
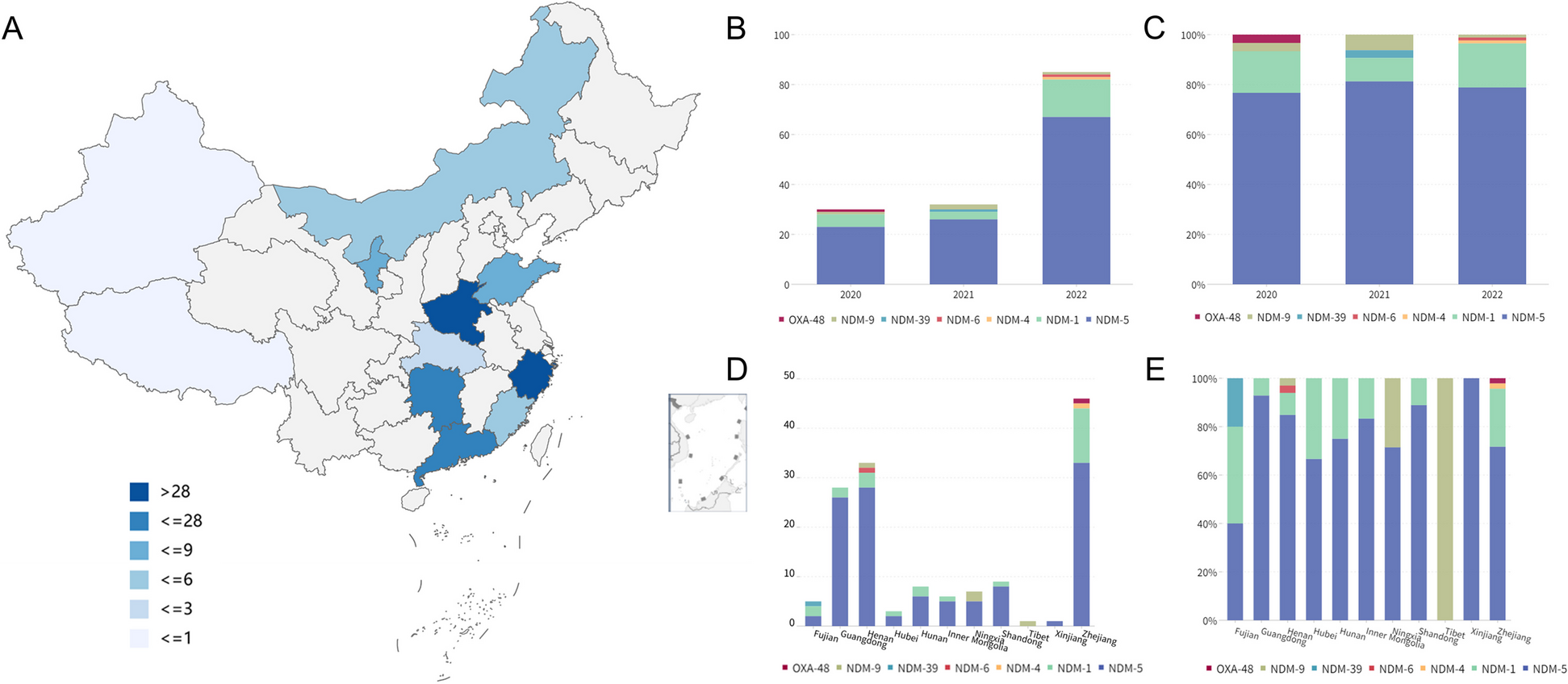
記住我


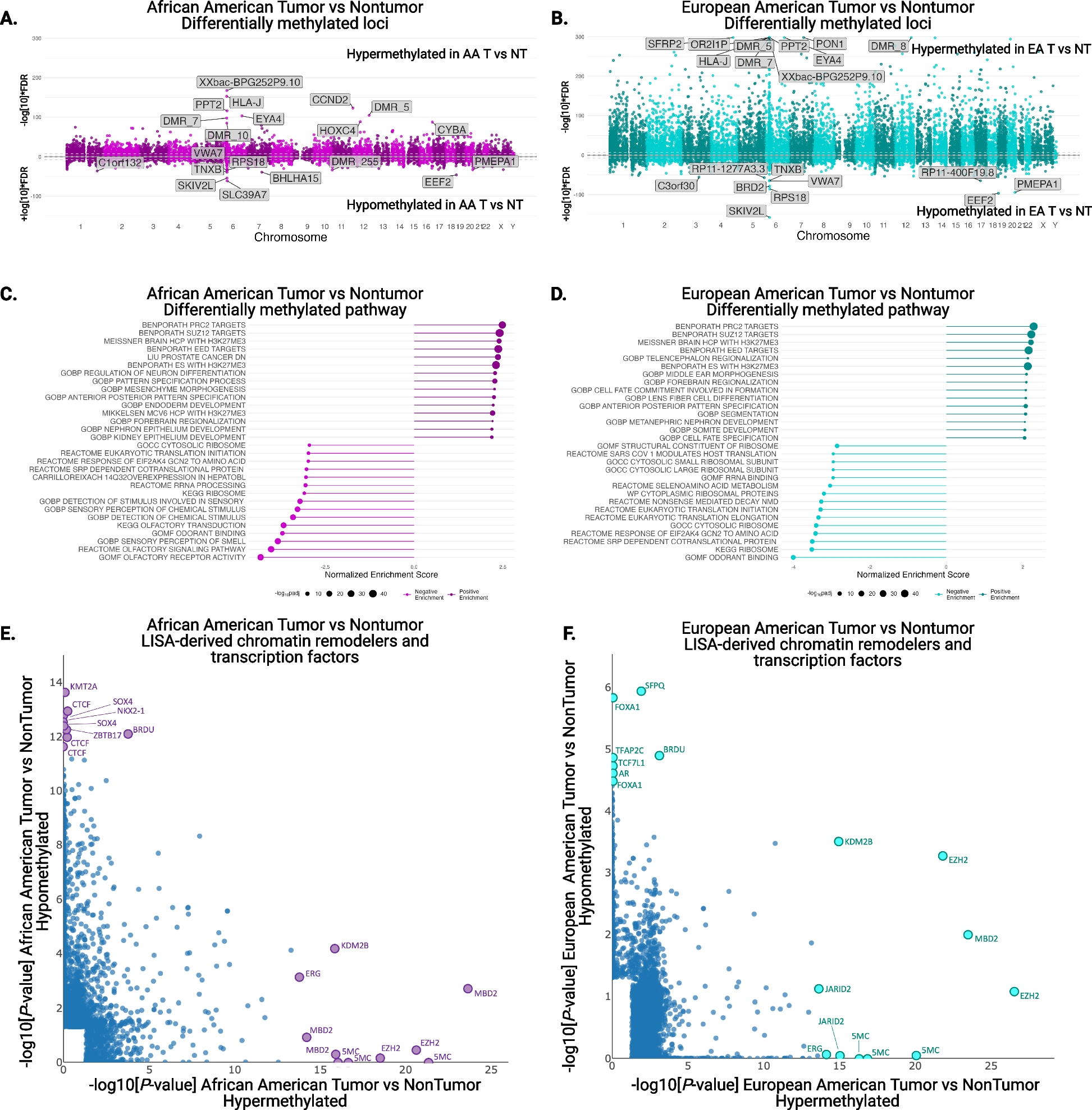
A total of 22 successful DPM models (29% of 75 total attempts) were established from 22 patients engrafted between the year 2014 and 2018. The demographic breakdown of the patients from whom the 22 DPM PDX models were established is noted in Table 1 and Additional file 1: Table S1. The median time to engraftment was 3.3 months (range: 0.5–9.2 months), and there was no significant difference between engraftment time for epithelioid (n = 15; median time to engraftment: 3.7 months, range 0.5–9.2) and non-epithelioid PDXs (n = 7; median time to engraftment: 3.3 months, range 0.8–3.9 months; p = 0.12). Immunohistochemistry (IHC), histologic subtyping, and targeted next generation sequencing (NGS) with MSK-IMPACT were performed on all patient and PDX models with available material. RNA sequencing was performed on all PDX models with available material that passed quality control (n = 18; Fig. 1A).
