
記住我





What can be said with certainty about the brain is that this is a complex dynamical system: (i) governed by the deterministic laws of nature at the physical (causal) or “hard” level which (ii) implements cognitive processing at the computational (unconscious) or “soft” level while (iii) its conscious manifestations occur at the phenomenal (mental) or “psyche” level. These three levels are also separated across different spatial-temporal scales. Neural activity is presented at the microscale of cellular interactions, cognitive processing occurs at the mesoscale of neural populations, and conscious experience emerges only at the macroscale of brain dynamics (Varela, 1995; Revonsuo and Newman, 1999). Another way to distinguish between the hard and the soft level in terms of network neuroscience is to relate the former to structural (anatomical) connectivity of hard-wired neurons, where causation really occurs. In contrast, the soft level corresponds to functional connectivity, where distant correlations between brain regions take place. Computations at the soft level cannot violate causal interactions at the hard level but are merely imposed upon physically interacting neurons. They cannot change brain dynamics that obey deterministic laws.
Thus, the mind-brain duality can be viewed in the formal terms of “hard/soft” parallelism between causation and computation. While computation is typically defined through symbolic manipulations of information, its processing depends ultimately on causal chains between inputs and outputs. The question of how those symbolic manipulations translate into conscious experience in the brain and why other information-processing systems such as computers and AI systems lack conscious experience at the psyche level is one of the most mysterious problems in neuroscience.
The main postulate of Gestalt psychology is that conscious experience is a unified whole, which is greater than the sum of its parts. More generally, this postulate is known in the context of the spontaneous emergence of unexpected higher-level phenomena that are not reducible to their low-level constituents (Bedau and Humphreys, 2008; Gibb et al., 2019). A related issue in neuroscience is specified as the binding problem: how brain regions composed of billions of (unconscious) neurons can generate a unified conscious experience at a given moment of time (Edelman, 2003). In fact, many (if not all) theories of consciousness originate from or, at least, can be reduced to how they decide this problem. Among the candidates proposed by known theories are integrated information (II) of irreducible causal mechanisms (Tononi, 2008), global workspace (GW) for broadcasting (Baars et al., 2013; Mashour et al., 2020), updating of priors (UP) in predictive processing (Knill and Pouget, 2004; Clark, 2013), meta-representation (MR) by recurrent processing (Lamme, 2006; Rosenthal, 2008), self-organized criticality (SOC) in brain dynamics (Werner, 2009; Kozma and Freeman, 2017), adaptive resonance (AR) of brain structures (Grossberg, 2017; Hunt and Schooler, 2019), and even large-scale quantum entanglement with a consequent collapse (Hameroff and Penrose, 2014; Fisher, 2015).
Those in turn can be grouped by the similarity of mechanisms or processes involved: II + GW by integration-differentiation processes, GW + UP + MR by feedback mechanisms, SOC + AR by spontaneous synchronization and phase transition. However, any grouping is somewhat arbitrary as the underlying mechanisms can converge to a thermodynamic account: the neural binding arises when brain activity is balanced on the edge between order and disorder. Broadly speaking, consciousness emerges in a very special state of matter somewhere between a perfect crystal and an ideal gas (Tegmark, 2015).
The cognitive evolution theory or CET (Yurchenko, 2022), outlined here, adopts the SOC approach for its apparent advantages over the above models in studying consciousness. SOC is neurophysiologically reliable in resolving the binding problem without resorting to exotic physics or mysterious mind-matter dualism. This provides rich mathematical formalisms applicable to brain dynamics. SOC also proposes the avenue for explaining universal dynamical capacities of the brain to account for large-scale emergent phenomena without involving the so-called downward or top-down causation that might make consciousness like a homunculus due to “synergistic emergence” (Lau, 2009; Hoel et al., 2013; Mediano et al., 2022). On the other hand, SOC is abundantly presented in nature (Bak, 1996; Jensen, 1998; Haken, 2004). However, we do not normally assume that an arbitrary physical system exhibiting critical signatures is conscious. Something else must be inherent to a system to generate consciousness.
CET starts from the obvious fact that the only place where consciousness certainly resides is the brain. There are four principled features that make the brain distinct from all other critical systems. First, the brain consists of neurons specialized for transmitting information over spike patterns. The neurons had evolved from autonomous biological cells possessing all properties of life not merely as mechanistic binary devices. Hence, consciousness is a property of living systems. Second, there are arousal mechanisms regulating sleep-wake cycles in these living systems which can be suppressed by anesthetics. It is also known that damage to arousal nuclei causes immediate coma when the rest of the brain can remain intact (Parvizi and Damasio, 2001; Giacino et al., 2014). Hence, consciousness is impossible without involving special neural nuclei in the brain responsible for arousal. Third, the brain learns and accumulates knowledge. Hence, the brain is a cognitively evolving system. However, AI systems can learn and even cognitively evolve without any kind of awareness. Is there something else that is inherent to the conscious brain but absent in unconscious machines?
The fourth and ultimate distinction is volition, the ability to make free decisions not causally predetermined from the past. This valuable property is thought to be intrinsic to many (if not all) brain systems regardless of their conscious features. In contrast, we do not normally grant volition to computers and AI systems even if they can sometimes surpass humans in cognitive performance. The volition of this kind is akin to one that can be ascribed to clockwork’s engine, i.e., it is an ordinary physical process carrying energy out of one place into another. How much our intuition is right by assuming that consciousness and volition are evolutionarily linked?
With the advent of causation neuroscience, the detailed relationship between statistical models of neural activity and actual causation in the brain is intensively debated (e.g., Albantakis et al., 2019; Reid et al., 2019; Weichwald and Peters, 2021). Statistical measures such as Granger Causality (Granger, 1969) or Entropy Transfer (Schreiber, 2000) had been suggested to infer some aspect of causal interaction among neural entities that are then modeled on a particular structural time-directed graph over a set of nodes. Their definition is based entirely on the predictability of some time-series. If Y contains information that helps to predict X beyond the degree to which X already predicts its own future, then Y is said to have a causal influence on X. Causal modeling must reflect dynamic processes irrelevant to the question of whether a system of interest processes information or not. In cognitive neuroscience, causal inference is based on a synthesis of functional and effective connectivity extracted from neuroimaging data (Friston, 1994). The “gold standard” to establishing whether a stimulus variable Y affects the target variable X is to introduce controlled perturbations to the brain.
