinase (PI3K) and dephosphorylates (3, 4, 5) phosphatidylinositol (PIP3) by the phosphatase domain and further inhibits the threonine kinase (AKT)/mammalian rapamycin target protein (mTOR) pathway (AKT/mTOR pathway). When in the nucleus, PTEN functions through the Jun-N-terminal kinase (JNK) pathway. In addition, PTEN plays a crucial role in fundamental cellular processes, including the regulation of DNA repair, cellular senescence, and cell migration, etc.5,25,49,55
To date, a few reports have suggested the role of PTEN in neuropathic pain. The research by Huang et al. revealed that neuropathic pain induced by nerve injury was correlated with PTEN and the AKT/mTOR pathway in the spinal cord and intrathecal administration of PTEN mediated by adenovirus mitigated nociceptive sensitization.19 Another study showed that PTEN and JAK2 were involved in trigeminal neuropathic pain.31 Moreover, in vivo inhibition of PTEN increased the percentage of δ-opioid receptors expressed on the surface of trigeminal ganglion neurons and allowed efficient δR-mediated antihyperalgesia in mice.44 Although these studies confirm the role of PTEN in nociceptive signaling, they provide little information about the specific mechanism of PTEN in neuropathic pain.
Cholesterol or lipid rafts, the main lipids in the mammalian plasma membrane, are responsible for the resilience of the biological membrane and related to cellular function and viability.45,57 Cholesterol and sphingomyelin form the raft-like domains involved in the trafficking of surface proteins and the formation of dynamic signaling platforms. These raft-like domains have been demonstrated as a regulating switch in nociceptive ion channels, such as Nav1.9 and TRPV1, in neuropathic pain and inflammatory hyperalgesia.1,36,43
In this study, we characterized the specific mechanisms of PTEN's contribution to neuropathic pain that produced the following findings: (1) PTEN was expressed mainly in astrocytes and was dramatically downregulated in the spinal cord after chronic constriction injury (CCI) surgery; (2) spinal upregulation of astrocytic PTEN effectively alleviated CCI-induced pain and reduced glial activation, neuronal activity, and neuroinflammation; (3) PTEN regulated cholesterol biosynthesis mainly by interacting with enzymes involved in cholesterol biosynthesis processing; and (4) cholesterol replenishment attenuated CCI-induced pain hypersensitivity and, importantly, suppressed neuronal activity and glial activation. These results imply that disturbance of PTEN-regulated endogenous cholesterol biosynthesis in spinal astrocytes may underlie the development of pain after peripheral nerve injury, and recovery of PTEN or cholesterol might be an effective therapeutic strategy for neuropathic pain.
2. Materials and methods 2.1. Animals and surgeryAdult Sprague–Dawley rats (specific pathogen-free [SPF], female, 180-220 g) were purchased from the National Institutes for Food and Drug Control (Beijing, China). Ptenfl/fl mice were a gift from Professor Hongbing Zhang at the Chinese Academy of Medical Sciences. All animal procedures performed in this study were approved by the Institutional Animal Care and Use Committee of the Chinese Academy of Medical Sciences, Institute of Basic Medical Sciences (approval number #211-2014). Rats and mice were kept in the Laboratory Animal Center of School of Basic Medicine, Peking Union Medical College, in a temperature-controlled and humidity-controlled environment with an automatic 12-hour light–dark cycle and access to food and water ad libitum. The rat model of CCI (n = 72) was produced according to the description by Bennett and Xie6 and our previous study.56 In brief, animals were anesthetized with sodium pentobarbital (50 mg/kg, i.p., Sigma-Aldrich, St. Louis, MO), and the right sciatic nerve was exposed to place 4 loose ligatures (4-0 chromic gut) around the nerve at 1-mm intervals and then to suture muscle and skin incisions. For sham operations, the surgery was performed only to expose the right sciatic nerve but without ligation.
2.2. Drugs and administrationThe inhibitor of PTEN, SF1670, was purchased from Selleckchem (Houston, TX). Indole-3-carbinol (I3C), a potent PTEN protector,10 was purchased from Sigma-Aldrich. These 2 reagents were dissolved in dimethylsulfoxide (DMSO) first and diluted in saline solution. SF1670 was injected intrathecally (0.3 mg/kg) in naive rats32 to observe the nociceptive effect of PTEN deficiency, whereas I3C was injected in CCI rats (14 d after surgery) intrathecally (2 mg/kg)29 to verify the analgesic effects under prevention of PTEN degradation. AAV2/9-GFAP-r-PTEN-3×Flag (gPTEN), AAV2/9-GFAP-r-HMGCR-3×Flag (gHMGCR), AAV2/9-GFAP-Null (gNull), and AAV2/9-GFAP-Cre-EGFP (AAV-gCre) were synthesized by Hanbio Biotechnology (Shanghai, China). Intrathecal injection of AAV (10 µL) was performed 1 week before CCI surgery in rats. Intrathecal injection of AAV-gCre (5 µL) into Ptenfl/fl mice was conducted to generate astrocyte-specific Pten knockout (Pten CKO) mice. Water-soluble cholesterol (Sigma-Aldrich) was administered intrathecally at a concentration of 0.5 or 1 mM. All intrathecal injections were made with a 30-G needle between the L5 and L6 intervertebral spaces under isoflurane (2%) anesthesia.20
2.3. Behavioral testRats and mice were acclimatized to the testing environment for at least 2 consecutive days before baseline testing and were tested for mechanical and thermal hyperalgesia in a blind manner. Before each test, habituation was performed for 30 minutes, as described in our previous studies.33,34,56
For the Hargreaves test,15 rats (n = 48) were placed individually in a transparent plexiglass box (10 × 20 × 20 cm) placed on a 2-mm-thick glass plate, and the plantar surface of the hind paw was exposed to a beam of radiant heat from a thermal stimulator (BME-410C Plantar Test Apparatus, China). A cutoff time of 20 seconds was setup to avoid potential damage, and the test time interval between the tests of 2 feet was 5 minutes. The average of 3 recorded test times was considered the paw withdrawal thermal latency (PWL, s). For the von Frey test, rats (n = 48) were placed in a Plexiglas chamber (10 × 20 × 20 cm) placed on a wire grid floor, and the plantar surface of the hind paw was stimulated using an electronic von Frey anesthesiometer (Electronic von Frey 2390, IITC Life Science, Woodland Hills, CA). The maximum force (g) was automatically recorded, and the average of 3 successive force (g) readings was used as the paw withdrawal mechanical threshold (PWT, g). For the mouse von Frey test, the up–down method by Dixon was used.11 In brief, the plantar surface of the hind paw was simulated with a series of von Frey hairs (0.02-2.56 g, Stoelting, Wood Dale, IL). If a mouse responded to a given filament, the next lowest filament was used, or the next highest filament was used. Thirty-two mice were used for the von Frey test. For the mouse tail immersion test, the water temperature was set at 48, 50, or 52°C. The lower tail sections of mice were soaked in water, and the time of tail withdrawal from water was determined as the reaction time. The cutoff time was set at 10 seconds to avoid potential injury. Sixteen mice were used for this test.