Table 1 Patient demographics at the time of PDX collectionFig. 1
Generation of PDX models and patient treatment histories. A Graphical overview of PDX collection and analysis. Samples were obtained by surgical resection (pleurectomy/decortication), biopsy, and aspirations. Both PDX and human samples were analyzed by IHC, targeted next generation sequencing (MSK-IMPACT), and histologic subtyping when material was available. RNA sequencing was performed on all PDX models with available data. The “n” represents the number of samples run at each step of the analysis. B Swimmers plot showing the clinical course of all 22 patients where the red arrow denotes the time of tissue collection for the PDX and C details of systemic therapy received prior to PDX collection in the 11 applicable patients. E, epithelioid; B, biphasic; S, sarcomatoid
Most patients were male (86%), current/former smokers (68%), and had stage I–IIIA disease at the time of PDX sample collection (64%). Half the patients (n = 11) had at least one systemic therapy prior to PDX sample collection. When comparing epithelioid to non-epithelioid histology, there were no significant differences in the distribution of age (p = 0.53), sex (p = 0.53), smoking status (p = 0.071), or receipt of systemic therapy prior to tissue collection for PDX generation (p = 0.56). The median clinical follow-up time was 16.2 months (range, 6.2–50.7 months). The median overall survival (OS) of the entire cohort was 17.4 months (range 6.2–50.8 months). OS at 6, 12, and 24 months after diagnosis was 100%, 81%, and 17%, respectively. At the time of last follow-up, 16 patients had died (73%).
Patients with epithelioid versus non-epithelioid disease had a median progression-free survival (PFS) on platinum therapy of 10.5 vs 7.3 months (Additional file 2: Fig. S1A; p = 0.54; hazard ratio [HR] = 0.74) and a median OS of 21.4 versus 12.5 months (Additional file 2: Fig. S1B; p = 0.13; HR = 0.46), respectively.
Treatment histories of the 22 patients are noted in Table 1 and Fig. 1. Seven PDX models were generated from resection specimens from pleurectomy/decortications, while the remaining 15 were from either biopsies or aspirations. During the study follow-up, pleurectomy/decortication was attempted in 45% (n = 10) of patients and 45% (n = 10) received thoracic radiation. Half the patients had at least one systemic therapy prior to PDX sample collection (Fig. 1B, C) which included platinum/pemetrexed (n = 11), immune checkpoint inhibitors (n = 3), vinorelbine with or without gemcitabine (n = 2), and other investigational agents (n = 3; including chimeric antigen receptor T cell therapy [NCT02414269] in MSK_LX892, tazemetostat [NCT02860286] in MSK_LX590, and LY3023414 [NCT01655225] in MSK_LX154). All but three patients initiated treatment (surgical or systemic) within 3 months of diagnosis: MSK_LX606/MSK_LX307 were delayed due to patient preference and MSK_LX413 due to an amended diagnosis from mesothelial hyperplasia to DPM (upon review at our institution).
Histology was conserved between matched PDX and clinical mesothelioma samplesNext, we performed a detailed histologic assessment on all patient samples with epithelioid DPM (n = 15) as well as matched PDX models with available material for review (n = 11) to identify the major architectural patterns/cytologic features (both predominant as well as the presence of any pattern (trabecular [T], tubulopapillary [TP], solid [S], micropapillary [MP], and pleomorphic [P]) and nuclear grade, using Kadota/MSKCC grading system, to assess for histologic features that are known to be prognostically significant in epithelioid DPM [3, 29, 30, 35]. We compared the histologies of the patient samples with a tissue microarray (TMA) derived from the matched PDX models. Of note, not all PDX samples were included on the TMA due to absence of available cores.