It must be however emphasized that causality measures do not necessarily reflect physical causal chains (Seth, 2008). Meanwhile, brain dynamics are commonly believed to evolve completely in causal ways over conscious and unconscious volitional repertoires of the brain. Within those volitional repertoires, the ability generally labeled “free will” is associated with the sum of executive functions, self-control, decision-making, and long-term planning. Thus, it makes free will inseparable from the biological function of consciousness, evolution-driven, and implicitly active (Feinberg and Mallatt, 2020). Putting the above question differently, could consciousness supervene on its own physical substrate to choose the course of action free of predetermination from the past?
To answer this question, a unified theory of consciousness should account for brain activity at three hierarchical levels: (i) at the causal (hard) level; (ii) at the computational (soft) level; and (iii) at the phenomenal (psyche) level. The first two levels should explain how subjective experience and self-awareness emerge from the underlying brain dynamics over which cognitive processing is carried out.
Consciousness and Volition in Brain DynamicsThe stochastic account of free volition can be often found in the literature. For example, Rolls (2012) suggests: “in so far as the brain operates with some degree of randomness due to the statistical fluctuations produced by the random spiking times of neurons, brain function is to some extent non-deterministic, as defined in terms of these statistical fluctuations.” Since the brain contains billions of neurons, causal processes can be only estimated with the help of network statistics extracted from different neuroimaging data. However, probabilistic (counterfactual) descriptions, as those derived from causality measures, reflect the state of our knowledge (ignorance) that, by itself, does not violate determinism. Brain dynamics can still be completely deterministic, i.e., predetermined.
Many free-will advocates usually suggest that if even conscious volition cannot violate determinism at the causal level, it can still be involved in long-term planning of actions at the psyche level. They argue that the ability to use optimal algorithms in predictive processing would be a much more important factor than whether the brain operates deterministically or not (Rolls, 2020). Does it mean that by using those algorithms AI systems might suddenly acquire free volition? Relevantly, relying on this pure computational aspect of volition in the context of the hard-soft duality would imply no obstacle in creating machine (hence, copyable) consciousness at the psyche level (Dehaene et al., 2017; VanRullen and Kanai, 2021). This comes from the observation that there can be no operational difference between a perfect computational simulation of, for instance, Alice’s actions and an in silico copy of her consciousness running automatically on many digital clones (Aaronson, 2016). Thus, ignoring the hard-soft duality also entails the problem of the privacy of consciousness: the clones should know what it is like to be Alice.
A typical scenario suggested for manifesting free volition at the computational (soft) level is one where Alice can consciously plan something ahead of time, for example, visiting her friends tonight. Thus, Alice’s freedom to choose can be proven by achieving her goal. Upon a closer examination, all such scenarios are behavior-driven, yet based on uncertain and rough assumptions about what occurs at the microscale of neural interactions. Within a rigorous physical framework, the spatial and temporal locations of action should be specified via the stream of conscious states, each processed by the brain at the hard level.
Suppose Alice plans to visit her friends at the moment t0 when her consciousness is in a state X. Whenever she could reach this goal, her conscious state at that exact moment t would be Y. But the state Y should in turn have been consistently processed from the previous state Y − 1 over ubiquitous causal chains. How could it be done freely? Moreover, the manifestation of the conscious will should be related not to Alice being at her goal state Y but to its mental initiation in X. Indeed, after the decision has been made, her goal-directed behavior could be completely deterministic. However, this state should also be causally processed from the previous state X − 1, and so on. How could mental initiation be free of the past? Hence, if consciousness cannot choose the next state from a given past state, no future state in the stream can be chosen at all.
On the other hand, if the brain cannot make a choice free of the past, the old-fashioned fatalism, also known as superdeterminism in the context of quantum mechanics (‘t Hooft, 2016; Hossenfelder and Palmer, 2020), would prevail. This states that neither consciousness nor even the brain might violate deterministic laws to do otherwise than what has been predetermined by the past. How could free volition be reconciled with computational models of consciousness at the hard level?
Volition in Theories of ConsciousnessConsciousness and volition are intrinsically linked, and both are largely ignored in neuroscience. Although there are now a plethora of various theories of consciousness, the free will problem still remains largely neglected. The theories do not explain how the brain integrates consciousness (psyche), cognition (soft), and volition (hard) seamlessly across the three hierarchical levels. Yet, as being static in nature, most of them aim to explain the structure of conscious experience per se without accounting for the successive alternation of conscious states over time. Many authors attempt to compare these theories (Doerig et al., 2021; Del Pin et al., 2021; Signorelli et al., 2021), or even to reconcile some of them (Shea and Frith, 2019; Chang et al., 2020; Graziano et al., 2020; Mashour et al., 2020; Northoff and Lamme, 2020; Mallatt, 2021; Seth and Hohwy, 2021; VanRullen and Kanai, 2021; Niikawa et al., 2022). Another tendency is to incorporate these static theories into a more general dynamical framework such as the Temporo-spatial Theory of Consciousness (Northoff and Zilio, 2022) which is somewhat reminiscent of Operational Architectonics (Fingelkurts et al., 2010), the whole-brain mechanistic models from a bottom-up perspective (Cofré et al., 2020), or the self-organizing harmonic modes coupled with the free energy principle (Safron, 2020).
In general, all these theories are not concerned with brain dynamics at the underlying hard level from which conscious states emerge. As being static in design, they have also missed another fundamental aspect of consciousness, namely, its cognitive evolution over a lifetime going by accumulating new knowledge and skills. Accordingly, the stream of consciousness (though implied) is not properly defined. At the same time, they all ascribe a special, active role to consciousness while being indifferent to the free will problem by adopting the view that consciousness can somehow influence brain dynamics at the psyche level. It is implicitly assumed that consciousness: (i) facilitates learning; (ii) is useful for voluntary decision-making; and (iii) provides stability to the objects of experience in an egocentric framework (Seth and Baars, 2005).
