
記住我
DNAm, a dynamic process that controls genome integrity and transcriptional activity is the most studied epigenetic mechanism. DNAm takes place at the 5-carbon location of the cytosine (C) base, mainly in the CG position (CpG loci), which are unevenly allocated throughout the genome and to a lesser extent in non-CG context. Several DNA methyltransferases (DNMTs) are responsible for ligating methyl groups to DNA. These DNMTs include DNMT 3 beta (DNMT3B), DNMT 3 alpha (DNMT3A), and DNMT1 (Fig. 3). These CpG loci are often found in the promoter regions of genes, and the addition of methyl becomes a steric barrier to transcription factor ligation and gene expression: hypermethylation is often related to transcriptional repression, whereas hypomethylation is related to the activation [6, 8]. The methyl donor is S-adenosylmethionine. There are active demethylation (by translocation enzymes, possibly oxidizing methyl groups to hydroxymethylation, and then repair) and passive demethylation. Furthermore, when methylated cytosine is first oxidized to hydroxymethyl-cytosine by translocation enzymes, targeted passive demethylation also occurs. This state remains until the next S-phase. During this period, hydroxymethyl-cytosine is not identified by DNMT1 and thus changes into an unmethylated cytosine on the newly synthesized strand. Thus, translocases can target cis-regulatory elements to lose methylation at specific loci [19]. Methylation changes in candidate genes are associated with growth, circadian clock regulation, immunity, inflammation, appetite control, metabolism, insulin signaling, and obesity or related phenotypes [8, 9]. Currently, the most commonly used biological samples for DNAm detection are blood samples including whole blood and leukocytes [20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53], adipose tissues [16, 54,55,56,57,58,59], skeletal muscles [60,61,62,63], placenta [64,65,66,67,68] and saliva [69,70,71,72,73] etc. (Fig. 4).
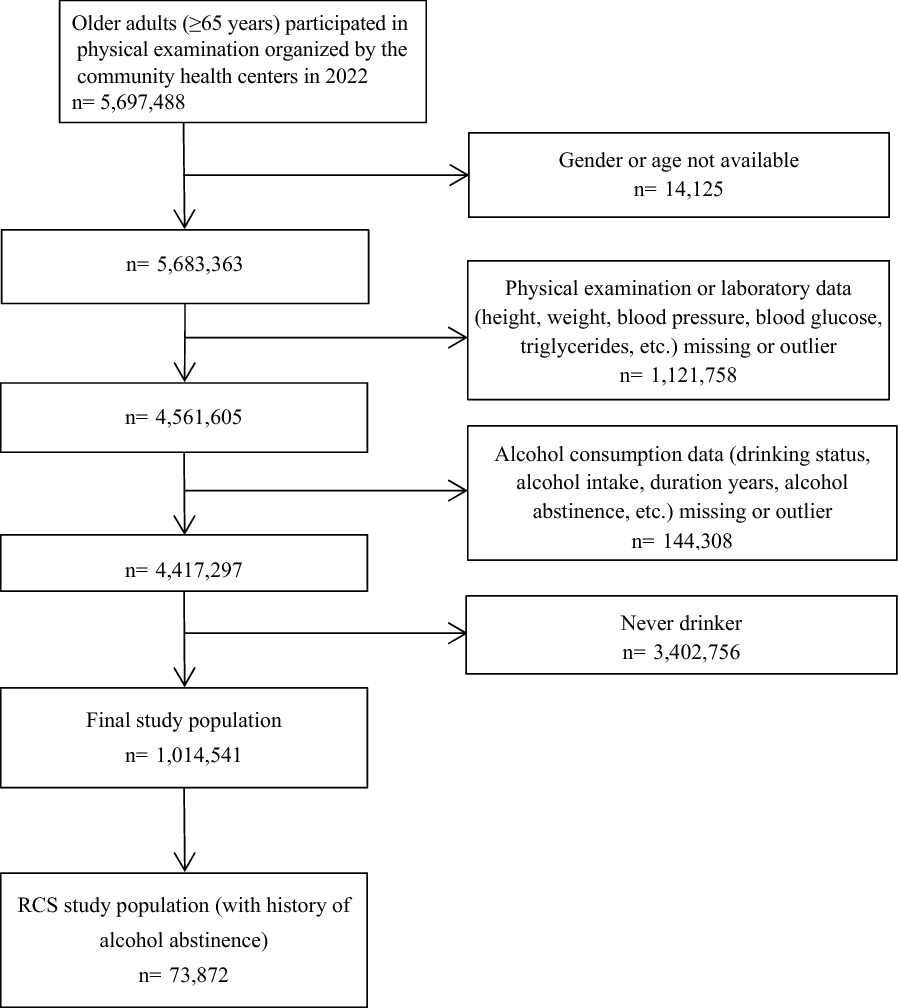
Fig. 3
Diagram showing the mechanism of DNA methylation (DNAm). DNAm often takes place at the 5-carbon location of the cytosine (C) base, mainly in the CG position (CpG loci), which are unevenly allocated throughout the genome and to a lesser extent in non-CG context. Several DNA methyltransferases (DNMTs) are responsible for ligating methyl groups to DNA. The methyl donor is S-adenosylmethionine (SAM). SAM is converted to S-adenosylhomocysteine (SAH)
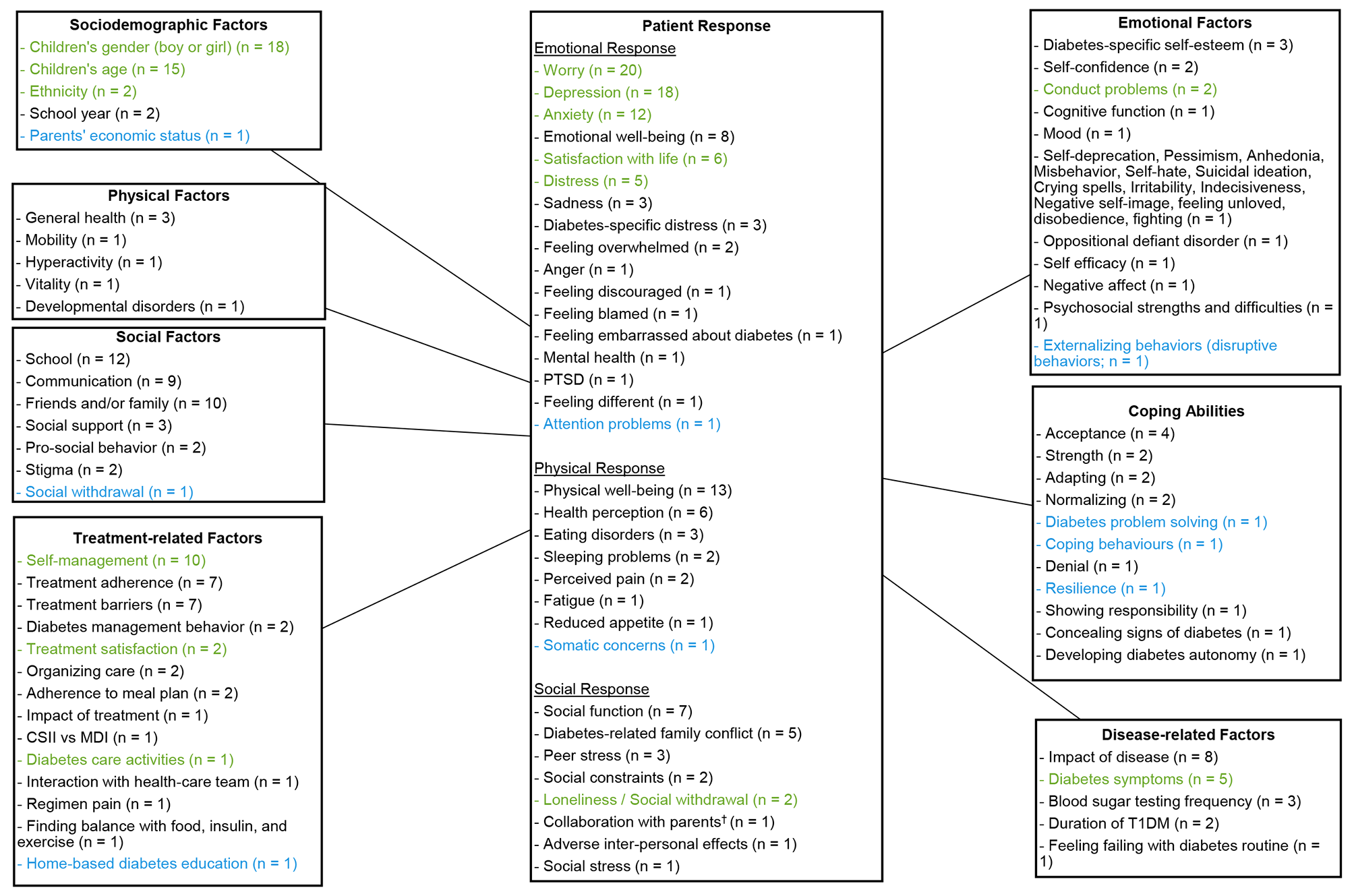
Fig. 4
