
記住我
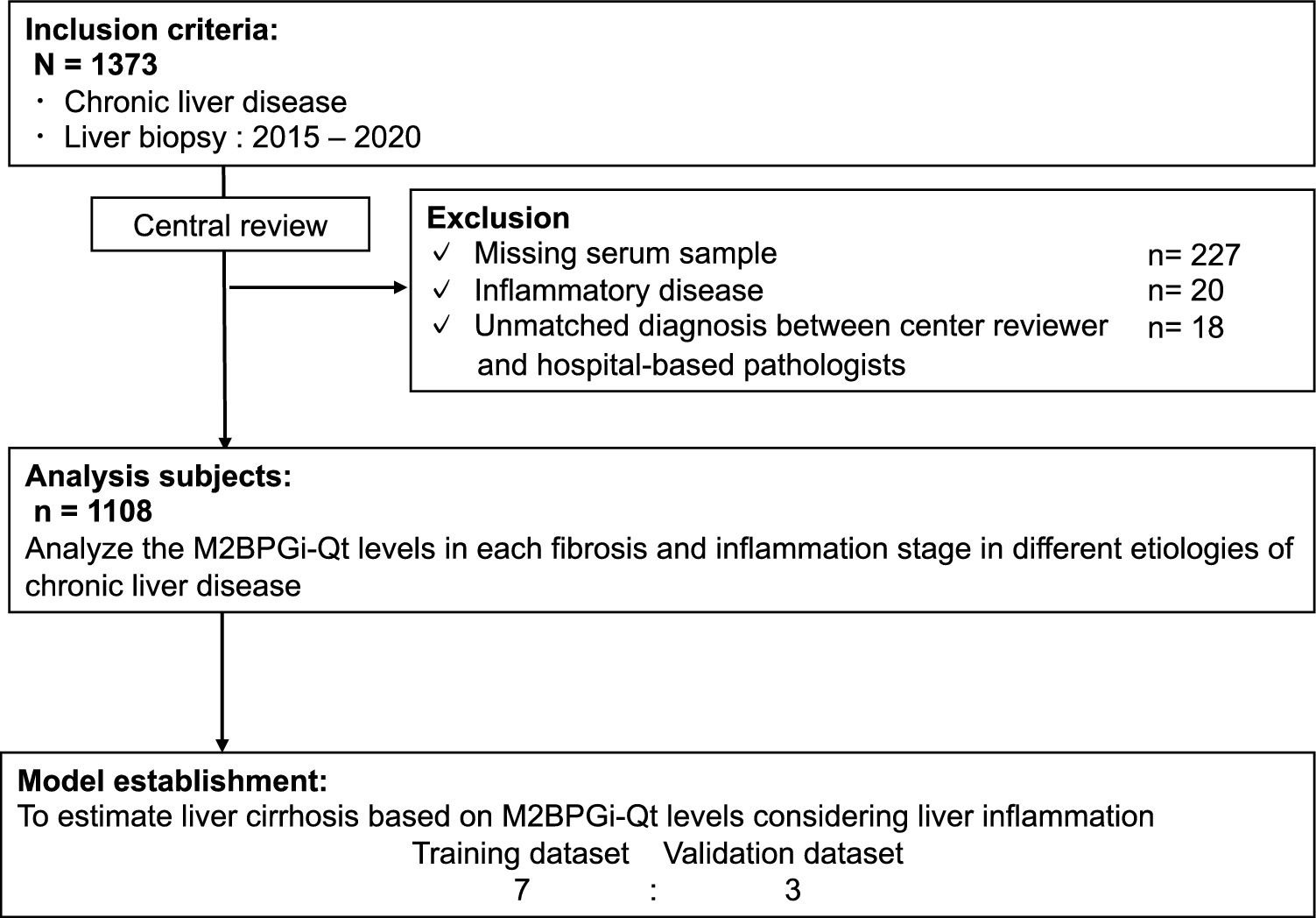


To investigate if loss of BMP signaling in resident stromal cells could contribute to polyp development, three different cell-specific conditional Cre-LoxP knockout mouse strains were generated. The VeCad-Cre driven model (BMPR1Afl/fl−VeCad) was used to investigate the loss of BMPR1A signaling specifically in the endothelial cells. A Col1a2 driven Cre (BMPR1Afl/fl−Col1a2) was used to investigate the effects of loss of BMP signaling in fibroblasts, while an SM22 driven Cre (BMPR1Afl/fl−SM22) was used to investigate the effect of BMPR1A loss in smooth muscle cells and myofibroblasts.
