
記住我
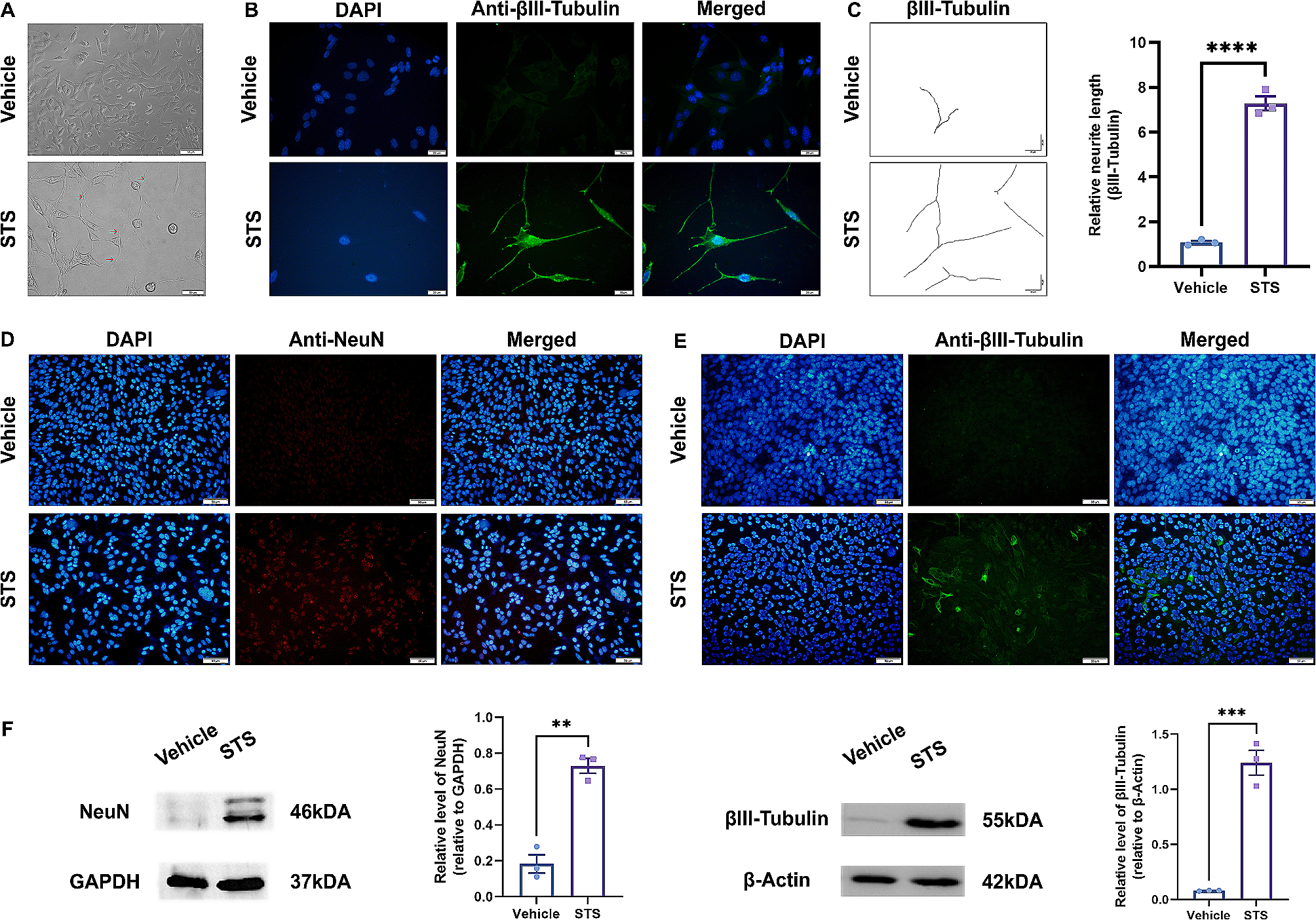



We had previously generated an insulin resistant diabetic neuronal cell model by differentiating N2A cells in the chronic presence of insulin (100 nM) in serum-free medium (MFI) [1, 25, 28,29,30]. In the present study we investigate the expression of PHLPP1 and PHLPP2 in insulin sensitive (MF) and insulin resistant (MFI) neuronal cells. To execute that, insulin sensitive (MF) and insulin resistant (MFI) differentiated N2A cells were stimulated with or without 100 nM insulin for 30 min. Cell lysates were western immunoblotted and probed with either anti-PHLPP1 or anti-PHLPP2 antibody. Under MF condition, insulin stimulation did not affect expression of PHLPP1 (Fig. 1A, lane 1 vs. lane 2; lane 3 vs. lane 4) or PHLPP2 (Fig. 1B, lane 1 vs. lane 2; lane 3 vs. lane 4). However, insulin resistance upregulated expression of PHLPP1 by 4.2 fold (Fig. 1A, lane 1 vs. lane 3) and of PHLPP2 by 2.3 fold (Fig. 1B, lane 1 vs. lane 3) irrespective of insulin stimulation.