For example, Integrated Information Theory (IIT) defines consciousness as information a system is able to integrate. It does not consider perception, cognition, and action at all by aiming at the quantitative account of phenomenal consciousness due to “irreducible causal mechanisms” (Oizumi et al., 2014). It makes the striking conclusion that “a conscious choice is freer, the more it is causally constrained within a maximally irreducible complex” (Tononi, 2013) without explaining how free volition might be manifested there. Such a kind of volition turns out to be just superdeterministic (see below). It could well account for the individuality of subjective experience as specified by the complex at the psyche level but not for the indeterminism of volition at the hard level. The IIT approach can be even extended to a general idea that the very evolution of life is just the triumph of determinism. It argues that living systems can thrive across the universe as soon as these autonomous systems have more cause-effect power than non-living environments (Marshall et al., 2017).
On the contrary, Predictive Processing Theory (PPT) can well explain perception, cognition, and action by making the brain an error-minimizing machine (Clark, 2013; Seth and Hohwy, 2021). Although PPT is unclear about where exactly, between priors and posteriors, the conscious experience should appear, it can in principle separate discrete conscious states emerging at the psyche level as ultimate decisions of Bayesian learning from unconscious predictive processing at the soft level. Nevertheless, PPT cannot still account for free volition, which is covertly embedded in attentional effort and active inference (Friston et al., 2013; Pezzulo et al., 2018). This is just the point where free will and active consciousness converge. If consciousness is an algorithm for the maximization of resilience (Rudrauf et al., 2017), then PPT has to explain how and why conscious processing in the brain should differ from deep machine learning in AI systems, which can exploit the same computational models but lack both volition and conscious experience.
Another dominant theory, Global Workspace Theory (GWT), relies explicitly on the active role of consciousness at the psyche level, which is required for global access, broadcasting information, and self-monitoring (Dehaene and Changeux, 2011; Baars et al., 2013). According to the theory, a physical system “whose successive states unfold according to a deterministic rule can still be described as having free will, if it is able to represent a goal and to estimate the outcomes of its actions before initiating them” (Dehaene and Naccache, 2001). Thus, GWT adopts just the aforementioned scenario with Alice deciding where she will be tonight. It is therefore not surprising that GWT does not suggest any obstacle to machine consciousness (Dehaene et al., 2017), which by virtue of its cognitive architecture could be spontaneously endowed with free will.
Finally, psychological theories such as Higher-Order Thought (Lau and Rosenthal, 2011), Attention Schema (Graziano et al., 2020), Radical Plasticity Thesis (Cleeremans, 2011), or Self Comes to Mind (Damasio, 2010) argue that self-awareness or metacognition would separate conscious states from unconscious processing via self-referential mechanisms. These mechanisms would make the brain aware of its own states, unlike other biological and artificial networks. Accordingly, conscious will in these theories is similar to “conscious veto” suggested by Libet (1985) to circumvent the findings of his famous free will experiments.
The Libet-type experiments have been based on two temporal measures: the readiness potential detected from the supplementary motor area, and the awareness of wanting to move reported with the clock. The delay observed between neural motor predictors at the hard level and conscious intentions at the psyche level was around several hundred milliseconds (Libet, 1985; Guggisberg and Mottaz, 2013; Schultze-Kraft et al., 2016), thereby making conscious intentions a post-factum phenomenon. Libet had proposed that it could occur due to conscious deliberations before consciousness could block an action with the explicit veto on the movement. Nevertheless, since any kind of intentional veto must also be causally processed, it has been noted that the blocking itself could be predetermined (Soon et al., 2013).
Yet, some authors assume that noisy neural fluctuations can be involved in self-initiated actions (Schurger et al., 2012). They argue that the key precursor process for triggering internally-generated actions could be essentially random in a stochastic framework (Khalighinejad et al., 2018). First, those actions generated internally by the brain from noisy fluctuations would have little relevance to the ability of consciousness to make its own choice. Meanwhile, in the psychological theories mentioned above, conscious will would involve higher-order thoughts to check many moves ahead of time, and then with this information to make a free choice (Rolls, 2020). Second, stochastic noise as being classical (thermal) in nature does not violate determinism to account for freely generated actions even on the brain’s authorship. Something else is needed.
In contrast, quantum-inspired theories of consciousness take seriously the free will problem by adopting quantum indeterminism that might affect brain dynamics at the hard level. They are based on the conceptualization of free will suggested by Bell in his famous theorem (Bell, 1993). The key assumption of Bell was that Alice’s free choice (as defined above) should not be controlled by any hidden (unknown to our modern knowledge) deterministic variables λ. For example, these variables might include all the internal and external information about the past of Alice’s brain and everything in her environment. The choice is then formalized by conditional probability,
The theorem had shown that under measurements of preciously prepared quantum experiments no such variables should exist in principle unless we had to agree on a “cosmic conspiracy” that constrained Alice to make not any choice but just the one that did not violate standard statistical correlations of the Bell’s inequality (Gallicchio et al., 2014). Note that, as suggested beyond the neuroscientific context of Libet-type experiments, Equation (1) does not discriminate between a choice made by Alice with her conscious will, and a choice internally generated by her brain itself. The only thing required there is that A has not been predetermined from the past by λ. Thus, by ruling out conscious will from Libet-type experiments, only the indeterminism of neural activity can account for the Bell theorem, which has been well-confirmed experimentally (e.g., Aspect et al., 1982).
Orchestrated Objective Reduction (Orch OR) of Hameroff and Penrose (2014), the most known of quantum-inspired theories of consciousness, is explicitly based on the indeterminism of the wavefunction collapse or “objective reduction” (OR), also known as the measurement problem. In quantum mechanics, the measurement problem has the striking property of observer-dependence. In contrast, Penrose argues for a real quantum-gravitational OR that occurs everywhere in the universe, independently of observation, as spontaneous “proto-conscious” events that are then orchestrated in microtubules of the brain to give rise to consciousness and volition. According to Orch OR, consciousness must be in principle incomputable as being orchestrated (Orch) by quantum entanglement with consequent OR. Nonlocal correlations between microtubules (for global binding of dissociated brain regions) and backward time referral (for closed causal loops) are then required “to rescue causal agency and conscious free will” (Hameroff, 2012).