Mice with endothelial-specific BMPR1A loss were sacrificed 90, 180 or 356 days post-induction, after which the intestines were examined. Loss of BMPR1A signaling in endothelial cells did not result in any histological abnormalities. Immunohistochemical staining for the EYFP reporter protein showed the presence of EYFP-positive cells constituting the blood vessels (Supplementary Fig.1), confirming successful Cre-mediated recombination. Since no histological changes in the intestine were observed, this suggests that endothelial BMP signaling does not affect intestinal homeostasis significantly.
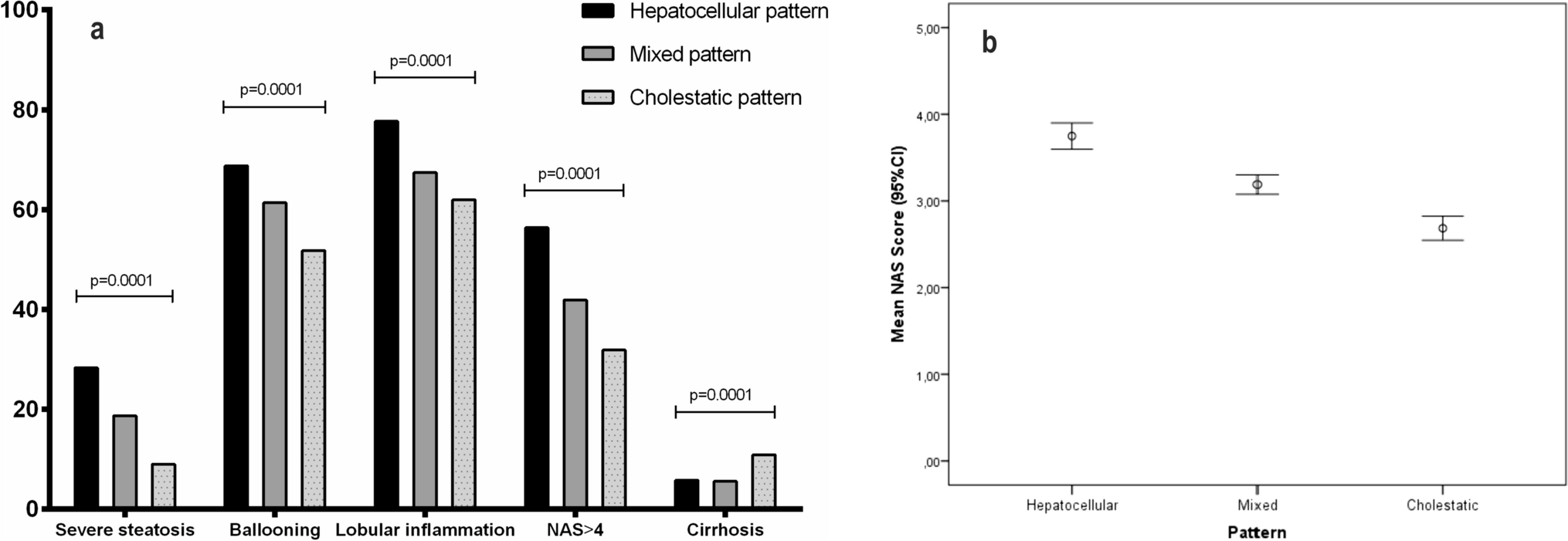
In contrast, major histological changes were observed in the intestines of BMPR1AΔ−Col1a2 mice 90 days post-induction (Fig. 1A–E). Immunohistochemical staining for the EYFP reporter protein showed the presence of EYFP-positive cells throughout the lamina propria, myofibroblasts and fibroblasts, of both the small intestine and colon verified successful Cre-induction (Supplementary Fig. 2A–E). Although the histological typical crypt-villus architecture was well preserved in the small intestine, numerous polyps had developed throughout the intestines, with an average of 29 polyps per mouse (Fig. 1F, P ≤ 0.0001). Even stronger histological changes were observed in the colon as almost the entire colonic epithelium showed signs of hyperproliferation (Fig. 1D and E). The crypt length was found to be 3.6-fold (P ≤ 0.0001) longer compared to the colonic mucosa of BMPR1Actrl−Col1a2 mice (Supplementary Fig. 3A). Large cystic structures filled with mucous were found scattered throughout the mucosa, surrounded by elongated crypts. The diffuse pattern of epithelial hyperproliferation made it difficult to distinguish individual lesions and hampered accurate polyp quantification. The Col1a2 promoter activity could explain the considerably stronger phenotype observed in the colon as Col1a2 expression was fourfold higher in the colon than in the small intestine (Supplementary Fig. 3B). In addition, colon tissue appears to have more EYFP-positive cells (Supplementary Fig. 3C–G), indicating increased recombination. Next, the BMPR1Afl/fl−SM22 mouse strain was used to investigate if a myofibroblast-specific BMPR1A knockout could also drive the phenotype observed in BMPR1AΔ−Col1a2 mice. The EYFP-staining showed the presence of EYFP-positive myofibroblasts in the submucosa and EYFP-positive smooth muscle cells in the muscle tissue verifying successful Cre-induction (Supplementary Fig. 2F–J). Although polyp formation was observed in the small intestines, a less severe phenotype was seen as only 3 out of 10 mice were found to have developed intestinal polyps (Fig. 1G–L). While BMPR1AΔ−Col1a2 affected the colon, myofibroblast-specific knockout did not lead to histological changes in the colon.