2.4. Quantitative proteomicsRats (n = 24) were deeply anesthetized with sodium pentobarbital, and protein samples were obtained from the L5 spinal dorsal horn in rats of 4 groups (6 rats per group): naive rats, CCI 21d rats, AAV-gNull rats, and AAV-gPTEN rats. After dissolving in 8 M urea and measuring the concentration using a NanoDrop 2000 (Thermo Fisher Scientific, MA), equal amounts of 6 samples from each group were mixed to make total protein at 100 µg. Equal amounts of proteins from each sample were reduced with dithiothreitol and alkylated with iodoacetamide (IAA) before dilution in 1 M urea. Then, the samples were sent to Beijing Qinglian Biotech Co, Ltd for further analysis, including tandem mass tag (TMT) labeling, peptide fractionation with high-performance liquid chromatography, liquid chromatography–mass spectrometry/mass spectrometry (LC–MS/MS) analysis, and subsequent data analysis.
For TMT labeling, 100 mg of protein from the 4 groups diluted in 1 M urea was reduced with dithiothreitol (Little Chalfont, Bucks, United Kingdom) and alkylated with iodoacetamide (IAA, Little Chalfont). After digestion in trypsin/Lys-C (Promega, WI) for 12 hours and termination by heating at 60°C, the proteins were then labeled using the TMT Mass Tagging Kit (Thermo Fisher Scientific) following the manufacturer's instruction. Four TMT labels were used to label the different groups: TMT-129 was used for the AAV-gPTEN group, TMT-128 for the AAV-gNull group, TMT-127 for the CCI 21d group, and TMT-126 for the naive group. After labeling, samples were dissolved in 0.1% trifluoroacetic acid (TFA). For high-performance liquid chromatography separation, TMT-labeled peptides (100 μL in 0.1% TFA) were mixed equally and desalted by peptide desalting spin columns (89852, Thermo Fisher Scientific) and then fractionated using a C18 column (Waters BEH C18 4.6 × 250 mm, 5 μm) on the UltiMate 3000 UHPLC (Thermo Fisher Scientific) with a flow rate of 1 mL/min. The column oven was set at 50°C. Mobile phases A (2% acetonitrile, pH 10.0) and B (98% acetonitrile, pH 10.0) were used to develop a gradient elution. Then, fractions were dried and resolved in 0.1% TFA for mass spectrometry analysis. For LC–MS/MS analysis, peptides were separated by a 120-minute gradient elution at a flow rate of 0.300 μL/min with the EASY-nLC 1000 system, which was directly interfaced with the Thermo Orbitrap Fusion mass spectrometer. The analytical column was purchased from Thermo Fisher (75 μm ID, 150 mm length; packed with C-18 resin). Mobile phase A consisted of 0.1% formic acid, and mobile phase B consisted of 100% acetonitrile and 0.1% formic acid. The Orbitrap Fusion mass spectrometer was operated in the data-dependent acquisition mode using Xcalibur 3.0 software, and the scan cycle consisted of a single full-scan mass spectrum in the Orbitrap (350-1550 m/z, 120,000 resolution) followed by 3 seconds data-dependent MS/MS scans in an ion routing multipole at 38% normalized collision energy HCD (higher energy collisional dissociation). The MS/MS spectra from each LC–MS/MS run were searched against the selected database using the Proteome Discovery searching algorithm (version 2.1). The MS/MS spectra from each LC–MS/MS run were searched against rat fasta from UniProt using an in-house Proteome Discoverer (version PD1.4, Thermo Fisher Scientific). The search criteria were as follows: full tryptic specificity was required, 2 missed cleavages were allowed, carbamidomethylation (C) was set as the fixed modifications, oxidation (M) and TMT 6-plex were set as the variable modifications, precursor ion mass tolerances were set at 15 ppm for all MS acquired in an Orbitrap mass analyzer, and fragment ion mass tolerance was set at 0.02 Da for all MS2 spectra acquired. The peptide false discovery rate (FDR) was calculated using the Target Decoy PSM Validator provided by PD. When the q value was smaller than 1%, the peptide-spectrum match (PSM) was considered to be correct. FDR was determined based on PSMs when searched against the reverse decoy database. Peptides only assigned to a given protein group were considered unique. The FDR was also set to 0.01 for protein identification.
Relative protein quantification was performed using Proteome Discoverer software (version 2.1) according to the manufacturer's instructions on the 4 reporter ion intensities per peptide. Quantitation was performed only for proteins with 1 or more unique peptide matches. Protein ratios were calculated as the median of all peptide hits belonging to a protein. Quantitative precision was expressed as protein ratio variability.
