
記住我
Mammalian p38 MAPK is crucial for immunity and inflammatory responses [23, 24]. PMK-1, a C. elegans ortholog of p38 MAPK, has been established as a key regulator of innate immunity, which upregulates transcription factors, including ATF-7/activating transcription factor 7 and SKN-1/NRF [12, 17, 20]. Therefore, investigating the role of PMK-1/p38 MAPK in immunosenescence was an intriguing and vital question to address.
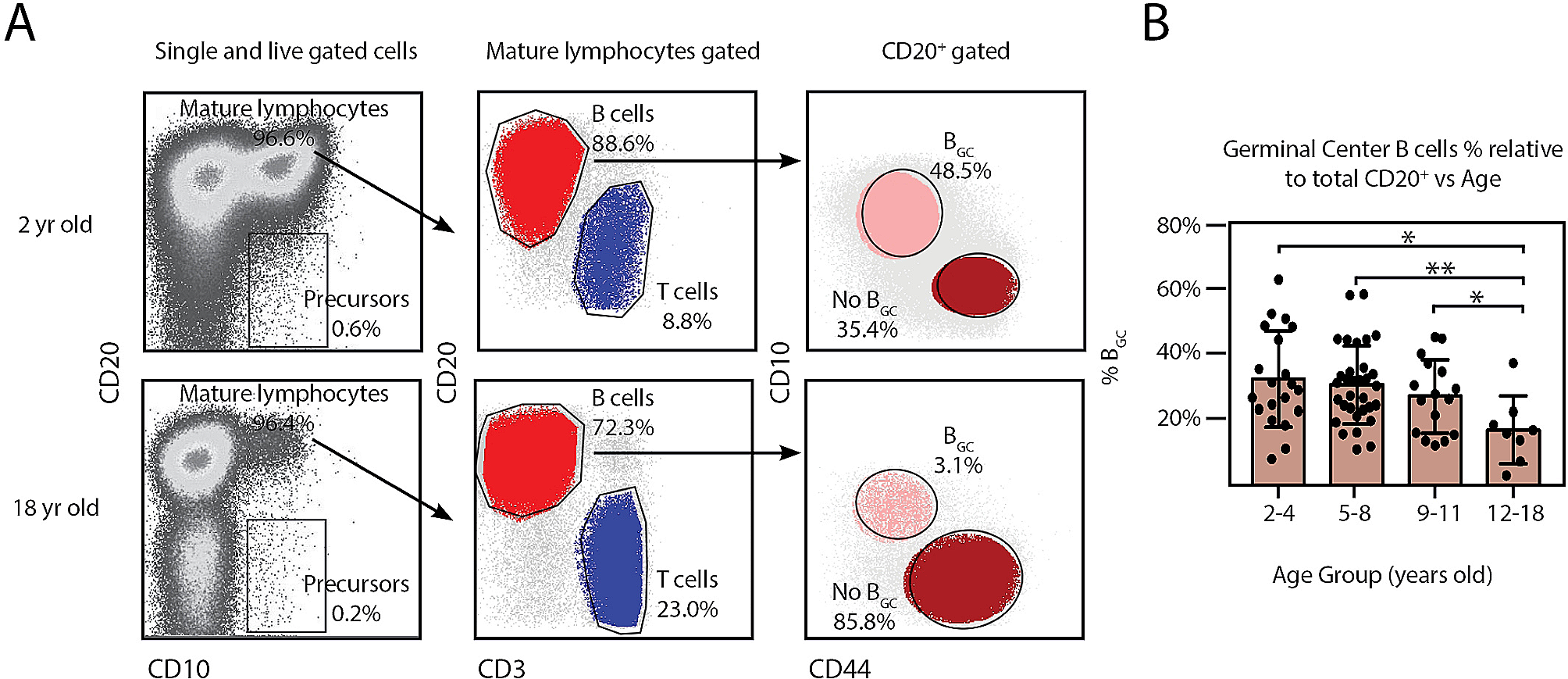
Youngman et al. reported that the age-dependent decline of PMK-1 activity underlies immunosenescence by accelerating the deterioration of the intestine, a main immune organ in C. elegans [16]. The authors showed that PMK-1/p38 MAPK is downregulated during aging at the transcriptional and translational levels. The mRNA levels of PMK-1/p38 MAPK targets decrease with age. The authors then performed survival assays by infecting animals with P. aeruginosa PA14 strain at the juvenile L4 larval stage, and day 3 (young), day 6 (middle age), and day 9 (old) of adulthoods, and determined the role of PMK-1/p38 MAPK in immunosenescence. The effects of pmk-1 mutations on impaired pathogen resistance decrease in aged animals depleted of PMK-1. Therefore, the authors concluded that the reduced levels of PMK-1/p38 MAPK in aged C. elegans contribute to age-dependent impairment of immune responses (Fig. 1A). The authors then showed that pmk-1 mutations induce the accumulation of bacteria in the intestine, leading to intestinal damage. In conclusion, gradual downregulation of PMK-1/p38 MAPK promotes intestinal aging and impairs immune function, accumulating