Fig. 1
Expression of PHLPP1 and PHLPP2 under insulin resistant condition in neuronal cells. A–B N2A cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Cells were lysed and subjected to western blotting, followed by probing with relevant primary antibodies. A Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. B Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. C–D SHSY-5Y cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Cells were lysed and subjected to western blotting, followed by probing with relevant primary antibodies. C Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. D Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. E–F Mice whole brain was lysed, subjected to western blotting, followed by probing with relevant primary antibodies. E Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. F Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. GAPDH has been used as a loading control. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01 compared to lane 1 IB Immunoblot
To determine whether the above-mentioned effects were cell or species specific, we tested the same in SHSY-5Y cells, a human neuronal cell line. Insulin sensitive (MF) and insulin resistant (MFI) differentiated SHSY-5Y cells displayed results similar to N2A cells (Fig. 1C, D). Expression of both PHLPP1 and PHLPP2 was upregulated under resistant (MFI) condition, however, PHLPP1 expression was more than PHLPP2 in N2A and SHSY-5Y.
Expression of PHLPP1 and PHLPP2 in High Fat Diet (HFD) fed diabetic mice whole brain tissue lysateWe next tested expression of PHLPP isoforms in whole brain tissue lysate of High-Fat-Diet (HFD) fed diabetic mice. Insulin resistance was generated in sixteen weeks old high-fat-diet (HFD) fed Swiss Albino male mice over ten weeks, and whole brain tissues of those mice, were used for our experiments, as previously reported [1]. Glucose tolerance test and insulin tolerance tests were performed; however, perfusion was not done before brain collection. Mice were divided into two groups: normal diet (ND) and high-fat-diet (HFD), each group containing three animals. Levels of triglyceride, cholesterol, serum glutamic-oxaloacetic transaminase (SGOT), serum glutamic-pyruvic transaminase (SPGT), and fasting blood glucose were elevated by 48%, 29%, 50%, 53%, 40%, respectively in HFD mice as compared to ND mice (kind gifts and information from Dr. Mandal, Indian Institute of Technology-Mandi). In the current study our data showed that, in HFD mice whole brain tissue, expression of PHLPP1 was elevated by 4.5 fold (Fig. 1E, lane 1 vs. lane 2) and of PHLPP2 by 2.2 fold (Fig. 1F, lane 1 vs. lane 2), as compared to respective ND mice whole brain tissue, as observed in resistant neuronal cells.