Remarkably, the mathematicians Conway and Kochen in their Free Will Theorem (a modified version of the Bell theorem) make a statement very similar to that of Penrose (though without concerning themselves with OR): “If conscious observers have a certain freedom to choose, then particles already have their own share of this freedom” (Conway and Kochen, 2008). It is often argued that the quantum randomness on which the statement reposes has little to do with “free will” thought to be caused by a reason rather than by chance (Koch, 2009; Aaronson, 2016). Conway and Kochen had however noted that if a subject’s action was indeed free of the past, then it should be difficult to find a testable (i.e., objective from a third-person perspective) difference between physical randomness and the subject’s genuine behavioral freedom, both not being predetermined by the previous history of the universe. In the context of neuroscience, their statement has to be inverted and specified as follows:
If particles have some freedom to respond to the environment independently of the past, then the observers have the same kind of initial freedom due to a quantum mechanism Bell-certified by Equation (1) in their brains.
Of course, ascribing free will to living or non-living systems does not depend anyhow on quantum effects. Volition is typically associated with the action a system is capable of initiating. It necessarily involves causation. However, what is the sense of saying that a clockwork toy has the volition to move, or, that a more sophisticated AI system is self-initiated in performing a cognitive task? To discriminate between volition and trivial energy flow in an arbitrary (natural or artificial) system the former must not be predetermined from the past. Volition is not a matter of consciousness or cognition but a matter of causal freedom. Under this definition, quantum particles can have some sort of freedom but this freedom is washed out in macroscopic systems.
To bind volition with consciousness, we need to translate physical predetermination into the notion of predictability. First of all, deterministic systems can in principle be perfectly predictable. It is already a matter of our current knowledge and technology. Now let us modify the famous Turing test where a machine might mimic a human by answering various questions. Suppose, a future machine can be trained to predict precisely Alice’s every choice ahead of time. The machine can thus perfectly simulate Alice’s stream of consciousness so that we might be fooled by asking any question. Why then should we deny that the machine possesses a copy of Alice’s consciousness and, thus, is itself conscious?
Hence, if we disagree that humans are very sophisticated but still deterministic and copyable machines, there must be something that prevents one from making a deterministic machine conscious. This is volition. Indeed, if Alice’s brain is able to make a choice not predetermined from the past, there cannot be in principle a perfect predictor of Alice’s stream of consciousness. This makes her consciousness unique, i.e., non-copyable for any future technology. Thus, the reason to assume the above statement stems fundamentally from the evolutionary perspective.
CET suggests a dynamical model based on a framework, drawn over diverse neuroscientific domains, with contributions from critical dynamics, predictive processing, and evolutionary neurobiology to approach a unified theory of consciousness grounded in physics. The widespread idea that brains are error-minimizing machines has neglected a crucial ingredient of its adaptive framework: before minimizing an error the brain should already have that error internally triggered under cognitive correction. CET argues that the brain can be viewed more generally as both an error-amplifier and a modulator of the primitive volitional reflexes based on chemo- and photoreceptors of unicellular organisms. In doing so, CET makes the assumption that brains have primarily evolved as volitional (quantum in origin) subsystems of organisms at the hard level. CET emphasizes the importance of randomness in evolution (Yurchenko, 2021a) contrary to the idea that life can be viewed as the triumph of determinism (Marshall et al., 2017).
Volition from The Evolutionary PerspectiveIn modern science of consciousness, the Cartesian presumption that animals are only biological automata incapable of experiencing conscious and emotional states looks chauvinistic or even perverse (Lamme, 2018; Fields et al., 2021). Nonetheless, the tradition to separate humans from the rest of the animal kingdom is still persistent. Now the division has shifted to the free will debate. It is assumed that only humans enjoy free will due to the sophisticated computations the human brain is able to carry out at the soft level, while other animals, though passively conscious, are deprived of this gift (Doyle, 2009). CET finds this division evolutionarily implausible.
Maintaining homeostasis in the face of dangerous and unfortunate environmental conditions is the basic imperative for the evolution of the brain (Ashby, 1960). The only valuable advantage the living systems might gain over non-living systems is the ability to decide their way freely among stimulus-reaction repertoires. This ability is commonly referred to as volition. Meanwhile, volitional mechanisms of the brain are still ignored in cognitive neuroscience, remaining hidden under attentional effort, enactive cognitive function, and conscious control. CET argues that volition is a key neural mechanism of evolution placed between organisms and non-living systems. Life could not have flourished on Earth without volitional mechanisms which are the only neural candidate that opens a door to the stimulus-reaction repertoires over the “tyranny” of causal chains.
The underlying idea here is the one of evolutionary neurobiology: the ultimate aim of brain evolution over species, from simplest organisms to humans, was to maximize their adaptive success. Under selection pressure, the main function of neural systems resulted in their ability to integrate and process a very large number of sensory inputs and motor responses occurring in parallel (Edelman, 2003). Thus, the cognitive evolution of the brain over every organism’s lifetime (ontogeny) and the general evolution of the brain over species (phylogeny) should go hand in hand to promote each other. CET makes the general assumption that after acquiring free volition mechanisms to overcome the fatalism of cause-effect interactions which govern completely non-living systems, organisms could have advanced their adaptive success only by evolving cognitive functions capable of predicting future events. Cognition and memory should have evolved exclusively as adaptive (computational) abilities of organisms at the soft level to benefit from some underlying volitional mechanism at the hard level. Otherwise, evolution would have had no reason to advance those adaptive properties over rigid causal chains in deterministic systems.