pathogenic bacteria in the gut, which induces damage in the intestine. This study highlights the role of PMK-1/p38 MAPK, which protects animals from intestinal tissue damage in young age, and its age-dependent downregulation leads to immunosenescence via a vicious circle of damage and pathogen infection in the intestine.
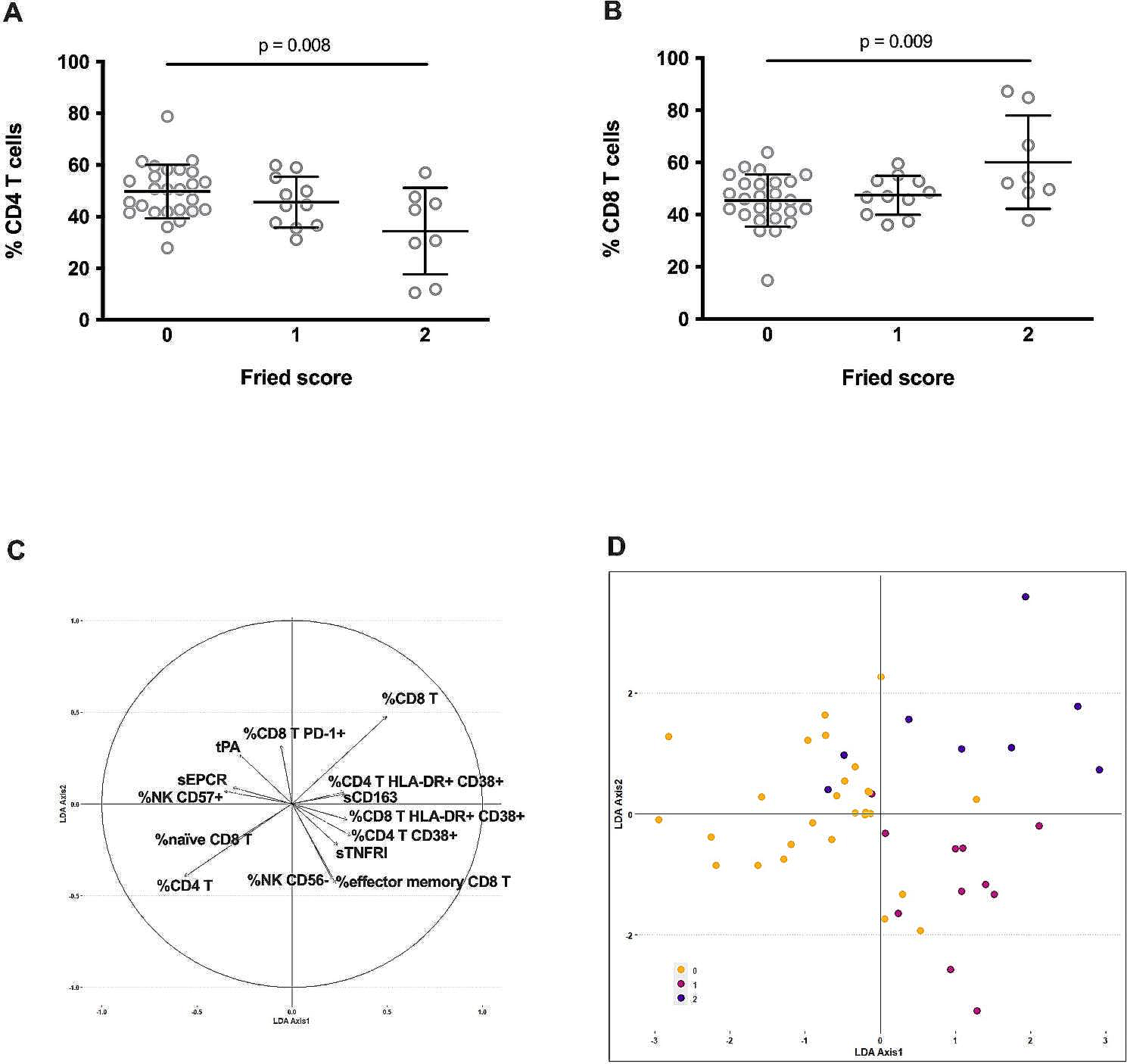
Fig. 1
Factors and pathways that regulate immunosenescence in Caenorhabditis elegans. A Upon pathogen infection of aged wild-type C. elegans, downregulation of PMK-1/p38 mitogen-activated protein kinase (MAPK) leads to intestinal damage and immunosenescence. In addition, downregulation of SKN-1/NRF transcription factor decreases the expression of stress-responsive genes and contributes to immunosenescence. B Aged daf-2 mutant C. elegans displays reversed immunosenescence. Genetic inhibition of DAF-2/insulin/IGF-1 receptor upregulates DAF-16/FOXO and HSF-1/heat-shock transcription factor 1. This leads to downregulation of INS-7/insulin-like peptide, which is also regulated by the ZIP-10/bZIP transcription factor, resulting in further reduction of DAF-2 signaling, and reverses immunosenescence
SKN-1/NRF prevents immunosenescence in young C. elegansThe bacterial virulence factors, such as bacterial toxins, often induce damage in the host intestine by counteracting the host antimicrobial defense mechanisms, and several stress regulators, including the components in IIS, regulate immunity [25, 26]. The transcription factor SKN-1/NRF, which regulates oxidative and xenobiotic stress responses, longevity, and immunity, acts in both p38 MAPK and IIS pathways [14, 21, 27]. However, the role of SKN-1/NRF in immunosenescence was unknown.