Effect of PHLPP1 and PHLPP2 silencing on Akt isoforms and AS160 in insulin signaling and insulin resistant neuronal cellsTo test the effect of silencing of individual isoforms on insulin signaling, PHLPP1 and PHLPP2 were silenced, and its effect on expression and activation were tested on the downstream substrates, with or without insulin stimulation. Silencing was optimized at 100 nM siRNA for PHLPP1 and PHLPP2 each (Additional files 1, 2: Figs. S1 and S2 respectively). Cells were transfected with 100 nM of either PHLPP1 or PHLPP2 siRNA and then subjected to differentiation under MF or MFI condition, with or without insulin stimulation. Lysates were subjected to western immunoblotting probed with relevant antibodies as and when mentioned with reference to the relevant experiment. PHLPP1 silencing decreased PHLPP1 expression by 70% under MF and MFI conditions (Fig. 2A, lane 1 vs. lane 3, 4; lane 1 vs. lane 7, 8) as compared to scrambled siRNA transfected cells. Consistent with our previous result (Fig. 1A, lane 1 vs. lane 5, 6), expression of PHLPP1 displayed increase under MFI condition, however, silenced PHLPP1 was effective under both conditions tested. PHLPP1 silencing did not affect expression of PHLPP2 under MF or MFI condition (Fig. 2B). Previously, Chen et al. had reported in mice brain, that PHLPP1 knockout did not lead to compensatory increase in PHLPP2 expression [16].
Fig. 2
Effect of PHLPP1 silencing on PHLPP isoforms, Akt isoforms and AS160 in insulin signaling and insulin resistant condition in neuronal cells (N2A). Three days post-proliferation, PHLPP1 was silenced using PHLPP1 specific siRNA. N2A cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Cells were lysed and subjected to western blotting, followed by probing with relevant primary antibodies. A Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. B Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. C Bar represents relative change in pAkt1 (Ser-473) probed with anti-Akt1 antibody. D Bar represents relative change in pAkt2 (Ser-474) probed with anti-Akt2 antibody. E Post-insulin stimulation, lysates were subjected to immunoprecipitation using anti-Akt3 antibody. Bar represents relative change in pAkt (Ser-473) probed with anti-Akt3 antibody. F Bar represents relative change in pAS160 (Ser-588) when probed with anti-AS160 antibody. G Bar represents relative change in pAS160 (Thr-642) when probed with anti-AS160 antibody. GAPDH has been used as a loading control. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01, *P < 0.05 compared to lane 1, ###P < 0.001, #P < 0.05 as compared to lane 2, $$P < 0.01, $$P < 0.05 compared to lane 6. IB Immunoblot; IP Immunoprecipitation
We have previously reported insulin resistance dependent decrease in serine phosphorylation of Akt1, Akt2 and Akt3 in neuronal cells [1].We next proceeded to test the effect of PHLPP1 silencing on Akt isoforms in neuronal insulin signaling and resistance. Post PHLPP1 silencing, there was no change in expression of Akt1 under all the conditions tested (Fig. 2C). Under control conditions (scrambled siRNA transfected) activation of Akt1, as determined by phosphorylation at Ser-473, was down-regulated from MF to MFI by 44% (Fig. 2C, lane 2 vs. lane 6). However, due to PHLPP1 silencing, activation of Akt1 was unaffected under MF and MFI conditions (Fig. 2C, lane 2 vs. lane 4; lane 6 vs. lane 8). Thus, PHLPP1 does not affect phosphorylation or expression of Akt1 in neuronal insulin signaling and resistance. PHLPP1 silencing, caused no change in expression of Akt2 under all the conditions tested (Fig. 2D). However, under all control conditions (scrambled siRNA transfected) activation of Akt2, as determined by phosphorylation at Ser-474, was down-regulated from MF to MFI conditions by 73% (Fig. 2D, lane 2 vs. lane 6). Due to PHLPP1 silencing, activation of Akt2 was up-regulated by insulin stimulation by 32% (Fig. 2D, lane 2 vs. lane 4), and by 87% (Fig. 2D, lane 6 vs. lane 8) under MF and MFI conditions, respectively. Post PHLPP1 silencing, there was no change in expression of Akt3 under all the conditions tested (Fig. 2E). However, under control conditions (scrambled siRNA transfected) activation of Akt3, as determined by phosphorylation at Ser-472, was down-regulated from MF to MFI conditions by 52% (Fig. 2E, lane 2 vs. lane 6). Due to PHLPP1 silencing, activation of Akt3 was up-regulated by insulin stimulation by 32% (Fig. 2E, lane 2 vs. lane 4), and by 46% (Fig. 2E, lane 6 vs. lane 8) under MF and MFI conditions, respectively. Data shows that PHLPP1 specifically regulates phosphorylation of Akt2 and Akt3, without affecting expression in insulin sensitive and resistant conditions. Additionally, PHLPP1 does not regulate phosphorylation or expression of Akt1 in insulin signaling.