The predominant architectural pattern in the 15 DPM epithelioid histology tumors included the following: S (n = 7, 47%), T (n = 4, 26.4%), and TP (n = 4, 26.5%) (Table 2). Unfavorable architectural patterns and/or cytologic features [3, 29, 36] were seen in the majority of epithelioid DPM (80%; n = 12) with S pattern in 12 and MP pattern and P features present in 2 and 1 tumors, respectively. S pattern was seen in all 11 paired PDX samples and was predominant in 9 (82%). Concordant predominant architectural patterns were observed in 64% (7/11) of paired patient and PDX samples. In the four cases with discrepant predominant patterns, the predominant pattern noted in the patient tissue was a favorable pattern (i.e., T (n = 2) and TP (n = 2)), while the predominant pattern in the corresponding PDX samples were S (i.e., unfavorable) [3, 36]. The predominant pattern noted in these 4 patient samples were present as a non-dominant histology in MSK_LX413 but not in MSK_LX333, MSK_LX944B, and MSK_LX96 which may be attributable to tumor heterogeneity of the sampled material.
Table 2 Histologic subtyping of available patient samples and PDX modelsHigher nuclear grade has been shown to be associated with worse prognosis in epithelioid DPM [3, 30, 35]. Nuclear grades of epithelioid tumors in patient tissue and in PDX samples were assessed and annotated as grade I (n = 3, 20%; n = 1, 9%), grade II (n = 7, 47%; n = 5, 45.5%), and grade III (n = 5, 33%; 5, 45.5%), respectively. Concordance between the nuclear grade of the patient and PDX samples was 64% (7/11) in the paired epithelioid samples. When evaluating our data from the 11 PDX models with epithelioid DPM to other publicly available datasets, we found relative enrichment of unfavorable histologic features including (predominant S pattern: 82%, nuclear grade III: 45%) in our cohort compared to 232 epithelioid DPM from previously published work from our group (S predominant: 38% [37], nuclear grade III: 15% [30]) and a multi-institutional study of 776 tumors [35] (nuclear grade III: 16%).
For the seven non-epithelioid patient specimens, paired PDX samples were available for five. Three of the five (60%) pairs showed concordant histology (Table 3). MSK_LX759 and MSK_LX651 were classified as biphasic in the patient samples (both 90% epithelioid in patient tissue), while only epithelioid histology was noted in the paired PDX samples; this discrepancy may be due to sampling bias from the predominantly epithelioid patient sample, clonal selection in model generation, and/or the region cored for analysis on the PDX TMA.
Table 3 Histologic subtyping of available patient samples and PDX modelsPatterns of mesothelioma markers in patient samples and PDX specimens were similar by immunohistochemistry (IHC)Next, IHC markers of interest were evaluated. Concordance was defined as expression within ± 25% for the PDX and patient samples. IHC was performed on the 16 cases available in the PDX TMA and compared to available paired patient samples (Fig. 2A; Additional file 3: Fig. S2-S5). BRCA-1-associated protein (BAP1) is a tumor suppressor gene which shows biallelic inactivation in approximately half of all mesotheliomas [38, 39]. BAP1 expression was retained in 69% (11/16) of PDX samples and 64% (9/14) of patient samples (Additional file 3: Fig. S2). Concordance in BAP1 expression in the available paired patient and PDX samples was 100% (14/14) (Additional file 3: Fig. 2A). WT1 is a useful marker for identifying DPM and may also play a role in chemotherapy resistance in the disease [40,41,42]. Among PDX samples, 94% (15/16) were WT1 positive and there was high concordance (86% [12/14]) with matched clinical samples (Fig. 2A; Additional file 3: Fig. S2). Mesothelin (MSLN) is a glycophosphatidylinositol-linked cell surface protein highly expressed in several types of malignant tumors, including mesothelioma [43, 44]. There was high expression of mesothelin both in the PDX models (94% [15/16]) and patient samples (92% [12/13]). There was a high rate of concordance between PDX and patient samples in those with available paired material for analysis (77% [10/13]; Fig. 2A; Additional file 3: Fig. S2).