DNA methylation (DNAm) in different biological specimens of obesity. The figure illustrates tissues and genes with observed alterations in DNAm in subjects with obesity. Some of these genes are also associated with differential gene expression. Changes in DNAm are more often in obese individuals than in non-obese participants. The full name of the abbreviations can be found in the abbreviate table
DNAm in blood samplesDNAm in newbornsMartin et al. [20] identified CpG loci in cord blood leukocytes using data on 173 mother-male and 187 mother-female from the Newborn Epigenetics Study (NEST) cohort. The results showed that maternal pre-pregnancy obesity was associated with 293 CpG loci in male offspring and 876 CpG loci in female offspring (false discovery rate, FDR < 5%). In female offspring, 57 CpG loci, including the top 18, were assigned to tapasin binding protein (TAPBP, methylation changes: -0.83% to 4.02%). CpG methylation differences for TAPBP were also found in male offspring (change: -0.30% to 2.59%), but none of the CpG loci were replicated in the Avon Longitudinal Study of Parents and Children (ALSPAC) cohort. In NEST, differences in the methylation of TAPBP’s CpG loci were related to body mass index z-scores (BMIz, cg23922433 and cg17621507) [20]. This suggests that maternal obesity may have gender-specific differences in the BMIz of offspring (Table 1). Jönsson et al. [21] also explored whether lifestyle interventions in obese pregnant women affect epigenetic variation in umbilical cord blood as well as body composition of offspring. They performed a genome-wide DNAm analysis of 208 offspring from the Treatment of Obesity Pregnant Women study. As compared to controls (standard of care), the offspring of mothers had alterations in DNAm at 379 loci in cord blood, annotated at 370 genes, after a lifestyle intervention such as physical activity with or without dietary recommendations. A total of 370 genes including response to fatty acids and development of adipose tissue were overrepresented in terms of Gene Ontology. The offspring of mothers who receive lifestyle interventions had more lean body mass at birth than the control group. DNAm was found at 17 loci, mainly annotated 4 genes, including the disrupted-in-Schizophrenia-1 (DISC1), gastrulation brain homeobox 2 (GBX2), HECT, UBA and wwe domains containing E3 Ubiquitin ligase 1 (HUWE1), and HECT and RLD domain containing E3 ubiquitin protein ligase 2 (HERC2), partially mediated the effect of lifestyle intervention on offspring lean body mass (FDR < 5%). Furthermore, 22 methylation loci were associated with offspring BMIz within 3 years after birth (P < 0.05) [21]. These findings suggest that lifestyle intervention is associated with epigenetic changes in offspring that can affect offspring lean body mass and early growth.