We had previously established how Akt isoforms regulate AS160 in neuronal insulin signaling and insulin resistance [1]. All Akt isoforms regulate serine and threonine phosphorylation of AS160, with Akt2 contributing the most, followed by Akt3 and Akt1 under MF and MFI condition [1]. Now, having studied PHLPP1 specific role in regulating Akt isoforms, the next question was whether this regulation extends to AS160. AS160 is a Rab-GAP (GTPase activating protein) involved in glucose transporter 4 (GLUT4) translocation [1]. Post PHLPP1 silencing, there was no change in expression of AS160 under all the conditions tested (Fig. 2F, G). However, under all control conditions (scrambled siRNA transfected) activation of AS160, as determined by phosphorylation at Ser-588 and Thr-642, was down-regulated from MF to MFI conditions by 24% and 37%, respectively (Fig. 2F, G, lane 2 vs. lane 6). Due to PHLPP1 silencing, activation of AS160 at Ser-588 was up-regulated by insulin stimulation by 51% (Fig. 2F, lane 2 vs. lane 4), and by 17% (Fig. 2F, lane 6 vs. lane 8) under MF and MFI conditions, respectively. Similarly, activation of AS160 at Thr-642 was up-regulated by insulin stimulation by 29% (Fig. 2G, lane 2 vs. lane 4), and by 21% (Fig. 2G, lane 6 vs. lane 8) under MF and MFI conditions, respectively. Previously, PHLPP isoforms have been reported to regulate substrate specificity and amplitude of various substrates of Akt isoforms, but AS160 downstream of PHLPP1 has never been studied in any tissue system before. Thus, we report PHLPP1 specific regulation of Akt isoforms which extends to regulation of AS160 serine and threonine phosphorylation, more so, in a neuronal insulin resistant system.
Having observed, as above, it was imperative to study role of PHLPP2 as well. PHLPP2 silencing decreased PHLPP2 expression by 80% under MF and MFI conditions, respectively (Fig. 3A, lane 1 vs. lane 3, 4; and lane 1 vs. lane 5, 6) as compared to scrambled siRNA transfected cells. Consistent with our previous result (Fig. 1B, lane 1 vs. lane 5, 6), expression of PHLPP2 displayed increase under MFI condition, however, silenced PHLPP2 was effective under both conditions tested. PHLPP2 silencing did not affect expression of PHLPP1 under MF or MFI condition (Fig. 3B). This essentially points to independent roles played by PHLPP isoforms without any compensatory upregulation in expression.