Fig. 2
Comparative immunohistochemistry and next generation sequencing of patient samples and PDX models. A Concordance of IHC markers between PDX and patient samples. BAP1 concordance was defined as loss/retained. WT1, mesothelin, VISTA, and PD-L1 concordance was defined as the PDX and patient sample expression being within ±25% expression of each other. B Genomic landscape of patients’ samples (n = 19), C PDX samples (n = 22), and D paired patient/PDX samples (n = 19) with available material using MSK-IMPACT targeted next generation sequencing. Genes were annotated if noted to have at least one alteration in the patient cohort
Expression of programmed death ligand 1 (PD-L1) has been studied as a prognostic biomarker in several tumors given its central role in antitumoral immune response evasion. While mesothelioma cells can express PD-L1, the association of PD-L1 expression with clinical outcomes on immunotherapy is controversial [8, 12, 45,46,47]. PD-L1 expression was relatively low across PDX samples and patient samples, but concordance was high, 100% (14/14) of cases (Fig. 2A; Additional file 3: Fig. S2). Lastly, we evaluated VISTA (V-domain Ig-containing suppressor of T cell activation), an inhibitory T cell checkpoint protein frequently expressed in DPMs independent of PD-L1 expression [34, 48]. In our cohort, VISTA expression was detected in 67% (8/12) of samples. Expression in PDX and matched clinical samples was 67% (8/12) concordant.
MSK-IMPACT targeted tumor sequencing of matched PDX and clinical mesothelioma samplesWhile our understanding of molecular diagnostics in mesothelioma has advanced [34, 49, 50], our ability to exploit identified genomic alterations remains elusive. Establishing murine models which faithfully recapitulate the genomics of a patient’s tumor is imperative to therapeutic progress. We compared the genomic landscapes of our patient tumors and PDX models to establish the genomic fidelity.
There were 19 patient samples and 22 PDX samples available for NGS analysis with the MSK-IMPACT platform, including 19 paired samples (Fig. 2B–D). Previously described recurrent alterations in mesothelioma were noted across the patient and PDX samples, including BAP1, NF2, CDKN2A/B, and TP53 [34, 49]. This cohort had fewer BAP1 (26%) and more NF2 (53%) alterations in the patient cohort compared to recently published data from our group using the same NGS platform on 194 patients with DPM (BAP1: 32%, NF2: 25%) [32]. Overall, genomic fidelity in somatic alterations was high across the paired PDX models and patient samples with strong concordance among BAP1 and NF2 alterations as well as less common alterations. However, CDKN2A/B alterations were less concordant. Prior work from our group deriving a novel machine learning algorithm, OncoCast-MPM, as a tool to prognosticate outcomes in patients with DPM using several demographic, histologic, and genomic parameters helped define multifactorial high risk features, including somatic alterations in NF2, and low risk features, such as BAP1 mutations [42]. Likely owing to the limited sample size (high-risk: n = 9; low-risk: n = 10), no discernable OS difference was noted in our patient cohort (Additional file 4: Fig. S6A). The minor differential representation of genomic alterations in our dataset may be due to the limited sample size, confounding variables such as tumor purity, as well as the possibility that differential biology favoring successful implantation of higher-risk subtypes could contribute.
Gene expression changes in mesothelioma PDX models as a function of histologic subtypeWith the establishment of high concordance of genomic, proteomic, and histologic factors between patient samples and PDX models, we shifted our focus to characterization of the PDX models. We conducted RNA sequencing of our PDX models to explore gene expression patterns across several predefined clinicopathologic features (Additional file 5: Table S2). While the currently accepted classification of DPM into epithelioid, biphasic, and sarcomatoid histologies has proven to be both predictive and prognostic [8,9,10,11], there remains substantial variability in individual patient outcomes and response within a given subtype. To compare DPM PDX’s of different subtypes, confirmed by dedicated pathologic review, we applied principal component analysis (PCA) to the top 500 genes ranked by variance over all samples (Fig. 3A, B). Epithelioid PDX models clustered together and were distinct from the non-epithelioid models which did not show tight clustering. Three of four models without histology (unknown subtype due to absence of cores in TMA) clustered close to the epithelioid subtype; the other two tumors clustered closer to the non-epithelioid tumors. LX651 and LX707 which were biphasic and unknown, respectively, were most distinct from the other PDX models. We performed differential gene expression analysis between epithelioid and non-epithelioid models (sarcomatoid and biphasic) and found 364 genes to be significant (adjusted p-value < 0.05, absolute value log2 fold change > 1.5; Fig. 3C). Among the top differentially expressed genes are KIR2DL4, HMKX1, and CXCL9 (Fig. 3C). To further validate the signal for upregulation of HMKX1 and the downregulation of KIR2DL4 and CXCL9 we performed qPCR analysis (Additional file 4: Fig. S6B). We selected three epithelioid (LX282, LX13, LX333), two biphasic (LX175, LX651), and one sarcomatoid (LX707) models for this analysis. qPCR analysis showed a higher mRNA expression of HMKX1 in epithelioid models as compared to non-epithelioid models. Furthermore, epithelioid models had a lower mRNA expression of KIR2DL4 and CXCL9 as compared to non-epithelioid models (Additional file 4: Fig. S6B). Though heterogeneous, the qPCR analysis confirms our observation from RNA sequencing.