2.5. Bioinformatic analysisRelative protein abundances are presented as ratios of TMT-127/126 (CCI 21d/naive) and TMT-129/128 (AAV-gPTEN/AAV-gNull). A log2 (127/126) ratio ≥ 0.298 and a log2 (129/128) ratio ≥ 0.383 were set as upregulated proteins, and a log2 (131/128) ratio ≤ −0.25 and a log2 (129/128) ratio ≤ −0.177 were set as downregulated proteins according to a 95% prediction interval (Supplemental Fig. 1A and B, available at https://links.lww.com/PAIN/B645). For hierarchical clustering analysis, the MS differentially expressed peptide (DEP) data (Supplemental Table S1, available at https://links.lww.com/PAIN/B645) were input into JMP Pro, and a heatmap was then constructed using hierarchical clustering. Gene Ontology (GO) analysis was performed using DAVID bioinformatics resources (https://david.ncifcrf.gov). The Wiki pathway was displayed with Cytoscape (version 3.7). Protein–protein interaction networks of DEPs were constructed with an online STRING database (http://string-db.org). “Experimental” and “database” evidence was set at a medium (0.4) confidence level.
2.6. CoimmunoprecipitationFor coimmunoprecipitation (Co-IP), protein samples from the spinal dorsal horn were harvested, lysed in NP-40–containing lysis buffer supplemented with protease inhibitor cocktail (CWBio, Beijing, China), and then immunoprecipitated as instructed by the protocol of the Pierce Crosslink Magnetic IP/Co-IP Kit (Thermo Fisher Scientific, #88805) with the indicated antibodies.
2.7. Western blottingAfter transcardial perfusion with PBS, the spinal cord was harvested and homogenized in lysis buffer (CWBio, Beijing, China) containing a protease inhibitor cocktail and a phosphate inhibitor cocktail (CWBio). Protein concentrations were measured using a Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Rockford, IL). The homogenates (20 µg total protein) were separated on 12% SDS–PAGE gels and transferred to polyvinylidene fluoride membranes. After blocking with 5% skim milk in TBST, the membrane was incubated overnight at 4°C with primary antibodies. The antibodies were as follows: PTEN (rabbit, 1:1000, Abcam, ab170941), FLAG (mouse, 1:3000, Sigma-Aldrich, F3165), HMGCR (mouse, 1:500, Santa Cruz, sc-271595), HMGCS1 (mouse, 1:500, Santa Cruz, sc-166763), MVD (mouse, 1:500, Santa Cruz, sc-376975), TNF-α (rabbit, 1:1000 Proteintech Group, 17590-1-AP), IL-1β (rabbit, 1:1000, Abcam, ab283818), IL-6 (rabbit, 1:1000, Abcam, ab6672), and GAPDH (rabbit, 1:3000, BBI Life Sciences, 110016). The membrane was further incubated with an HRP-conjugated secondary antibody for 1 hour at room temperature. Bands were detected using an eECL Kit and scanned by the ChemiDoc XRS system (Bio-Rad, Hercules, CA). The intensity of the selected bands was analyzed using ImageJ software.
2.8. Cholesterol quantitation assayFor dorsal horn total cholesterol (free cholesterol and cholesteryl esters) content determination, a Cholesterol/Cholesteryl Ester Quantitation Assay Kit (ab65359, Abcam) was used. Equal tissues (1 g) were taken from the ipsilateral and contralateral spinal cord of each rat (naive rats, CCI 21d rats, SF1670-treated or I3C-treated rats, and AAV-gNull or AAV-gPTEN or AAV-gHMGCR rats) or mouse (littermate, Ptenfl/fl, or Pten CKO). Then, the tissue was immediately processed for lipid extraction and colorimetric assay according to the manufacturer's instructions.
2.9. ImmunohistochemistryRats and mice were deeply anesthetized with sodium pentobarbital and transcardially perfused with 0.01 M PBS, followed by 4% paraformaldehyde in 0.01 M PBS. The L5 spinal cord segment was isolated and used for immunohistochemistry as described previously.24,35 In brief, the tissue sections were first permeabilized in 0.1% Triton X-100 in PBS and then blocked with 10% donkey serum for 2 hours at room temperature, followed by overnight incubation at 4°C with primary antibodies. The following primary antibodies were used: PTEN (rabbit, 1:400, Cayman Chemical, 10005059), glial fibrillary acidic protein (GFAP, mouse, 1:400, Abcam, ab4648), NeuN (mouse, 1:400, Abcam, ab104224), CD11b/c (mouse, 1:400, Abcam, ab1211), c-fos (rabbit, 1:400, CST, 2250S), anti-FLAG (DDDDK tag, rabbit, 1:400, Abcam, ab205606), and calcitonin gene-related peptide (rabbit, 1:400, CST, 14959T). Sections were then incubated with the proper secondary antibodies (Alexa Flour 488-conjugated donkey anti-mouse, 1:400; Alexa Flour 594-conjugated donkey anti-mouse, 1:400; Alexa Flour 488-conjugated donkey anti-rabbit, 1:400; and Alexa Flour 594-conjugated donkey anti-rabbit, 1:400, Jackson ImmunoResearch Laboratories) for 1 hour at room temperature. Alexa Fluor 594-conjugated IB4 (1:400, Invitrogen) was added as a secondary antibody. After washed with 0.01 M PBS and coverslipped with VECTASHIELD mounting medium with DAPI (ZSGB-BIO, China), the stained sections were examined using a laser confocal microscopic imaging system (FV1000 and Olympus FluoView software, Olympus, Japan). Images were analyzed with ImageJ software.
2.10. Statistical analysisData values are presented as the mean ± SEM. SPSS software (version 17.0) was used for statistical analysis. The Student t test was used to test the difference between 2 groups. To determine statistical comparisons of differences among 3 or more groups in western blotting and immunohistochemistry experiments, we used one-way analysis of variance followed by the Bonferroni post hoc test. Two-way analysis of variance followed by the Bonferroni post hoc test was used to determine significant differences in behavioral tests. A statistically significant difference was defined as a 2-sided P value < 0.05.