Fig. 3
Effect of PHLPP2 silencing on PHLPP isoforms, Akt isoforms and AS160 in insulin signaling and insulin resistant condition in neuronal cells (N2A). Three days post-proliferation, PHLPP2 was silenced using PHLPP2 specific siRNA. N2A cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Cells were lysed and subjected to western blotting, followed by probing with relevant primary antibodies. A Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. B Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. C Bar represents relative change in pAkt1 (Ser-473) probed with anti-Akt1 antibody. D Bar represents relative change in pAkt2 (Ser-474) probed with anti-Akt2 antibody. E Post-insulin stimulation, lysates were subjected to immunoprecipitation using anti-Akt3 antibody. Bar represents relative change in pAkt (Ser-473) probed with anti-Akt3 antibody. F Bar represents relative change in pAS160 (Ser-588) when probed with anti-AS160 antibody. G Bar represents relative change in pAS160 (Thr-642) when probed with anti-AS160 antibody. GAPDH has been used as a loading control. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01, *P < 0.05 compared to lane 1, ###P < 0.001, ##P < 0.01, #P < 0.05 as compared to lane 2, $$P < 0.01 compared to lane 6 IB Immunoblot; IP Immunoprecipitation
PHLPP2 has also been reported to regulate Akt isoforms specifically in cancer cells [2, 3]. However, unlike PHLPP1, which has been studied as an upstream regulator of Akt isoforms in insulin resistant systems [9, 10], data on PHLPP2 is very limited. Post PHLPP2 silencing, we observed no change in expression of Akt1 under all the conditions tested (Fig. 3C). However, PHLPP2 silencing up-regulated activation of Akt1 post insulin stimulation by 37% (Fig. 3C, lane 2 vs. lane 4), and by 54% (Fig. 3C, lane 6 vs. lane 8) under MF and MFI conditions, respectively. There was no change in expression of Akt2 post PHLPP2 silencing under all the conditions tested (Fig. 3D). Interestingly, due to PHLPP2 silencing, activation of Akt2 was unaffected under MF and MFI conditions (Fig. 3D, lane 2 vs. lane 4; lane 6 vs. lane 8). Post PHLPP2 silencing, there was no change in expression of Akt3 under all the conditions tested (Fig. 3E). Due to PHLPP2 silencing, activation of Akt3 was also up-regulated by insulin stimulation by 33% (Fig. 3E, lane 2 vs. lane 4), and by 51% (Fig. 3E, lane 6 vs. lane 8) under MF and MFI conditions, respectively. Data demonstrates that isoforms of PHLPP regulate isoforms of Akt in insulin sensitive and insulin resistant neuronal insulin signaling.
PHLPP2 silencing caused no change in expression of AS160 under all the conditions tested (Fig. 3F, G). However, under all control conditions (scrambled siRNA transfected) activation of AS160 was down-regulated from MF to MFI conditions by 24% and 35%, respectively (Fig. 3F, G, lane 2 vs. lane 6). Due to PHLPP2 silencing, activation of AS160 at Ser-588 was up-regulated by insulin stimulation by 40% (Fig. 3F, lane 2 vs. lane 4), and by 31% (Fig. 3F, lane 6 vs. lane 8) under MF and MFI conditions, respectively. Similarly, activation of AS160 at Thr-642 was up-regulated by insulin stimulation by 18% (Fig. 3G, lane 2 vs. lane 4), and by 25% (Fig. 3G, lane 6 vs. lane 8) under MF and MFI conditions, respectively. AS160 regulation by PHLPP isoforms has never been studied before. Data on the effects of PHLPP1 and PHLPP2 silencing on downstream substrates has been summarized in Additional file 6: Table S1. Data consolidates that two isoforms of PHLPP regulate three isoforms of Akt in insulin sensitive and insulin resistant neuronal insulin signaling.
Effect of PHLPP1 and PHLPP2 over-expression on Akt isoforms and AS160 in insulin signaling and insulin resistant neuronal cellsHaving seen how PHLPP isoforms regulate Akt isoforms and AS160 by PHLPP isoform specific silencing, we proceeded to test the effect of overexpression of PHLPP isoform as well. We transfected each isoform individually in cells and transfected cells were then subjected to MF and MFI conditions, and the effect of over-expression was tested with or without insulin stimulation. Lysates were subjected to western immunoblotting probed with relevant antibodies as and when mentioned with reference to the relevant experiment.