Fig. 3
Gene expression changes in mesothelioma PDX models as a function of histologic subtype. A Sample to sample subtype correlation plot using the top 100 highest variance genes. The spearman correlation was used, and the samples were ordered using hierarchical clustering with complete linkage. B Principal component analysis plot showing mesothelioma PDX samples color-coded based on subtype annotation. C Volcano plot showing top differentially expressed genes (DEGs) in epithelioid vs non-epithelioid subtype comparison D Pathway enrichment analyses on the DEGs of the epithelioid vs non-epithelioid subtype comparison
To identify the top differentially expressed genes and activated pathways between the epithelioid and non-epithelioid PDX model subtypes, we next performed gene set enrichment analysis (GSEA). The majority of the top suppressed pathways in the epithelioid tumors were immune signaling-related, including type I interferon alpha and type II interferon gamma signaling (Fig. 3D), TNF alpha signaling via NFκB, IL6 JAK STAT3 signaling, and inflammatory response (Fig. 3D). Other suppressed pathways included DNA repair and G2M checkpoint signaling (Fig. 3D), which has previously been shown to potentially correlate with response to immunotherapy in other cancer types [51, 52]. Few pathways were enriched in epithelioid PDX model subtypes and myogenesis and KRAS signaling down had the highest enrichment scores (Fig. 3D). A similar analysis was performed in the TCGA dataset [34] to compare pathways in our PDX models (Additional file 6: Fig. S7A and S7B). Patients with epithelioid mesothelioma were enriched for known pathways such as (1) EMT signaling, (2) hedgehog signaling, and (3) beta catenin signaling (Additional file 6: Fig. S7B). These pathways were not significantly enriched in our epithelioid PDX models. Similar to our PDX models, epithelioid patient samples in TCGA were suppressed for immune response pathways such as (1) IL2 STAT5 signaling; and (2) inflammatory response.
Gene expression changes in mesothelioma PDX models grouped by platinum doublet and clinical outcomes in the contributing patientsChemotherapy, consisting of a platinum doublet, remains a standard of care option for patients with mesothelioma with an initial response rate of approximately 40% and a relatively wide distribution of duration of benefit among patients [7]. We sought to identify the top genes and pathways altered between PDX models derived from patients with DPM with time from the completion of platinum doublet to progression of < 6 months (n = 10) vs. ≥ 6 months (n = 10) [22,23,24]. Those with < 6 months response to platinum had a PFS of 3.1 months vs. 12.6 months for those with responses ≥ 6 months (Additional file 7: Fig. S8A; p < 0.0001; HR = 5.05). When compared by OS, those with < 6 months of response to platinum had an OS of 13.3 months vs. 25.7 months for those with response ≥ 6 months (Additional file 7: Fig. S8B; p = 0.02; HR = 3.194).
Despite the widespread utilization of platinum doublets in clinical practice, our understanding of the spectrum of differential patient response remains poor. Comprehensive gene expression analyses of DPM PDX models and their implications for patient prognosis have not been reported. We investigated the gene expression landscape of our murine models to evaluate putative gene and pathway associations with clinical outcomes. We compared gene expression and pathway enrichment analyses (GSEA) of PDX samples based on the PFS of the corresponding patient, stratifying based on platinum doublet PFS < or ≥ 6 months (Fig. 4A). Samples clustered based on PFS, with the groups exhibiting distinct gene expression landscapes. PDX histology and OncoCast-MPM risk score did not appear associated with this stratification. Though heterogeneous, the samples with PFS ≥ 6 months had higher expression of some immune activation-related genes, including ITGAL, IL12RB1, IL2RA, and CXCL9.