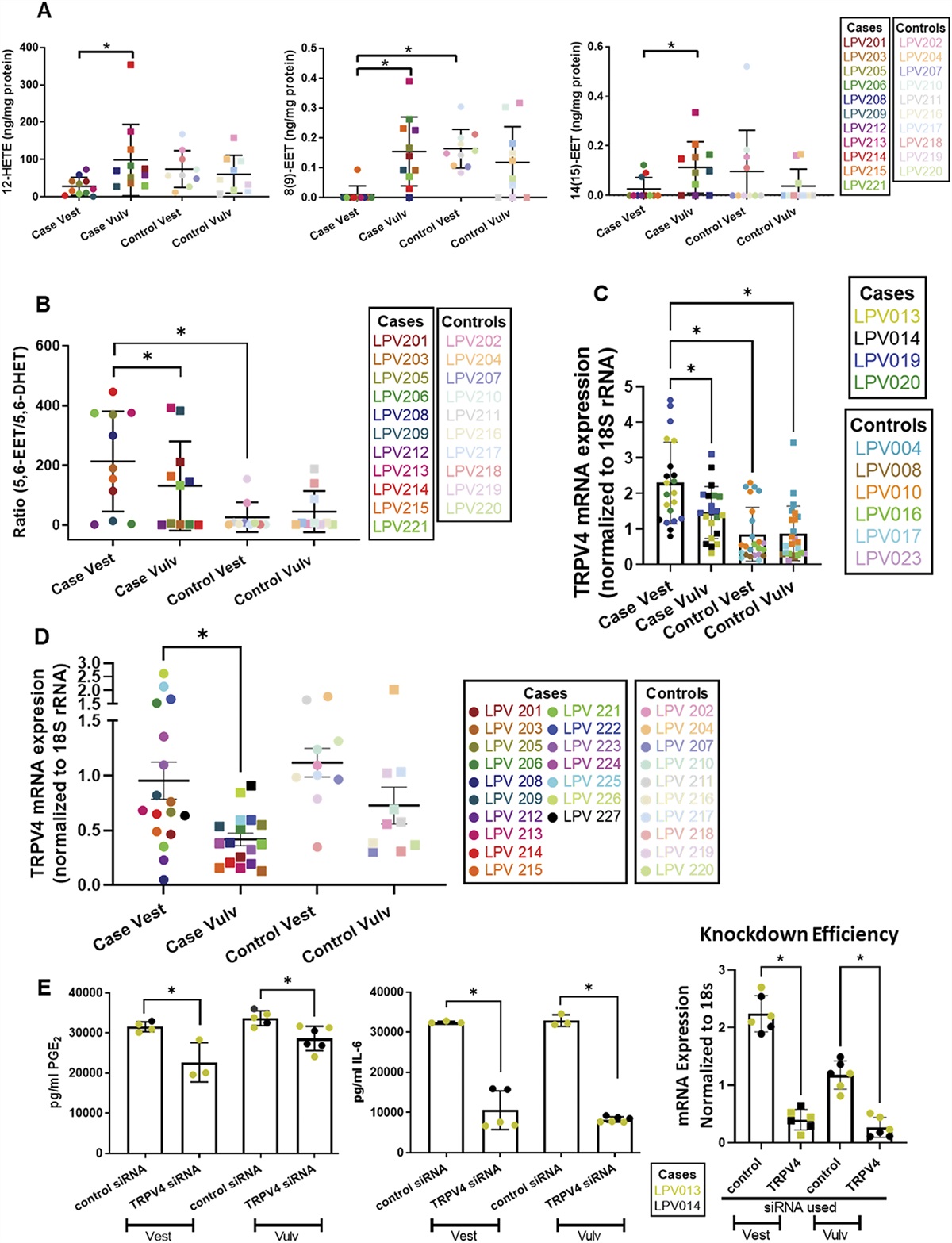
3. Results 3.1. Chronic constriction injury dramatically downregulated the expression of phosphatase and tensin homolog deleted on chromosome 10 in spinal astrocytesTo determine the expression and distribution of PTEN in the spinal cord, we performed western blotting to compare protein levels between naive, sham, CCI 7d, CCI 14d, and CCI 21d rats. The results revealed that PTEN expression in the spinal cord was reduced significantly at postoperative days 14 and 21 in the CCI group compared with naive animals (P < 0.05 vs naive group; Fig. 1A), whereas CCI 7d group rats and animals receiving the sham operation showed no significant difference (P > 0.05 vs naive group; Fig. 1A). We then performed a spinal immunofluorescence analysis using antibodies against PTEN. Compared with the sham group (Fig. 1B), immunofluorescence assessment of the ipsilateral dorsal horn at 7 d and 14 d after injury revealed significant decreases in PTEN (Fig. 1B), similar to the findings in a previous study.19 Quantification of the fluorescence intensity further confirmed the downregulation of PTEN (P < 0.01 vs ipsilateral in the sham group or contralateral in the same group; Fig. 1B) 7 and 14 days after CCI surgery.
 Figure 1.:
Figure 1.: PTEN is downregulated in spinal astrocytes after CCI surgery. (A) The time course of PTEN protein levels in ipsilateral dorsal horns from naive, sham, and CCI-operated rats. Quantification showed that CCI decreased PTEN expression at 14 days and 21 days compared with naive rats. (n = 3). (B) Representative images of PTEN immunofluorescence in the L5 dorsal horn. Scale bar, 200 μm. Immunofluorescence intensity analysis showed that the immunoreactivity of PTEN in the ipsilateral spinal dorsal horn was reduced at postoperative days 7 and 14 in the CCI group. n = 3 rats per group. *Compared with the ipsilateral dorsal horn of the sham group; #compared with the contralateral dorsal horn in the same group. (C) Double immunofluorescence labeling for PTEN and the cell-type markers CD11b/c, NeuN, and GFAP in the L5 dorsal horn in naive rats. Scale bar, 100 μm and 10 μm (insets). * or # P < 0.05, ** or ## P < 0.01. CCI, chronic constriction injury; PTEN, phosphatase and tensin homolog deleted on chromosome 10.
To clarify the specificity of PTEN expression in cells, double immunofluorescence staining was applied using different cell-type markers. The results showed that in naive rats, PTEN was almost expressed in GFAP-positive astrocytes but not in NeuN-positive neurons or CD11b/c-positive microglia (Fig. 1C). Thus, PTEN is expressed in astrocytes in the rat spinal cord.