PHLPP1 over-expression increased PHLPP1 expression (Fig. 4A), without affecting PHLPP2 expression (Fig. 4B) under MF and MFI condition respectively, further corroborating that PHLPP1 and PHLPP2 do not show compensatory upregulation in expression. Due to PHLPP1 overexpression, there was no change in expression of Akt1, Akt2 or Akt3 under all the conditions tested (Fig. 4C–E). However, phosphorylation of Akt2 (Fig. 4D) and Akt3 (Fig. 4E) was down-regulated by insulin stimulation under MF and MFI conditions, without any effect on Akt1 phosphorylation (Fig. 4C). As observed in PHLPP1 silencing data, there was no change in expression of AS160 under all the conditions tested (Fig. 4F, G). However, activation of AS160 at Ser-588 and Thr-642 was down-regulated by insulin stimulation under MF and MFI conditions, respectively. The effect was more pronounced under over-expressed condition as compared to silencing in the resistant cells, which may be attributed to elevated endogenous expression of PHLPP1 in inulin resistant neuronal cells. PHLPP2 over-expression increased PHLPP2 expression (Fig. 5A), without affecting PHLPP1 expression (Fig. 5B) under MF and MFI condition, respectively. Post PHLPP2 over-expression, there was no change in expression of Akt1 (Fig. 5C), Akt2 (Fig. 5D) or Akt3 (Fig. 5E) under all the conditions tested. However, phosphorylation of Akt1 (Fig. 5C) and Akt3 (Fig. 5E) was down-regulated by insulin stimulation under MF and MFI conditions, without any effect on Akt2 phosphorylation (Fig. 5D). Post PHLPP2 over-expression, there was no change in expression of AS160 under all the conditions tested (Fig. 5F, G). However, activation of AS160 at Ser-588 and Thr-642 was down-regulated by insulin stimulation under MF and MFI conditions, respectively. This PHLPP1 and PHLPP2 over-expression data in insulin sensitive and resistant N2A cells, and how it affects downstream substrates has been summarized in Additional file 6: Table S2.
Fig. 4
Effect of PHLPP1 over-expression on PHLPP isoforms, Akt isoforms and AS160 in insulin signaling and insulin resistant condition in neuronal cells (N2A). Three days post-proliferation, PHLPP1 was over-expressed using PHLPP1 specific plasmid. N2A cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Cells were lysed and subjected to western blotting, followed by probing with relevant primary antibodies. A Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. B Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. C Bar represents relative change in pAkt1 (Ser-473) probed with anti-Akt1 antibody. D Bar represents relative change in pAkt2 (Ser-474) probed with anti-Akt2 antibody. E Post-insulin stimulation, lysates were subjected to immunoprecipitation using anti-Akt3 antibody. Bar represents relative change in pAkt (Ser-473) probed with anti-Akt3 antibody. F Bar represents relative change in pAS160 (Ser-588) when probed with anti-AS160 antibody. G Bar represents relative change in pAS160 (Thr-642) when probed with anti-AS160 antibody. GAPDH has been used as a loading control. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01, *P < 0.05 compared to lane 1, ###P < 0.001, ##P < 0.01, #P < 0.05 as compared to lane 2, $$$P < 0.001 as compared to lane 6. IB Immunoblot; IP Immunoprecipitation
Fig. 5
Effect of PHLPP2 over-expression on PHLPP isoforms, Akt isoforms and AS160 in insulin signaling and insulin resistant condition in neuronal cells (N2A). Three days post-proliferation, PHLPP2 was over-expressed using PHLPP2 specific plasmid. N2A cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Cells were lysed and subjected to western blotting, followed by probing with relevant primary antibodies. A Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. B Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. C Bar represents relative change in pAkt1 (Ser-473) probed with anti-Akt1 antibody. D Bar represents relative change in pAkt2 (Ser-474) probed with anti-Akt2 antibody. E Post-insulin stimulation, lysates were subjected to immunoprecipitation using anti-Akt3 antibody. Bar represents relative change in pAkt (Ser-473) probed with anti-Akt3 antibody. F Bar represents relative change in pAS160 (Ser-588) when probed with anti-AS160 antibody. G Bar represents relative change in pAS160 (Thr-642) when probed with anti-AS160 antibody. GAPDH has been used as a loading control. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01, *P < 0.05 compared to lane 1, ###P < 0.001, ##P < 0.01, #P < 0.05 as compared to lane 2, $$$P < 0.001, $$P < 0.01 as compared to lane 6. IB Immunoblot; IP Immunoprecipitation
To determine whether the above-mentioned effects were not cell or species specific, we tested the effect of PHLPP1 and PHLPP2 over-expression on expression and phosphorylation of AS160 (Thr-642 and Ser-588 sites), under MF and MFI condition in differentiated SHSY-5Y cells (Additional file 3: Fig. S3). The effects followed the similar trend as in N2A (data has been consolidated in Additional file 6: Table S3). Thus, data shows that PHLPP regulate Akt isoforms affecting AS160 in neuronal insulin signaling and resistance in an isoform specific manner.