Papp et al. reported that SKN-1/NRF is required for normal immunity against gram-negative P. aeruginosa PA14 and gram-positive Enterococcus faecalis [17]. PA14 infection increases the nuclear localization of SKN-1/NRF through Toll/IL-1 Receptor domain protein/TIR-1 and PMK-1/p38 MAPK and subsequently increases the expression of SKN-1/NRF target genes, γ glutamylcysteine synthetase 1 (gcs-1) and glutathione S-transferase 4 (gst-4). They also showed an age-dependent decrease in the expression of SKN-1/NRF target genes upon PA14 infection. The authors experimentally evaluated the relationship between SKN-1/NRF activity and immunosenescence and found that SKN-1/NRF target gene expression decreases with age (Fig. 1A). In addition, the survival of day 4 wild-type animals on PA14 is similar to that of day 1 skn-1 mutant animals. These results indicate that a decline in SKN-1/NRF function contributes to immunosenescence. Furthermore, SKN-1/NRF is required to enhance pathogen resistance mediated by reduced IIS or oxidative preconditioning. However, the hyperactivation of SKN-1/NRF by the genetic inhibition of WDR-23, a direct repressor of SKN-1, impairs pathogen resistance. Thus, the optimal activity of SKN-1/NRF appears crucial for maintaining proper immune responses, consistent with our recent study on the proper regulation of SKN-1/NRF for enhanced immunity and healthy longevity [28]. Overall, this study identifies SKN-1/NRF as a critical factor that contributes to preventing premature immunosenescence in C. elegans.