3.2. Spinal astrocytic phosphatase and tensin homolog deleted on chromosome 10 suppresses neuropathic pain after chronic constriction injuryAfter CCI, neuropathic pain symptoms such as mechanical allodynia and heat hyperalgesia developed on day 3 and sustained for many weeks.56 To assess the role of PTEN in neuropathic pain, the PTEN inhibitor SF1670 was intrathecally (i.t.) injected into naive rats. Then, we checked the time course of pain behavior 6 hours, 12 hours, 1 day, and 2 days after injection. The i.t. administration of SF1670 (0.3 mg/kg, 15 μL) significantly induced thermal hyperalgesia (PWL) and mechanical allodynia (PWT) from 6 hours to 1 day postinjection compared with that of i.t. injection of DMSO (P < 0.01 vs DMSO injection group; Figs. 2A–C). Rats returned to baseline levels after 2 days of injection.
 Figure 2.:
Figure 2.: Astrocytic PTEN attenuates CCI-induced mechanical allodynia and thermal hyperalgesia. (A) Schematic time line for intrathecal administration and behavioural tests. (B and C) Intrathecal injection of the inhibitor of PTEN, SF1670, induced thermal hyperalgesia (B) and mechanical allodynia (C) in naive rats (n = 6). (D and E) Intrathecal injection of I3C (n = 7) alleviated CCI-induced thermal hyperalgesia (D) and mechanical allodynia (E). (F and G) CCI-induced thermal hyperalgesia (F) and mechanical allodynia (G) were attenuated in rats receiving intrathecal injection of AAV-gPTEN 7 days before CCI. n = 6. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. i.t., intrathecal injection. Arrows, SF1670 or I3C injection; arrowheads, CCI surgery. CCI, chronic constriction injury; PTEN, phosphatase and tensin homolog deleted on chromosome 10.
We further examined the antinociceptive effect of indole-3-carbinol (I3C) in CCI rats. I3C is a derivative of glucobrassicin in cruciferous vegetables, which inhibits WWP1-mediated PTEN polyubiquitination and ultimately reactivates PTEN.10 Consistent with previous reports,2,10,19 CCI surgery–induced mechanical allodynia and thermal hyperalgesia were sustained for several weeks. After we intrathecally injected I3C (2 mg/kg, 15 μL) on day 14 after CCI surgery, thermal hyperalgesia induced by CCI operation was alleviated at 6 hours, 12 hours, and 3 days after I3C injection (P < 0.01; Fig. 2D). Meanwhile, mechanical allodynia was also markedly attenuated. The antiallodynic effect became evident as early as 6 hours after the injection and was maintained for 4 days (P < 0.01; Fig. 2E).
To further investigate the function of PTEN in spinal astrocytes, injection (i.t.) of AAV9-mediated astrocytic PTEN (AAV-gPTEN) was conducted to overexpress spinal PTEN in naive rats 7 days before CCI injury. Then, we tested the rat behavior until 21 days after surgery. In contrast with vehicle injection (AAV-gNull), i.t. injection of AAV-gPTEN significantly attenuated CCI-induced thermal hyperalgesia at days 7, 14, and 21 after CCI (P < 0.001 or 0.0001; Fig. 2F) and mechanical allodynia from 7 to 21 days after CCI (P < 0.001 or 0.0001; Fig. 2G). After behavior testing, we harvested the L4-6 spinal cord on day 30 after AAV injection. Further immunofluorescence and western blot assessment of the spinal cord indicated that FLAG and PTEN expression levels were upregulated by more than 2-fold compared with vehicle injection (Figs. 3A and B) and that FLAG and PTEN were expressed specific in astrocytes (Figs. 3C–E). Together, these results suggested that the expression level of PTEN in spinal astrocytes was successfully upregulated by i.t. AAV-gPTEN administration.
 Figure 3.:
Figure 3.: i.t. administration of AAV-mediating PTEN upregulates PTEN expression levels in spinal astrocytes. (A) Representative gel bands and quantification of PTEN and FLAG proteins. (B) Immunofluorescence staining of PTEN in the spinal dorsal horn in rats at sham 21days, CCI 21 days , and i.t. injection AAV (AAV‐gPTEN or AAV‐gNull) rats at CCI 21 days. Scale bar, 100 μm. Immunofluorescence intensity analysis showed that the immunoreactivity of PTEN in the bilateral spinal dorsal horn was reduced at postoperative day 21 in the CCI group but reversed in the AAV-gPTEN group. n = 3. *compared with the ipsilateral dorsal horn in the sham group; #compared with the ipsilateral dorsal horn in the AAV-gNull group. (C) Representative fluorescence image showing FLAG expression in the spinal cord 21 days after i.t. injection of AAV. Scale bar, 200 μm. (D) Double immunofluorescence labeling of FLAG with GFAP, NeuN, and CD11b/c in the L5 dorsal horn. Scale bar, 50 μm. (E) Double immunofluorescence labeling for PTEN and cell-type markers in AAV-gPTEN rats. Scale bar, 50 μm. PTEN, phosphatase and tensin homolog deleted on chromosome 10.