Effect of PHLPP1 and PHLPP2 silencing and over-expression on glucose uptake in insulin-resistant neuronal cellsHaving observed PHLPP isoform specific regulation of Akt isoforms and AS160, we sought out to determine the effect of this on neuronal glucose uptake, if any. PHLPP1 or PHLPP2, silenced or over-expressed differentiated N2A cells were stimulated with or without insulin and subjected to glucose uptake assay. PHLPP1 silencing caused an increase of insulin stimulated 2-NBDG uptake by 42% (Fig. 6A, lane 2 vs. lane 4), and 21% (Fig. 6A, lane 6 vs. lane 8) respectively, when compared MF to respective MFI condition. Similarly, PHLPP1 over-expression caused a decrease (Fig. 6C). PHLPP2 silencing caused an increase of 23% (Fig. 6B, lane 2 vs. lane 4), and 12% (Fig. 6B, lane 6 vs. lane 8) respectively, in insulin stimulated 2-NBDG uptake when compared MF to MFI condition. Similarly, PHLPP2 over-expression caused a decrease (Fig. 6D). This PHLPP isoform specific silencing data has been consolidated in Additional file 6: Table S1, and over-expression data in Additional file 6: Table S2. We conducted the same tests in differentiated SHSY-5Y cells and the effects followed a similar trend (Fig. 6E, F). The data has been consolidated in Additional file 6: Table S3. These data reports PHLPP isoform specificity in regulating glucose uptake in neuronal insulin signaling and resistance.
Fig. 6
Effect of PHLPP isoform specific silencing or over-expression on glucose uptake. Three days post-proliferation, PHLPP1 or PHLPP2 was silenced or over-expressed using isoform specific siRNA or over-expression plasmid respectively, as indicated. (A–D) N2A cells or (E–F) SHSY-5Y were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Differentiated cells were serum starved for 2 h, followed by 100 nM insulin for 30 min. Uptake of 2-NBDG was then measured. Bar represents relative change in uptake of 2-NBDG. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01, *P < 0.05 compared to lane 1, ###P < 0.001, ##P < 0.01, #P < 0.05 as compared to lane 2. A.U Arbitrary Units
Effect of insulin on expression and membrane localization of Scribble in insulin signaling and insulin resistant neuronal cells. Insulin-sensitive (MF) and insulin-resistant (MFI) N2A cells were stimulated with or without insulin (100 nM, 30 min), lysed and fractionated into cytoplasmic and membrane fractions [1]. Lysates were subjected to western immunoblotting probed with anti-Scribble antibody. Scribble was found to be localized in the membrane fraction only as compared to the cytoplasmic fraction (Fig. 7A), and the expression was not regulated by insulin stimulation (Fig. 7A). Expression of Caveolin-1 is considered as a plasma membrane specific marker and the degree of purity of membrane fractions [31]. Expression of Caveolin-1 was only observed in membrane fraction as compared to cytoplasm (Fig. 7A–D). Scribble was also observed in the membrane fractions and in similar quantity in MF and MFI and their (trans)location to the membrane was irrespective of insulin stimulation. This was determined by using GAPDH as a loading control, to perform densitometric analyses, because GAPDH is present in both “cytoplasm” and “membrane” [32], as only membrane-specific markers cannot be used for densitometry analysis of “cytoplasm” fractions. Data shows that Scribble is present in N2A cells and localized at the membrane.
