Toxoplasmosis, a widespread zoonotic infection, is caused by a pathogen within the phylum Apicomplexa named Toxoplasma gondii (T. gondii) in all warm-blooded animals, including about 30% of the human population worldwide (Dubey, 2008). Even cold-blooded animals can serve as a source of infection for humans and other hosts that feed on them (Nayeri et al., 2021). T. gondii can cause a serious infectious disease during pregnancy and may lead to abortion or severe congenital defects, such as intellectual disability, blindness, and hydrocephaly, in the affected fetus (Elsheikha, 2008). In addition, this infection in immunocompromised patients may lead to severe and progressive complications, such as encephalitis or pneumonitis, and even may lead to death if left untreated (Wang et al., 2017). Sulfadiazine/pyrimethamine as available chemical treatments lead to multiple side effects (e.g., fatal bone marrow suppression, hematologic toxicity, and severe allergic reactions) and cannot eliminate the cyst form of the parasite (McAuley et al., 1994; Montoya and Liesenfeld, 2004). Therefore, studies should focus on vaccine production for the prevention and control of T. gondii infection. Over recent decades, numerous researchers have examined the immunogenicity of T. gondii antigens, including surface antigens (SAGs), micronemal antigens (MICs), dense granule antigens (GRAs), rhoptry antigens (ROPs), and other antigens involved in motility, adhesion, proliferation, and invasion in a parasitophorous vacuole (PV) (Dodangeh et al., 2019; Faridnia et al., 2018; Hajissa et al., 2019; Pagheh et al., 2020; Rezaei et al., 2019). Among these, SAG1/P30, a protein with an extreme antigenicity and immunogenicity, is a major surface antigen for tachyzoite (Kasper and Khan, 1993) that interacts with host cells and is involved in adhesion, signal transduction, material transport, and cell invasion; moreover, it is able to induce both specific humoral and cellular immunity against T. gondii infection (Pagheh et al., 2020). The MIC4 protein is secreted at different stages of the parasite life cycle (i.e., the merozoites, sporozoites, bradyzoites, and tachyzoites) and has received much attention due to its effective binding to host cells (Wang et al., 2009). Furthermore, the ROP18 protein is only detectable in the tachyzoite stage, and this antigen is released during parasite invasion of host cells, inhibiting the accumulation of immune-related GTPases (IRGs) on PV; moreover, it is involved in the pathogenesis (Jones et al., 2017; El Hajj et al., 2007; Saeij et al., 2006; Taylor et al., 2006). Several studies have been carried out on ROP18, MIC4, and SAG1 as potential vaccine candidates (Dodangeh et al., 2021; Letscher-Bru et al., 2003; Solhjoo et al., 2007; Pinzan et al., 2015; Wang et al., 2011; Yuan et al., 2011). The DNA vaccine of MIC4 has been displayed to elicit immunity in a murine model against toxoplasmosis (Peng et al., 2010). The SAG1-encoded DNA vaccine, in combination with aluminum phosphate as an adjuvant, induces a humoral immune response in mice against Toxoplasma infection (Solhjoo et al., 2007). The MIC4, along with SAG1, has been revealed to induce a humoral and cellular immune response in vaccinated mice (Wang et al., 2009). The DNA vaccine combining ROP18 with MIC3 triggers stronger humoral and Th1-type cellular immune responses in immunized mice (Qu et al., 2013). A multi-epitope vaccine containing SAG1, ROP8, and MIC3 demonstrated a stronger Th1 immune response and increased survival time in vaccinated mice, compared to controls (Dodangeh et al., 2021). A combination of three antigens (ROP18, MIC4, and SAG1) was used for the first time in the present study. On the other hand, intracellular parasites, such as T. gondii, have a large number of antigens and antigenic epitopes. The most immunogenic epitopes of one antigen were selected using the in silico method, in combination with the most immunogenic epitopes of the other two antigens, and this is another novelty of this study. Furthermore, in this study, three adjuvants [Freund adjuvant, calcium phosphate nanoparticles (CaPNs), and chitosan nanoparticles (CNs)] were used. Adjuvants enhance the immunogenicity of weaker immunogens, improve vaccine efficacy, and affect the nature of the immune response, directing it to the cell or humoral responses (Reed et al., 2013). In recent decades, various adjuvants, such as vaccine delivery systems (e.g., microparticles, emulsions, and liposomes) and immune-stimulating adjuvants (e.g., monophosphoryl lipid A, lipopolysaccharide, and CpG DNA), have been used in new vaccine formulations (Li and Zhou, 2018). Moreover, genetic adjuvants, the adjuvant ginseng stem-and-leaf saponins, and T. gondii profilin-like protein triggered a strong immune response to the vaccine (Cao et al., 2015; Hedhli et al., 2016; Zhuo et al., 2017). Finally, this research evaluated the immunogenicity of RMS as a vaccine candidate with or without adjuvants (Freund, CaPNs, and CNs) by subcutaneous immunization of BALB/c mice against acute toxoplasmosis.

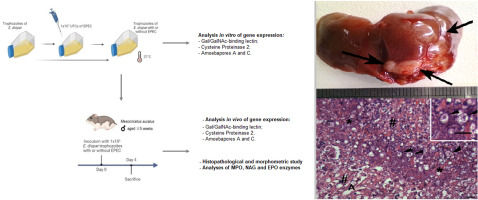
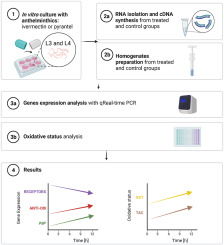


留言 (0)