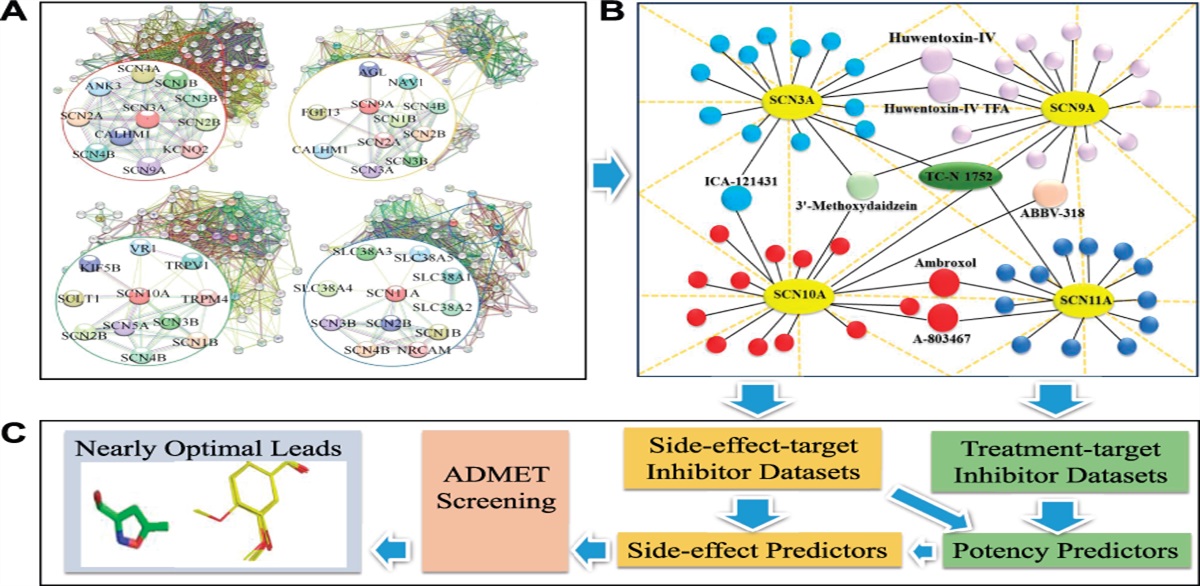
3.3. Astrocytic PTEN is involved in the cholesterol biosynthesis pathway of the spinal dorsal hornTo figure out the potential downstream regulatory target of PTEN in the spinal dorsal horn of CCI rats, TMT-based quantitative proteomics was performed using samples from the dorsal horn of 4 groups: naive rats, CCI 21d rats, AAV-gNull rats, and AAV-gPTEN rats. A total of 4697 proteins (Score SEQUEST HT > 0) were identified, 221 of which were differentially expressed (Supplemental Table S1, available at https://links.lww.com/PAIN/B645). The DEPs were then grouped into 4 clusters based on the protein expression ratio and are shown in the heatmap (Fig. 4A). The DEPs of the CCI model in cluster 2 (upregulated proteins) and cluster 3 (downregulated proteins) were reversed by PTEN overexpression. The next GO classification indicated that DEPs in cluster 3 were enriched in the cholesterol biosynthesis process, isoprenoid biosynthetic process, intracellular protein transport, and leucine metabolic process (Fig. 4B, details in Supplemental Table S2, available at https://links.lww.com/PAIN/B645). Next, the proteins in cluster 2 and cluster 3 were then analyzed with STRING, respectively. The protein–protein interaction results of cluster 3 (Fig. 4C) showed that 7 enzymes catalyze cholesterol biosynthesis, including HMGCR, HMGCS1, MVD, MVK, acetoacetyl-CoA synthetase, phosphatidate cytidylyltransferase 2, and PCYT1B. These enzymes were downregulated in the CCI group and upregulated by PTEN overexpression. These data suggest that PTEN regulates enzymes of the cholesterol biosynthesis process and may ultimately affect cholesterol content in CCI rats.
 Figure 4.:
Figure 4.: Proteome and bioinformatic analyses show that PTEN upregulates enzymes in the cholesterol biosynthesis pathway. (A) Heatmap constructed into clusters by JMP (version Pro 13) shows contrasting and similar expression levels of DEPs in the CCI (127/126) group and corresponding proteins in the PTEN overexpression (129/128) group. MS data were put into JMP with log2 transformation. C1 to C4 refers to clusters 1 to 4. (B) The biological process part of the GO analysis of downregulated proteins in the CCI model (127/126) group. The red column represents the significance of the enrichment result (−log10(P value)); the blue column represents the gene count in each enrichment term. (C) The DEPs of C3 in the protein–protein interaction diagram are shown as nodes. The left panel shows the network of DEPs in the CCI model (127/126) group, and the right panel shows the DEPs in the PTEN overexpression (129/128) group. MS data matched to STRING networks with log2 transformation. Upregulation or downregulation of identified proteins is indicated by colors in either the heatmap or STRING network (upregulated in red and downregulated in green). DEPs, differentially expressed peptides. (D) HMGCR, HMGCS1, and MVD protein levels in the ipsilateral dorsal horn were downregulated at postoperative day 21 in the CCI group and were reversed by injection (i.t.) of AAV-gPTEN. (E) Coimmunoprecipitation (Co-IP) showed the interaction between PTEN and HMGCR, HMGCS1, and MVD. (F) Quantification of cholesterol levels in the spinal cord 6 hours after intrathecal injection of SF1670 in naive rats (n = 6) and I3C in CCI rats at 14 d (n = 6). (G) Quantification of cholesterol levels in the ipsilateral dorsal horn in rats at sham 21 days, CCI 21days, and i.t. injection AAV (AAV‐gPTEN and AAV‐gNull) rats at CCI 21 days. (n= 4‐7/group). (H) Diagram showing that PTEN may directly interact with enzymes in the cholesterol synthetic pathway and affect cholesterol content. CCI, chronic constriction injury; DEPs, differentially expressed peptides; HMGCR, 3-hydroxy-3-methylglutaryl-CoA reductase; PTEN, phosphatase and tensin homolog deleted on chromosome 10.
Next, we checked the protein expression of 3 enzymes (HMGCR, HMGCS1, and MVD) catalyzing cholesterol biosynthesis and cholesterol content according to the results of quantitative proteomics. Western blotting showed that the protein levels of HMGCR, HMGCS1, and MVD were significantly downregulated in CCI rats at 7 d and 21 d (P < 0.05 or P < 0.01 vs sham or naive) and were reversed by pretreatment with AAV-gPTEN (P < 0.01 vs AAV-gNull group; Fig. 4D). We then explored how PTEN regulates the expression level of enzymes in the cholesterol biosynthesis pathway. Because PTEN is a phosphatase with a dephosphorylation function, we detected its interactions with HMGCR, HMGCS1, and MVD through Co-IP experiments using spinal dorsal horn tissues in naive rats. As shown in Figure 4E, immunoprecipitation with commercially available anti-PTEN antibodies resulted in coprecipitation of HMGCR, HMGCS1, and MVD. Immunoprecipitation with anti-IgG antibody was used as a negative control. To further confirm the specificity of this interaction, immunoprecipitation with commercially available anti-HMGCR, HMGCS1, or MVD antibodies was performed in spinal dorsal horn tissue. Similarly, immunoprecipitation of these 3 enzymes resulted in coprecipitation of PTEN (Fig. 4E). These results demonstrate a physical interaction between PTEN and enzymes (HMGCR, HMGCS1, and MVD).