Fig. 7
Effect of Scribble silencing on Scribble, membrane localization of PHLPP isoforms, and neuronal glucose uptake under insulin sensitive and insulin resistant condition in neuronal cells (N2A). N2A cells were differentiated in serum-free medium in the absence of (MF) or chronic presence of 100 nM insulin (MFI) for 3 days. Three days post-differentiation, membrane and cytosol fraction were isolated and subjected to western blotting, followed by probing with relevant primary antibodies. A Bar represents relative change in Scribble when probed with anti-Scribble antibody. (B-D) Three days post-proliferation, Scribble was silenced using Scribble specific siRNA. N2A cells were differentiated in serum-free medium in the absence of insulin (MF) for 3 days. Three days post-differentiation, membrane and cytosol fraction were isolated and subjected to western blotting, followed by probing with relevant primary antibodies. B Bar represents relative change in Scribble when probed with anti-Scribble antibody. C Bar represents relative change in PHLPP1 probed with anti-PHLPP1 antibody. D Bar represents relative change in PHLPP2 probed with anti-PHLPP2 antibody. E Three days post-proliferation, N2A cells were transfected with Scribble specific siRNA and then differentiated under MF MFI condition for 3 days. Differentiated N2A cells were serum starved for 2 h, followed by 100 nM insulin for 30 min. Uptake of 2-NBDG was then measured. Experiments were executed three times and a representative result is shown. Data expressed are mean ± SE. ***P < 0.001, **P < 0.01, *P < 0.05 compared to lane 1, ###P < 0.001, ##P < 0.01, #P < 0.05 as compared to lane 2. IB Immunoblot; A.U Arbitrary Units
Effect of Scribble silencing on membrane translocation of PHLPP1 and PHLPP2Scribble was silenced and its effect on membrane localization was tested on PHLPP1 and PHLPP2, with or without insulin stimulation. 100 nM siRNA was required to silence 70% of Scribble (Additional file 4: Fig. S4). Cells were lysed and fractionated into cytoplasmic and membrane fractions, and lysates were subjected to western immunoblotting probed with anti-Scribble, anti-PHLPP1, anti-PHLPP2 antibody, as and when indicated.
Consistent with the previous result (Fig. 7A), insulin stimulation did not affect Scribble expression, and Scribble was found to be localized on the membrane only as compared to the cytoplasm. Scribble silencing decreased its expression by 70% (Fig. 7B, lane 3 vs. lane 7, lane 4 vs. lane 8) in the membrane fraction. Under control (scrambled siRNA transfected) condition, PHLPP1 was localized on the membrane only (Fig. 7C, lane 3, 4). As a result of Scribble silencing, PHLPP1 translocated from the membrane to the cytoplasm by 60% (Fig. 7C, lane 5 vs. lane 7, lane 6 vs. lane 8). Similarly, under control (scrambled siRNA transfected) condition, PHLPP2 was also localized on the membrane only, with limited expression in the cytoplasm (Fig. 7D lane 3, 4). Scribble silencing caused PHLPP2 translocation from the membrane to the cytoplasm by 50% (Fig. 7D, lane 5 vs. lane 7, lane 6 vs. lane 8). Data shows Scribble determines cellular localization of both PHLPP1 and PHLPP2. This is in coherence with previous study by Li et al., where they reported that in colorectal cancer cells Scribble silencing affected membrane localization of PHLPP isoforms, affecting PHLPP1 more than PHLPP2 [18].
Effect of Scribble silencing on neuronal glucose uptake in insulin signaling and insulin resistant neuronal cellsScribble has previously been reported to play varied roles as a tumor suppressor [29,
留言 (0)