How much is it plausible that evolution had endowed primitive networks of simplest organisms with some kind of freedom which would have been of value for them in survival and reproduction? Can invertebrates have some volitional mechanism, evolutionarily embedded in their neural networks to make a choice not predetermined by the past? CET argues that primitive neural networks should have primarily evolved as free-volitional subsystems of organisms, not as deterministic prediction machines (Knill and Pouget, 2004; Clark, 2013), requiring larger biological resources. Accordingly, their conscious properties, typically related to higher animals, should have appeared later than their unconscious cognitive functions presented in invertebrates (Brembs, 2011). Instead of being an active participant that creates an internal representation of the environment, the conscious experience would be an extension of primitive volitional-emotional motivations (Mashour and Alkire, 2013). Thus, contrary to the idea that evolution had evolved consciousness to perform complex volitional actions (Pennartz et al., 2019; Feinberg and Mallatt, 2020), in CET consciousness is a byproduct of both volitional mechanisms and unconscious cognitive processing grounded in sensorimotor coupling (Engel et al., 2013). With this assumption on the origins of consciousness, CET suggests a radically new and physically rigorous solution to the free will problem.
Unlike the known quantum-inspired models involving the most mysterious quantum-mechanical effects to account for quantum computing and/or quantum memory storage that might directly mediate consciousness (Hameroff and Penrose, 2014; Fisher, 2015; Georgiev, 2020), CET suggests a minimal use of quantum randomness at the sub-cellular level. This refers to the most reliable mechanisms like the Beck-Eccles quantum trigger, proposed as a quantum-mechanical model for the mechanism of exocytosis (Beck and Eccles, 1992, 1998). Exocytosis is a discrete all-or-nothing event consisting of the opening of presynaptic membrane channels with the release of neurotransmitters in the synaptic cleft. The trigger is based on the tunneling of a quasi-particle across a potential energy barrier. Beck and Eccles (1992) argue that “the mental intention (the volition) becomes neurally effective by momentarily increasing the probability of exocytosis in selected cortical areas such as the supplementary motor area.” Thus, they maintain the cortex-centered conceptualization of conscious free will in Libet-type experiments.
Instead, CET places the mechanism in the brainstem to account for the indeterminism of unconscious free volition initiated in the arousal nuclei. This key quantized event could then be physically amplified across many spatiotemporal scales due to neuronal avalanches which are intrinsic to SOC (Blanchard et al., 2000; Beggs and Plenz, 2003; Chialvo, 2010; Tognoli and Kelso, 2014). The brainstem is a phylogenetically ancient brain region that comprises various nuclei executing many vital autonomic functions for maintaining homeostasis such as blood pressure, heartbeat, or blood sugar levels (Paus, 2000; Parvizi and Damasio, 2001). Those autonomic functions should have evolved much early than thalamocortical regions engaged in elaborating cognitive contents and conscious experience.
Remarkably, this ancient region also contains the arousal centers responsible for permanent vigilance upon which the stream of consciousness reposes. Its arousal machinery is a precondition for behavior and conscious experience (Mashour and Alkire, 2013). CET proposes that each conscious state in the stream is volitionally driven at the causal (hard) level from subcortical arousal centers via the ascending reticular activating system (ARAS) to thalamocortical systems, involved in perception and cognition at the computational (soft) level. In CET, conscious experience emerges at the psyche level as a passive phenomenon of cognitive brain evolution that goes by accumulating new knowledge and skills freshly updated in memory networks.
By postulating the quantum mechanism that will be called the “neurophysiological free-volition mechanism” (NFVM) and placed into the brainstem, CET can account for the indeterminism of brain dynamics without resorting to large-scale quantum mysteries. The corollaries are the following. While brain dynamics will still be presented by classical stochastic processes as those traditionally described in standard models of neuroscience, the stream of consciousness will no longer be predetermined from the past by hidden deterministic variables λ due to the presence of the NFVM. This makes Alice’s consciousness (whose secured privacy is guaranteed by the Bell-certified NFVM) unique but gives her consciousness no power over the brain unlike typical scenarios mentioned above.
In general, the controversy around free will has been inherited by the science of consciousness from the mind-body problem, originally discussed in philosophy. What exactly should be associated with Alice? Is it her brain or her consciousness generated by her brain (leaving aside her body that makes the brain alive and functional)? In CET, if Alice is associated with her brain, she has free volition. On the contrary, if Alice is associated with her consciousness, she has no free will. Consider, for instance, the following sentence: “I can have true free will: I can have true alternatives, true freedom to choose among them, true will to cause what I have decided, and eventually true responsibility” (Tononi et al., 2022). For CET, the validity of this statement depends on how the “I” is conceptualized.
This also implies that not integrated information of irreducible causal mechanisms (Tononi, 2008; Oizumi et al., 2014), architecture peculiarities of neural networks (Dehaene and Naccache, 2001; Baars et al., 2013), cognitive processing (Clark, 2013), or higher-order linguistic thoughts (Lau and Rosenthal, 2011; Rolls, 2012) but free volition, inherited by the brain from fast and random reflexes rooted in chemo-, magneto-, and photoreceptor cells which are very sensitive to quantum effects (Arndt et al., 2009; Brookes, 2017; McFadden and Al-Khalili, 2018) is the main obstacle that prevents computer scientists from making deterministic machines conscious.
The Stream of Consciousness in Brain DynamicsConsciousness has been a prescientific concept with a number of different connotations that relate to various aspects of conscious experience. The science of consciousness must rely on the fact that consciousness is a dynamic process, not a thing or a capacity (James, 1950). It should also take into account that the observation of single cells activity has little to say about mental processes. Large-scale interactions between neural networks are therefore more important than the contribution of individual neurons per se. Thus, only neural dynamics at the macro-scale can account for global brain states accompanied by conscious experience. Most attempts to understand the neural mechanisms of consciousness have proceeded by searching for the “neural correlates of consciousness” or NCC (Crick and Koch, 2003). However, correlations by themselves do not provide explanations and there is a need for a framework connecting fundamental aspects of conscious experience at the phenomenal (psyche) level to the corresponding aspects of brain dynamics at the underlying causal (hard) level.
The evolution of consciousness depends on the physical continuous dynamics of the brain, comprising about 1011 neurons connected to each other by thousands of synapses. Clearly, consciousness not only requires neural correlates with their appropriate anatomical architecture but also the time to be processed. A wealth of experimental evidence suggests that conscious experience is a discrete phenomenon processed unconsciously at the soft level (see Section “Temporal resolution of the stream of consciousness”). Moreover, a pure biophysical approach to studying neural activity at micro- and mesoscopic scales cannot account for subjective, internally generated mental phenomena without resorting to their contextual emergence at the macroscopic scale (Atmanspacher and beim Graben, 2007). It has been pointed out many times that there should be an optimal spatio-temporal grain size at which the brain can generate phenomenal experience (Tononi, 2008; Chang et al., 2020). In general, the correspondence between conscious states and neural processes should be provided by mapping brain dynamics onto the state space of consciousness.