Table 1 Epigenome-wide DNA methylation association analyses of obesity in different populationsDNAm in childrenHuang et al. [22] studied DNAm profiles in whole blood from 78 obese children and 71 normal children matched for age and sex, respectively. By comparing the methylation profiles between the two groups of children, the investigators identified 129 differentially methylated CpG (DMCpG) loci and the methylation of these genes differed by > 10%. Signaling pathways with high DMCpG enrichment include immune system regulation, cell signaling regulation, developmental processes, and small GTPase-mediated signaling. In individual subjects, sodium bisulfite pyrosequencing was used to verify the association of specific DMCpG methylation with childhood obesity, such as the FYN oncogene related to SRC, FGR, YES; GALE, UDP-galactose-4-epimerase (FYN), Piwi-like RNA-mediated gene silencing 4 (PIWIL4), and thousand-and-one amino acid kinase 3 (TAOK3). In obese individuals, three CpG loci in FYN were hypermethylated, while obesity was significantly associated with hypomethylation of the CpG loci in TAOK3 and PIWIL4. Each 1% increase in TAOK3 methylation was associated with a 0.91-fold decrease in the odds of obesity, while a 1% increase in FYN CpG3 methylation was associated with a 1.03-fold increase in the odds of obesity [22]. These findings provide ample evidence that childhood obesity is associated with the level of specific DNAm in whole blood and that some DNAm may serve as biomarkers of the risk of obesity in the future. Fradin et al. [23] also investigated DNAm profiles of 40 obese children and controls, and identified 31 distinct methylated CpG loci in obese children. The majority of these CpG loci were hypermethylated in obesity. In severely obese children, 151 differentially methylated CpG loci were identified, 10 of which differed by more than 10% methylation. The main pathways enriched at the identified CpG loci include “insulin receptor substrate 1 (IRS1) target genes” and several cancer pathways [23]. This research may contribute to understand obesity and its associated complications. In 374 preschool children, 212, 230, 120, 24 and 15 specific DNAm variants were identified in whole blood samples, and they were associated with body mass index (BMI), fat-mass, fat-free-mass, fat-mass-index and fat-free-mass-index, respectively. These DNAm sites were also significantly associated with probes in genes of non-coding RNAs (ncRNAs) LOC101929268, myeloid/lymphoid or mixed-lineage leukaemia, translocated to 4 (MLLT4), cartilage intermediate layer protein 2 (CILP2), chromosome 3 open reading frame 70 (C3orf70), ST6 beta-galactosamide alpha-2,6-sialyltranferase 1 (ST6GAL1), Zinc finger protein 643 (ZNF643/ZFP69B), SOS