We then quantified the total cholesterol (free cholesterol and cholesteryl esters) contents of weighed ipsilateral dorsal horn tissue (1 g) from SF1670-injected rats, I3C-injected rats, and AAV-treated rats. In contrast with injection (i.t.) of an equal volume of DMSO, SF1670 administration significantly (13 ± 3.2%) decreased cholesterol levels in naive rats, whereas I3C injection (i.t.) exhibited a 12 ± 4.2% increase in cholesterol levels (Fig. 4F) in CCI 14d rats. In addition, CCI surgery (CCI 21d) induced a 12 ± 4.2% reduction in cholesterol levels, whereas pretreatment with AAV-gPTEN and AAV-gHMGCR (7 d before surgery) reversed this decreasing trend (Fig. 4G). The present data indicate that peripheral nerve injury decreases the cholesterol level in the spinal dorsal horn and that PTEN accelerates cholesterol synthesis.
Together, the above results indicated that PTEN may directly interact with enzymes catalyzing cholesterol biosynthesis, further intervening in cholesterol levels and ultimately regulating pain (Fig. 4H).
3.4. Astrocytic HMGCR overexpression relieves neuropathic pain after chronic constriction injuryHMGCR is the rate-limiting enzyme in cholesterol biosynthesis and close coupling with PTEN. Therefore, we tested whether HMGCR, like PTEN, is also essential for CCI-induced pain. Injection (i.t.) of AAV9-mediated astrocytic HMGCR (AAV-gHMGCR) was conducted to overexpress spinal HMGCR in naive rats 7 days before CCI injury. The western blot of HMGCR expression level and immunofluorescence of FLAG checked successful injection of AAV and showed that FLAG was almost expressed in GFAP+ astrocytes (Figs. 5A–E). The subsequent cholesterol content determination verified that the total cholesterol level increased in the HMGCR overexpression group (Fig. 5F).
 Figure 5.:
Figure 5.: Astrocytic HMGCR alleviates CCI-induced neuropathic pain. (A) Representative western blots and quantification of HMGCR showing overexpressed HMGCR levels in the AAV-gHMGCR group. n = 3. #compared with the AAV-gNull group. (B–E) The expression of the FLAG tag in the spinal cord after intrathecal injection of AAV-gHMGCR. (B) Representative fluorescence image showing FLAG expression in the spinal cord 21 days after i.t. injection of AAV. Scale bar, 200 μm. (C–E) Double immunofluorescence labeling of FLAG with GFAP (C), NeuN (D), and CD11b/c (E) in the L5 dorsal horn. Scale bar, 50 μm. (F) Quantification of cholesterol levels in the ipsilateral dorsal horn in sham, CCI 21days rats and i.t. injection AAV (AAV‐gHMGCR or AAV‐gNull ) rats (n =4‐5/group). (G and H) CCI-induced thermal hyperalgesia (G) at day 7 and mechanical allodynia (H) at days 7, 14, and 21 were attenuated in rats that received i.t. administration of AAV-gHMGCR. n = 6 rats/group. CCI, chronic constriction injury; HMGCR,3-hydroxy-3-methylglutaryl-CoA reductase.
Then, we tested the pain-like behavior of rats until 21 days after CCI. In contrast with vehicle injection (AAV-gNull), i.t. injection of AAV-gHMGCR significantly attenuated CCI-induced thermal hyperalgesia at day 7 after CCI (P < 0.05, Fig. 5G) and mechanical allodynia from 7 to 21 days after CCI (P < 0.05 or 0.0001; Fig. 5H). Thus, these data indicate that astrocytic HMGCR is essential for CCI-induced pain behavior.
3.5. Cholesterol replenishment alleviates chronic constriction injury–induced neuropathic painWe then tested the effect of cholesterol replenishment in vivo on CCI-induced neuropathic pain. Water-soluble cholesterol was intrathecally injected into rats (injection of saline solution as a control) on postoperative day 14. We observed that both thermal hyperalgesia and mechanical allodynia induced by CCI surgery were significantly attenuated for at least 5 hours after 1 mM cholesterol injection (P < 0.05 or P < 0.01 vs saline; Fig. 6A and B). Cholesterol injection (0.5 mM) (i.t.) also alleviated thermal hyperalgesia in CCI rats for 4 hours (P < 0.001 vs saline; Fig. 6A) but showed no significant inhibitory effect on mechanical allodynia (P > 0.05 vs saline; Fig. 6B). Thus, our data show an antalgic effect of cholesterol supply on neuropathic pain.
 Figure 6.:
Figure 6.: Cholesterol replenishment alleviates neuropathic pain and glial activation in CCI rats. (A and B) Injection (i.t.) of water-soluble cholesterol (both 0.5 mM and 1 mM) alleviated CCI-induced thermal hyperalgesia (A) and mechanical allodynia (B) in comparison with vehicle (saline solution) injection. n = 7/group. Arrows, cholesterol injection. (C) Astrocyte and microglial activation were inhibited by cholesterol injection in a dose-dependent manner. Scale bar, 100 μm. n = 3/group. *compared with the saline solution injection (vehicle) group. (D) Representative images of GFAP and CD11b/c immunofluorescence staining in the L5 dorsal horn in sham, CCI, and i.t. injection AAV (AAV‐gPTEN or AAV‐gNull) rats. Scale bar, 200 μm. Immunofluorescence intensity analysis shows that the immunoreactivity of GFAP (left) and CD11b/c (right) in the ipsilateral spinal dorsal horn was reduced at postoperative day 21 in the CCI group. n = 3 rats per group. * compared with the sham group, # compared with the AAV-gNull group. (E) Immunofluorescence staining showing that the immunoreactivity of GFAP in the ipsilateral dorsal horn was reduced in rats receiving AAV-gHMGCR. n = 3. Scale bar, 100 μm. (F) Immunofluorescence staining showing that immunoreactivity of CD11b/c in the ipsilateral dorsal horn was reduced in rats receiving AAV-gHMGCR. n = 3, compared with the AAV-gNull group. *P < 0.05; **or ##P < 0.01, and *** P < 0.001. pre, presurgery; CHOL, cholesterol; CCI, chronic constriction injury.