CET compresses all these requirements into three key prerequisites:
1. Physicalism (mind-brain identity): consciousness depends entirely on brain activity governed by natural laws at the hard level, not on anything else;
2. Dynamism (temporality): consciousness not only requires the NCC but also the time to be cognitively processed at the soft level;
3. Contextuality (scale-dependence): only large-scale brain dynamics can account for the emergence of conscious states at the psyche level.
A principled distinction between CET and classical theories is that CET involves quantum indeterminism. On the other hand, while quantum-inspired theories engage quantum computing across the whole cortex to account for consciousness and free volition, CET makes minimal use of quantum randomness placed into the arousal nuclei of the brainstem to initiate cognitive processing due to stochastic brain dynamics that are classical in nature. Thus, CET is a semi-classical physical theory of consciousness.
Deriving consciousness from brain dynamicsBased on the three prerequisites, CET will model consciousness as the stream of macrostates, each specified by a particular structural-functional configuration of the whole-brain network ?, where NCC ⊆ ?. Here ? stands for a graph G = (N, E), where N = |?| is the set of nodes (ideally, neurons), and E ⊆ N × N is the set of edges (ideally, synapses). Because it is computationally impossible to operate on N ≈ 1011, the first step in formalizing the stream of consciousness is to approximate large-scale brain dynamics at the hard level. To derive the stream from the SOC approach, CET refers to the Langevin formalism as a most general description of a system that depends upon a deterministic flow y and stochastic fluctuations ω [which, note, do not discriminate between quantum (i.e., ontic) and statistical (i.e., epistemic) randomness, e.g., in Brownian motion]
dψ=−γψ(t)dt+dω(t)(2)The formalism can then be transformed into different models depending on the way researchers will adopt that formalism in their study. Those models may be biophysical (mean-field) approximations (Breakspear, 2017; Parr et al., 2020) or phenomenological (synchronization) models such as Stuart-Landau, Kuramoto, Haken-Kelso-Bunz, and other models (Cofré et al., 2020; Kelso, 2021; Kraikivski, 2022; Vohryzek et al., 2022).
Here ψ (?, t) is a descriptive function whose representation by the order parameter in a phase space O should account for metastability, avalanches, and SOC of the global neural activity wandering among dynamical attractors (Kelso, 1995). Originally grounded in physics, chemistry, and biology (Bak, 1996; Jensen, 1998; Haken, 2004), SOC is thought to be of crucial importance in neural activity as it poises the brain on the edge between order and disorder near a bifurcation (Blanchard et al., 2000; Chialvo, 2010; Beggs and Timme, 2012; Deco et al., 2015). This allows the neural network ? to exhibit the properties of scale-free networks and to produce flexible patterns of coordination dynamics at the hard level. This, in turn, can generate a repertoire of different conscious states at the psyche level, thereby increasing an adaptive behavioral response of the organism to given environmental stimuli (Hesse and Gross, 2014; Cocchi et al., 2017; Dahmen et al., 2019).
CET postulates the emergence of consciousness from brain dynamics at critical points as its derivative extracted in discretized time τ,
The continuous/discrete dichotomy appears naturally between the brain dynamics, described at the causal (hard) level, and the stream S(τ) of conscious states, presented at the phenomenal (psyche) level. In effect, Equation (3) should capture the instantaneous transitions from continuous brain dynamics to discretized conscious states, identified with a single point o ∈ O in a phase space. The phase transitions in brain dynamics occurring at discrete moments of time can then be viewed as the manifestation of pulsating consciousness in the framework of cinematic theory of cognition (Freeman, 2006). This approach finds now experimental evidence in many studies (Haimovici et al., 2013; Mediano et al., 2016; Tagliazucchi, 2017; Demertzi et al., 2019; Kim and Lee, 2019) showing that only states integrated near criticality can ignite conscious experience in the brain.
CET suggests a simple mathematical analogy between consciousness and the physical force, derived from the momentum in Newtonian mechanics,
Seeing consciousness as a “mental force” seems to be more moderate and accurate, at least ontologically, than viewing consciousness as a fundamental property of matter like mass, charge, and energy (Tononi, 2008). The analogy between conscious experience and mass, advocated by IIT, is based on a quantitative account of the level of consciousness measured by integrated information Φ a single complex of irreducible causal mechanisms might generate (Oizumi et al., 2014). IIT argues: if a complex can generate Φ, no matter whether it is organic or not, it will have consciousness (Tononi and Koch, 2015). Instead, CET brings into focus the dynamism of consciousness. The analogy with force goes in line with the fact there is a tiny but principled distinction between mass and force in physics by Equation (4): the former is a scalar quantity which is indeed constantly intrinsic to a system, whereas the latter is a dynamical characteristic of motion defined by a vector quantity of a system’s action that can vanish in inertial states.
Similarly, Equation (3) represents consciousness as a dynamical characteristic of the neural network ? not as an intrinsic potency of causal mechanisms in the brain or anywhere else. According to this conceptualization, the brain has no mental force if its dynamics depart from criticality, as it occurs in unconscious states such as coma, sleep, or general anesthesia (Hudetz et al., 2014; Tagliazucchi et al., 2016; Golkowski et al., 2019; Huang et al., 2020), but not in resting states where neural activity preserves criticality (Deco and Jirsa, 2012; Barttfeld et al., 2015). On the other hand, even in critical dynamics, the brain lacks the mental force during some interval Δt until a next conscious state is unconsciously processed.
To make the above analogy more comprehensive, imagine a clockwork toy, say, a jumping frog. The engine of the toy will impel it to iterate the same jump over and over. The force occurs only at discrete moments of jump-initiation between which motion decays. In critical dynamics, the brain exhibits flexible patterns of coordinated dynamics which decay after some critical value (Gollo et al., 2012; Tognoli and Kelso, 2014). Conscious experience can be ignited only on the edge between the two phases as if the brain accumulated information for triggering the next “jump” with some mental force at discrete moments of time (Figure 1A).