Ras/Rac guanine nucleotide exchange factor 1 (SOS1), previously PR domain containing 14 (PRDM14), PR/SET domain 14, CASP8 and FADD like apoptosis regulator (CFLAR), extracellular leucine-rich repeat and fibronectin type III domain containing 2 (CYTH4-ELFN2), cytohesin 4, WD repeat domain 51A/centriolar protein A (WDR51A/POC1A), Kelch like family member 6 (KLHL6) and Sushi, nidogen and EGF like domains/insulin-responsive sequence DNA-binding protein 1 (SNED1/IRE-BP1) after Bonferroni correction [23]. These findings support the potential epigenetic modifications in body composition and obesity [24]. Li et al. [25] identified 226 methylated CpG loci differed between 3 obese children (mean BMI: 21.67) and 4 age/sex matched controls (mean BMI: 14.92). These methylated CpG sites were mainly engaged in immunity and lipoprotein metabolism. Then, the candidate CpG sites within the histone deacetylase 4 (HDAC4), regulator of axillary meristems 2 (RAX2), apolipoprotein A5 (APOA5), carboxylesterase 1 (CES1) and solute carrier family 25, member 20 (SLC25A20) genes were validated using bisulfite sequencing PCR in a cohort of 42 controls and 39 obese cases. The results revealed that methylation levels within the RAX2 and HDAC4 loci were positively associated, whereas those within the CES1 and APOA5 loci were negatively associated with childhood obesity [25]. Thus, changes in the specific CpG loci may contribute to obesity, which may provide a new direction for the etiology of obesity. Sambras et al. [26] analyzed the potential association between DNAm and childhood obesity. DNA samples of 24 children in white blood cells were obtained from the Grupo Navarro de Obesidad Infantil (GENOI) study (obesity and control groups, n = 12; respectively). The association between the microarray data of two CpG loci and obesity were validated in 91 children. A total of 734 CpGs (783 genes) differentially methylated were identified between the two groups. These genes were mainly enriched in circadian and oxidative stress signaling pathways. Furthermore, DNAm levels in several genes such as protein tyrosine phosphatase receptor type S (PTPRS), period3 (PER3), adenylate cyclase activating polypeptide receptor 1 (ADCYAP1R1), N-methyl-D-aspartate receptor subtype 2D (GRIN2D), glutamate receptor, ionotropic, and vasoactive intestinal peptide receptor 2 (VIPR2) were associated with obesity traits. Significant correlations between methylation levels of CpG sites on PTPRS and PER3 with BMIz were also identified by Sequenom MassArray technique from the replication population (n = 91, r = -0.238, P = 0.011 for CpG3 PTPRS, r = -0.198, P = 0.029 for CpG6 PTPRS, and r = -0.280, P = 0.004 for CpG1 PER3) [26].