3.6. Phosphatase and tensin homolog deleted on chromosome 10–HMGCR–cholesterol pathway suppresses glial activation and the release of proinflammatory cytokinesActivation of spinal glial cells, such as microglia and astrocytes, leads to central sensitization and eventually increases neuronal excitability, which has been demonstrated to play a pivotal role in the pathogenesis of neuropathic pain.22,23,59 We first investigated the activation of glial cells in CCI rats using antibodies against GFAP and CD11b/c. Spinal immunofluorescence analysis showed obvious activation of astrocytes and microglia (∼2-fold increase compared with the sham group; Supplemental Fig. 2A–H, available at https://links.lww.com/PAIN/B645) in the ipsilateral spinal cords 7 and 14 days after CCI. However, the immunoreactivity of NeuN, a neuronal-specific nuclear protein, did not show a significant difference (P > 0.05, vs sham, Supplemental Fig. 2I–L, available at https://links.lww.com/PAIN/B645) in CCI rats.
Next, we explored whether PTEN affected glial activation after CCI. Both astrocyte (GFAP+) and microglial (CD11b/c+) activation were significantly reduced in the ipsilateral dorsal horn in AAV-gPTEN rats (P < 0.01 vs AAV-gNull group; Fig. 6C). By contrast, the contralateral dorsal horn did not exhibit a significant difference in glial activation after AAV-gPTEN injection (Fig. 6C). We then checked the expression level of proinflammatory cytokines (TNF-α, IL-1β, and IL-6). In the ipsilateral dorsal spinal cord, all 3 of them were markedly increased at postoperative day 21 in CCI rats but decreased after the injection of AAV-aPTEN (P < 0.01 or 0.001 vs sham or AAV-null group Supplemental Fig. 3A, available at https://links.lww.com/PAIN/B645). These data indicate that PTEN plays an essential role in the activation of spinal astrocytes and microglia after CCI.
We also observed the effect of cholesterol replenishment on glial activation. The immunostaining results revealed that both microglia (CD11b/c+) and astrocyte (GFAP+) activation were significantly reduced in the ipsilateral dorsal horn in cholesterol-treated rats (P < 0.01 or P < 0.05 vs ipsilateral dorsal horn in the vehicle group; Fig. 6D). By contrast, the contralateral dorsal horn did not exhibit a difference in glial activation after cholesterol supply (Fig. 6D). These data suggest the potential mechanism of cholesterol involvement in nociceptive behavior. Next, we found that both astrocyte (GFAP+) and microglial (CD11b/c+) activation were significantly reduced in the ipsilateral dorsal horn in AAV-gHMGCR rats (P < 0.05 vs AAV-gNull group; Fig. 6E and F). Overall, the above results suggest that the PTEN–HMGCR–cholesterol pathway mediates nociception at least partly by reducing central sensitization. Moreover, the expression level of proinflammatory cytokines (TNF-α, IL-1β, and IL-6) in the ipsilateral dorsal spinal cord were markedly increased at postoperative day 21 in CCI rats but decreased after the injection of AAV-gHMGCR (P < 0.05 or 0.01 vs sham or AAV-null group Supplemental Fig. 3B, available at https://links.lww.com/PAIN/B645). Thus, these data indicate that astrocytic HMGCR is essential for CCI-induced central sensitization.
Next, we confirmed the activation of pain signals at the cellular level by examining the expression of neuronal activity marker, c-fos. The immunohistofluorescence analysis showed abundant expression of c-fos protein in ipsilateral superficial laminae of the spinal dorsal horn 21 d after CCI (Supplemental Fig. 3C, available at https://links.lww.com/PAIN/B645). i.t. injection of AAV-gPTEN significantly reversed the activation of c-fos (P < 0.001 vs ipsilateral dorsal horn in the AAV-gNull group; Supplemental Fig. 3D, available at https://links.lww.com/PAIN/B645). The following western bolt result verified that the expression level of c-fos in the ipsilateral dorsal horn was markedly increased at postoperative day 21 in CCI rats but decreased after the injection of AAV-aPTEN (Supplemental Fig. 3E, available at https://links.lww.com/PAIN/B645). In addition, i.t. injection of cholesterol also reversed the activation of c-fos (P < 0.05 vs ipsilateral dorsal horn in the vehicle group; Supplemental Fig. 3F, available at https://links.lww.com/PAIN/B645).
Overall, the above results suggest that the PTEN–HMGCR–cholesterol pathway is involved in both the neuronal activity and central sensitization that results in neuropathic pain.
3.7. HMGCR overexpression reverses conditional knockout of astrocytic PTEN-induced pain in miceWe conducted intrathecal injection of AAV-gCre (5 µL) into Ptenfl/fl mice to generate astrocyte-specific Pten knockout (Pten CKO) mice (Fig. 7A). The GFP was expressed only in GFAP-positive astrocytes, and spinal PTEN expression was specifically lost in astrocytes in Pten CKO mice, as shown by immunofluorescence staining and western bolting (Figs. 7B–F, Supplemental Fig. 4A–D, available at https://links.lww.com/PAIN/B645), indicating the successful injection of AAV and Pten knockout. Pten CKO mice showed naive gross anatomy (Supplemental Fig. 4E, available at https://links.lww.com/PAIN/B645). Pten CKO mice also showed normal distribution patterns in the dorsal horn of the cell marker NeuN (Fig. 7G) and normal innervations of calcitonin gene-related peptide (Supplemental Fig. 4F, available at https://links.lww.com/PAIN/B645) and IB4 (Supplemental Fig. 4G, available at https://links.lww.com/PAIN/B645).
 Figure 7.:
Figure 7.: Astrocytic PTEN conditional knockout mice show mechanical allodynia and glial activation. (A) Diagram showing the strategy used to establish astrocyte-specific PTEN conditional knockout (Pten CKO) mice. (B) Immunostaining shows PTEN in GFAP-positive astrocytes in littermates. Scale bar, 50 μm. (C) Representative fluorescence image showing GFP and PTEN
留言 (0)