Reduction of IIS can reverse immunosenescence in C. elegans through a positive feedback loopIncreased innate immunity at a young age is a general feature in various long-lived C. elegans mutants [13, 14, 25, 29, 30, 31]. Thus, interventions that alter longevity may have causal effects on immunosenescence, but whether and how these genetic factors affect immunosenescence remained elusive.
Our study showed that DAF-2/insulin/IGF-1 receptor contributes to immunosenescence by inducing INS-7/insulin-like peptide 7 through DAF-16/FOXO, HSF-1/heat-shock transcription factor 1, and ZIP-10/bZIP transcription factors [19]. We aimed to identify longevity-promoting interventions that modulate immunosenescence by using five long-lived mutants: insulin/IGF-1 receptor-defective daf-2, sensory neuron-compromised osm-5, dietary restricted eat-2, mitochondrial respiration-impaired isp-1, and germline-deficient glp-1 mutants. Surprisingly, we found that day 9 (old) daf-2 mutants displayed enhanced pathogen resistance against PA14 compared to day 1 (young) daf-2 mutants that are more resistant to PA14 than wild-type animals. The effect is, at least in part, independent of PA14 intake or clearance. These results indicate that genetic inhibition of DAF-2, which substantially increases lifespan, stress resistance and immunity [13, 14, 32, 33, 34], can cause immune rejuvenation (Fig. 1 B).
DAF-16/FOXO, SKN-1/NRF, and HSF-1 are key transcription factors acting downstream of the DAF-2/insulin/IGF-1 receptor [13, 14], and are crucial for longevity and immunity [17, 25, 35, 36]. We showed that DAF-16/FOXO and HSF-1 are required for enhanced immunocompetence in old daf-2 mutants compared with young daf-2 mutants, whereas SKN-1/NRF is not essential. We also found that depletion of PMK-1/p38 MAPK, a major immune-regulatory factor whose downregulation contributes to immunosenescence [16], is not required for the further enhanced immunocompetence of aged daf-2 mutants. Thus, DAF-16/FOXO and HSF-1 are key factors that mediate immune rejuvenation in daf-2 mutants.
We searched for genes that mediate enhanced immunocompetence in old daf-2 mutants by analyzing RNA-seq data using day 1 (young) and 9 (old) adult wild-type and daf-2 mutant animals. We found that ins-7, an agonistic insulin-like peptide [37, 38, 39], is downregulated during aging in daf-2 mutants by activating DAF-16/FOXO and HSF-1. Mutations in ins-7 are sufficient to enhance pathogen resistance in old animals. We concluded that the downregulation of ins-7 contributes to delayed immunosenescence in daf-2 mutants.
We then focused on the ZIP-10/bZIP transcription factor, whose expression increases with age in wild-type animals but decreases in daf-2 mutants, similar to that of ins-7. ZIP-10/bZIP transcription factor plays a role in the defense response against pathogens in plants [40]. We showed that genetic inhibition of ZIP-10 suppresses age-dependent upregulation of ins-7 and increases PA14 resistance in day 9 adult animals. Therefore, downregulation of ZIP-10/bZIP transcription factor in aged daf-2 mutants prevents immunosenescence by increasing the activities of DAF-16/FOXO and HSF-1 by decreasing the levels of INS-7 (Fig. 1 B). This study provides clues for reversing immunosenescence by revealing the role of an immunity-regulating positive feedback loop in IIS in aged C. elegans.
留言 (0)