DNAm in adolescentsObesity is associated with a higher risk of cardio-metabolic diseases even in childhood and adolescence, but it is unknown whether this association is mediated by epigenetic mechanisms. Huang et al. [27] found that mid-childhood BMIz (mean age 7.7 years) was associated with cardio-metabolic risk score in early adolescence (mean age 12.9 years) via mid-childhood DNAm. DNAm in leukocytes was measured among 265 children in the Project Viva. In a high-dimensional mediation analysis and a natural effect model, an association between mid-childhood BMIz and cardio-metabolic risk score in early adolescence was observed at 6 CpG loci (natural effect model: β = 0.04, P = 0.032, accounting for 13% of the total effect). The natural direct effect of BMIz on cardio-metabolic risk score remained significant (β = 0.27, P = 1.1E-25). In addition, there were also 5 CpG sites in the opposite direction from the total effect (natural effect model: β = -0.04, P = 0.02) [27]. Mediation in different directions implies a complex role of DNAm in the association between BMI and cardio-metabolic risk in the childhood and adolescence. In the population-based Penn State Child Cohort follow-up exam, the epigenome-wide single nucleotide resolution of DNAm in the CpG loci and surrounding regions of 263 adolescents was obtained from peripheral leukocytes. Among the 5669 loci related to the BMI percentile with the screening rule of P < 0.05, 28 were identified within genes related to obesity. Obesity-related genes were significantly enriched among 103,466 intragenic loci. Furthermore, increased methylation at one locus within single-minded homolog 1 (SIM1) was significantly related to higher BMI percentile [28]. These data might suggest that DNAm may be related to risk of obesity in adolescents. In peripheral blood DNAm of the leptin gene (LEP) from the Isle of Wight Birth Cohort, 23 CpG loci were obtained in 10 years of age (n = 297) and 16 CpG loci in 18 years of age (n = 305) samples. The duration of total and exclusive breastfeeding was associated with DNAm in 4 LEP CpG loci at 10 years, and not at 18 years. Differential methylation region (DMR) analysis identified 5 significant differentially methylated regions. Breastfeeding duration was associated with LEP methylation and BMI trajectory. One DNAm loci of LEP methylation was associated with an early transient overweight trajectory and the other loci were associated with an early persistent obesity trajectory [29]. Furthermore, a total of 174 candidate CpG loci from whole blood DNAm of 325 subjects were screened, which also included correction loci previously found to be associated with BMI in children and adults. Of these 174 CpG loci, 38 CpG loci in prepubertal DNAm were associated with shifts in BMI status, including 30 CpG loci that were sex-specific [30]. Prepubertal DNAm was associated with changes in BMI status during pubertal development, and these associations may be gender-specific. Vehmeijer et al. [31] performed a meta-analysis of EWAS encompassing 4133 children from 23 studies to explore the association between DNAm in umbilical cord blood and whole blood and BMI in children aged 2 to 18 years. DNAm at three CpG loci (cg05937453, cg25212453, and cg10040131) of different age was associated with Bonferroni-significant BMI. DNAm at 9 additional CpG loci in the cross-sectional childhood model was associated with BMI with significance of FDR. The strength of association between DNAm and adult BMI for the 187 CpG loci previously identified was positively correlated with age in children. Additionally, the correlation coefficients between the estimates of the effect of these CpG loci in adults and children increased. The enrichment of CpG loci was also found to be higher in adults in all age groups [31].
DNAm in youthXu et al. [32] analyzed genome-wide methylation profiles of more than 470,000 CpG loci in peripheral blood samples from obese and lean (48 each) African-American youth aged 14–20 years. A large number of differentially variable CpG loci (DVC) and differentially methylated CpG loci (DMC) were identified. DVC often exhibit abnormal structures and were more variable in cases. Both DVC and DMC determined from the first group independently predicted obesity status in the second group. Furthermore, both DMC and DVC-containing genes showed significant enrichment in genes identified by GWAS for obesity and related diseases (eg, hypertension, dyslipidemia, and T2D) [32]. These findings suggest that different variants are an important feature of obesity-associated methylation. A total of 76 CpG loci associated with obesity were identified in the EWAS study of 700 African Americans aged 14–36 years. Fifty-four of these loci were validated in the Atherosclerosis Risk in Communities (ARIC) study of 2097 African Americans aged 47–70 years, 29 of which were newly associated with obesity. Thirty-seven CpG loci were replicated in neutrophils, and 51 CpG loci were associated with at least one cardiometabolic risk factor. However, the number of CpG was reduced to 9 after adjustment for obesity. Seventeen CpG loci were associated with the expression of 17 genes in the cis, and five of which were expressed at different levels between obese and lean individuals [33].
DNAm in womenWilson et al. [
留言 (0)