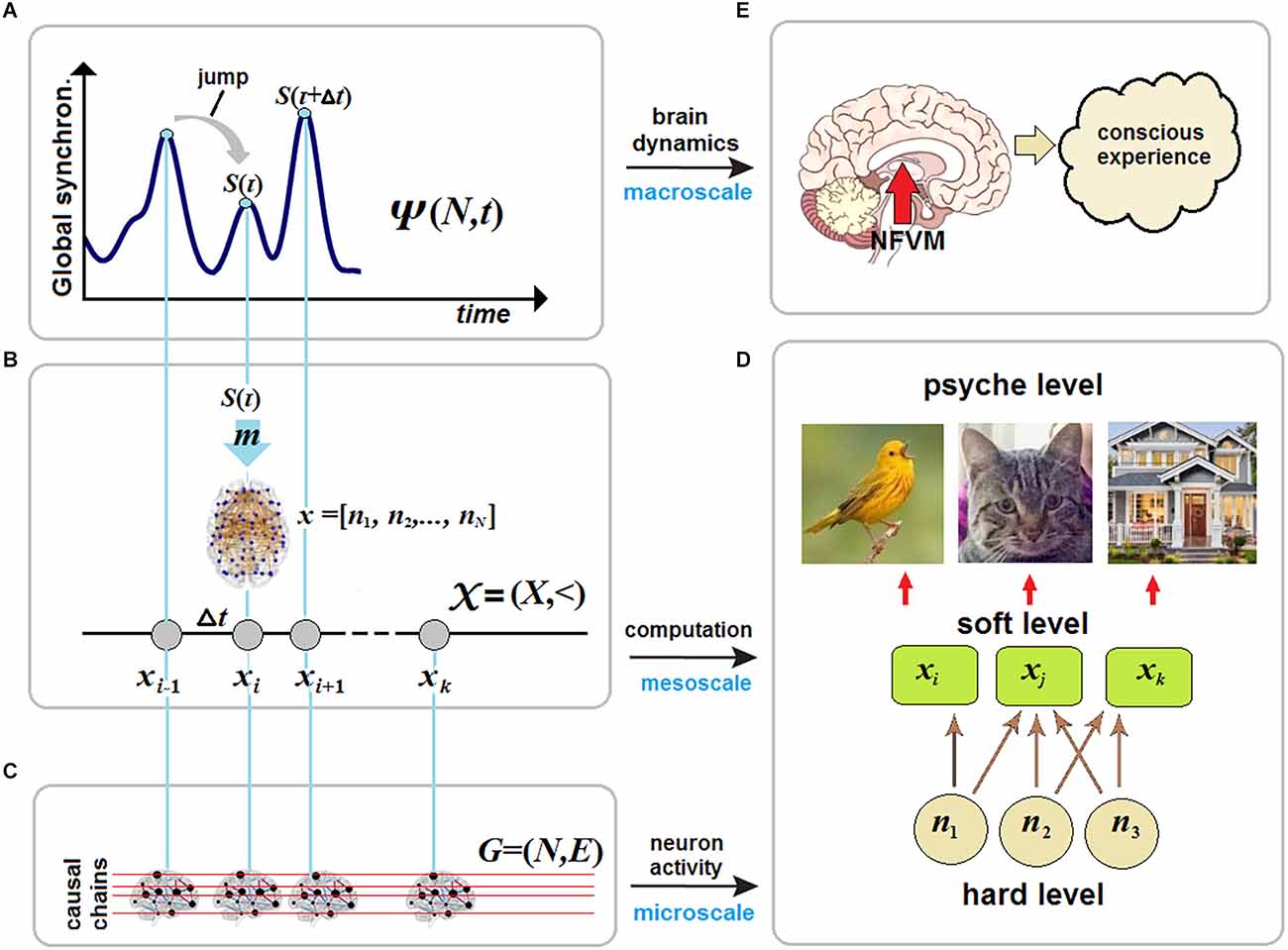
Figure 1. Brain criticality and conscious experience. (A) In large-scale brain dynamics, conscious states emerge at critical points near phase transitions between synchronization (order) and desynchronization (disorder) patterns of neural activity at the microscale. (B) The map m transforms each critical point of brain dynamics, described in a phase space, onto the whole-brain network ? in a vector space as a particular NCC responsible for a certain conscious state at that moment of time. (C) The stream of consciousness can then be formalized as a discrete chain of states (N-dimensional vectors) and studied in causal dynamical modeling as a directed acyclic graph. (D) In neural pleiotropy, many neurons constitute a particular NCC for producing a certain conscious percept, thereby involving a single neuron in generating very different percepts. (E) Conscious experience is a product of unconscious computations initiated by the brainstem at the hard level and accomplished by various thalamocortical systems at the soft level.
Temporal resolution of the stream of consciousnessAccording to Equation (3), the brain needs some time to process a new conscious state from the previous one. There are two complementary ways to estimate the interval Δt– either by monitoring brain dynamics to calculate phase transitions at the hard level or by obtaining a subjective report at the psyche level. Unfortunately, both approaches do not provide an exact estimate. The monitoring of brain dynamics is non-trivial because of the heterogeneous intrinsic timescales involved. An averaged interval is usually reported to be about 200 ms (Kozma and Freeman, 2017; Deco et al., 2019). The second approach, based on first-person reportability, is affected by the problem of multiple temporal resolutions (White, 2018).
To comprise both approaches, we assign the interval to a wide window Δt ≈ 100–450 ms that encompasses multiple experimental findings—from the periodicity of attentional cycles at approximately 7–13 Hz (VanRullen et al., 2014) to the attentional blink on masked targets separated by 200–450 ms (Shapiro et al., 1997; Drissi-Daoudi et al., 2019). Yet, an important neurophysiological aspect of brain dynamics is that the stream S(τ) cannot be normally delayed for a period longer than about 300 ms according to the timescale proposed to be crucial for the emergence of consciousness (Dehaene and Changeux, 2011). Consciousness spontaneously fades after that period, for example, in anesthetized states (Hudetz et al., 2014; Tagliazucchi et al., 2016).