Fig. 4
Gene expression changes in mesothelioma PDX models grouped by platinum doublet and clinical outcomes in the contributing patients. A Heatmap of gene expression of PDX samples for top genes altered by PFS on platinum doublet (< 6 months vs ≥ 6 months). B Pathway enrichment analyses of the top pathways altered by platinum doublet PFS (< 6 months vs ≥ 6 months) in PDXs. C Heatmap of gene expression of PDX samples for top genes altered between OS (< 2 years vs ≥ 2 years). D Pathway enrichment analyses of the top pathways altered between OS (< 2 years vs ≥ 2 years)
Pathway enrichment analyses based on platinum doublet PFS found that PDXs from patients with PFS < 6 months (Fig. 4B) showed upregulation of genes involved in (1) WNT and β-catenin signaling; (2) hedgehog signaling; and (3) mitotic spindle. GSEA analyses also showed suppression of genes involved in major immune activation pathways, including (1) type II IFNγ signaling, (2) IL6/JAK/STAT3 signaling, (3) type I IFNα signaling, and (4) inflammatory response (Fig. 4B).
We next performed differential gene expression and pathway enrichment analyses (GSEA) of PDX samples based on OS using a cut point of < 2 years vs. ≥ 2 years. A heatmap (Fig. 4C) shows samples clustered based on OS and with the groups showing distinct gene expression landscapes. Though heterogeneous, the samples with OS ≥ 2 years had higher expression of some immune activation-related genes, including GZMB and CD53 (Fig. 4C).
Pathway enrichment analyses performed on genes differentially expressed based on OS (< 2 years vs. ≥ 2 years) found that PDXs from patients with OS < 2 years showed upregulation of genes involved in (1) hedgehog signaling, (2) EMT transition, and (3) Notch signaling (Fig. 4D). As in the platinum PFS analysis, GSEA analyses showed suppression of genes involved in major immune activation pathways in models with OS < 2 years, including (1) type I IFNα signaling, (2) IL6/JAK/STAT3 signaling, (3) type II IFNγ signaling, and (4) inflammatory response (Fig. 4D). To compare our results to patient tissue, we performed a similar analysis in the TCGA dataset [34] (Additional file 8: Fig. S9A and S9B). Similar to our PDX models, patients with OS < 2 years were enriched for (1) EMT transition, (2) Hedgehog signaling, and (3) apical junction. Patients with a shorter OS were similarly suppressed for immune pathways such as (1) type I IFNα signaling and (2) type II IFNγ signaling as well as the (3) oxidative phosphorylation pathway (Additional file 8: Fig. 9B).
Taken together, our data shows PDXs from patients with shorter PFS and OS following platinum doublet treatment exhibit enrichment in pathways involved in WNT/β-catenin, hedgehog, and NOTCH signaling and suppression of pathways involved in immune activation pathways including type I IFNα/β, type II IFNγ, IL6/JAK/STAT3, and inflammatory response.
Gene expression changes in mesothelioma PDX models grouped by platinum/pemetrexed exposure at the time of PDX sample collectionTo further analyze the transcriptional changes in DPM, we performed differential gene expression and pathway enrichment analyses (GSEA) of PDX samples based on platinum/pemetrexed exposure at time of tissue collection. For the top 50 differentially expressed genes, there was not clear clustering by treatment status suggesting the gene expression landscape was not distinct between the groups (Fig. 5A). However, there were a few samples in the treated group which were distinct, namely LX175, LX651, LX13, and LX707. There was no other corresponding clinical feature that clustered with these samples (Fig. 5A).
Fig. 5
Gene expression changes in mesothelioma PDX models grouped by platinum/pemetrexed exposure at the time of PDX sample collection and OncoCast-DPM risk score. A Heatmap of gene
留言 (0)