Only states that emerge globally and are integrated at critical points are conscious. In the stream, each state appears instantaneously as a snapshot accompanied by a phenomenal percept of the “specious present” (Varela, 1999), which provides human (and most likely animal) observers of the external world with an egocentric frame of reference, preserving its stability over time (Seth and Baars, 2005). Due to the cognitive updating of this self-referential frame by the acquisition of new contents, consciousness remains well informed at the psyche level about what is occurring around. Thus, while conscious states emerge only at discrete critical points of brain dynamics, subjects can still feel the continuity of being and acquire the illusion of a persistent presence.
There are also arguments for a continuous flow of conscious experience (Fekete et al., 2018; Kent and Wittmann, 2021). However, these are typically grounded in the phenomenology of consciousness and time instead of being based on the neuroscience of consciousness and the physics of time. Yet, they do it without making a difference between the rigorous notion of mathematical continuum and something that can be continued. If such arguments have any merit, their core can be formulated like this: “If conscious experience is produced by the brain, then it would seem that there must be a lawful relation between the state of the brain at the given time and the percept that the person is experiencing… because experiencing or perceiving is an activity, not something to be looked at” (Brette, 2019). Indeed, such a “lawful relation” must be suggested.
Equation (3) explains how discrete conscious snapshots at the psyche level can be separated from continuous brain dynamics at the hard level. Moreover, because consciousness does not observe how the experience was processed by the brain, awareness requires no time to be ignited. Since the ignition across the brain’s workspace occurs phenomenally due to SOC, there is no dynamical process that should transmit information into a special site of the brain to reach the subject’s awareness. Experience is the information the brain has unconsciously computed at that specious moment τ. The illusion of temporal continuity of consciousness is merely a trivial corollary of its self-evidential nature: consciousness cannot in principle detect its own absence in the brain. Whenever consciousness searches for itself, the search happens successfully for an obvious reason: in the stream, consciousness will always be present in introspection. Likewise, whenever we look in a mirror, we always see ourselves as if we were constantly there. We know that it is not true, but it is impossible to catch the mirror before our image will appear.
In particular, the discreteness of conscious experience explains how the subjective distortion of perceived temporal duration produced by emotionally charged events and causing the sense of dilated time in the stream S(τ) can occur. Overestimation of time is associated with many factors such as enhanced memory encoding or intensive involvement of the limbic system and the interconnection with medial cortical areas (Dirnberger et al., 2012). This depends on how the states were unconsciously processed over multiple timescales in brain dynamics (Merchant et al., 2013) while their sensory-cognitive contents had been compressed over temporal chunks T = ∑ Δt, composed of non-overlapping intervals. Otherwise, for the sake of continuous conscious perception, we should agree on something like space-time effects in Relativity theory as if a perceived chunk T, as compared to a physically real TR, would indeed be dilated by the Lorenz transformations T=TR/1−υ2/c2. Thus, the discrete model of the stream S(τ) proposes a more reasonable and parsimonious explanation of the subjective distortion in time perception as merely depending on a ratio TΔt, i.e., on a variation of Δt during which those emotionally charged conscious states were processed.
The passage of time (whatever it is physically) cannot be perceived at all without sensory-cognitive contents processed by different brain systems at the soft level and integrated globally at the psyche level. This also explains why time perception vanishes completely in altered states of consciousness such as sleep, anesthesia, or epileptic seizures. It occurs as brain dynamics deviate from criticality upon which discrete conscious experience entirely reposes (Tagliazucchi et al., 2016).
The stream of consciousness as a causal chain of discrete statesNow let m: O → V be a map from the phase space O onto a vector space V over the product N × N of all neurons of the brain network ?. The map returns S(τ) from a point o ∈ O to an N-dimensional vector x = [n1, n2,‥., nN], where ni = 1 or ni = 0 stand for neurons active or inactive at the given time. We write (omitting details),
Thus, each state S(τ) is represented now by x as a certain structural-functional configuration of ? that is responsible for that subjective snapshot at the moment τ. The discreteness of the stream signifies that all the conscious states could, at least in principle, be naturally enumerated from a subject’s birth, not merely by a lag in experimental settings.
Let the brain bring consciousness to a state xi at the moment τ = t. We can rewrite Equation (5) as follows.
The next conscious state will emerge over the time interval as
ψ (?,t+Δt)=xi+1(7)In the timeless description, the stream S(τ) is a discrete causal chain ? = (X,<) (Figure 1B), where xi ∈ X and whose relation < standing for the causal (and temporal) order is transitive and irreflexive. Here the irreflexivity means ∀ixi<xi that forbids closed causal loops and, in particular, instantaneous feedback circuitry in brain dynamics that might somehow enable consciousness with causal power over the brain, for example, due to the presumed quantum temporal back-referral of information in Orch OR (Hameroff and Penrose, 2014). CET strictly rejects the possibility that consciousness could—classically or quantum-mechanically—choose a state to arrive at.
Consciousness is a physically classical macro-phenomenon (though quantum-triggered) that is always local in space and time. For consciousness, planning something, as discussed in Alice’s scenario, does not mean to be already in that state. In other words, every state in S(τ) is actual, and any chosen state should also be actual. However, it is physically impossible for the brain or for the stream of consciousness to be ahead of itself. Physically, the brain is exactly where it is. Mentally, consciousness is what it is just now. Even assuming quantum temporal back-referral, closed causal loops would be involved, as if consciousness would be now in a state it had already chosen in the state before. On any account, this would imply that before choosing a state consciousness should already be in that state despite irreflexivity.
Herzog et al. (2016) suggest a discrete model of consciousness very similar to that of CET. The authors assume that conscious states represent integrated, meaningful outputs of unconscious (modular) computations carried out at the soft level which had been causally provided by dynamical attractors at the hard level of interacting neurons. They compare a conscious snapshot with a k-dimensional feature vector Vi that contains a value for each relevant feature i of an external stimulus (e.g., color, size, sh
留言 (0)