Fig. 1
Loss of the BMPR1A in fibroblasts and myofibroblasts leads to serrated polyp formation. A The loss of BMPR1A signaling in Col1a2 expressing fibroblasts results in substantial histological changes. A–C The villi appeared to be longer and wider and numerous polyps could be observed throughout the small intestine. D–E In the colon, substantial hyperproliferation could be observed and the crypt length was found to be increased 3.6-fold. Numerous cysts were observed scattered throughout the mucosa. F BMPR1A∆−Col1a2 mice formed on average 29 polyps/mouse (SD = 6). G–I On average 1 polyp developed per mouse in the BMPR1A∆−SM22 mice (SD = 2). D) Bars represent mean ± SD. P < 0.0001 (****)
Next, we analyzed the polyp histological subtype in the BMPR1AΔ−Col1a2 and BMPR1AΔ−SM22 mice. Contrary to what was expected, the polyps found in the BMPR1A KO mice did not resemble hamartomatous polyps, as observed in JPS patients. Instead, the polyps showed sawtooth-like serrations of the epithelium with abundant goblet cells (Supplementary Fig. 4 and 5). These features, typical for serrated polyps, are found in the polyps of both the fibroblast-specific BMPR1A knockout mice (BMPR1AΔ−Col1a2) and the myofibroblast-specific BMPR1A knockout mice (BMPR1AΔ−SM22). The presence of serrated polyps was confirmed by three independent pathologists.
Taken together, these data suggest that abrogation of fibroblast or myofibroblast-specific BMPR1A signaling, in contrast to endothelial BMPR1A signaling, leads to disturbed intestinal homeostasis and serrated polyp formation.
Intestinal BMPR1A-expressing fibroblasts regulate epithelial cell proliferationTo further investigate the severe histological changes observed in the intestines after disrupting fibroblast-specific BMPR1A signaling, we performed immunohistochemical stainings to investigate changes in the cellular composition. Staining for the stromal marker vimentin showed that the stroma had significantly expanded in the small intestines, polyps and colon of BMPR1AΔ−Col1a2 mice compared to control mice (Fig. 2A–C and J–L). Interestingly, the stromal abundance as quantified with the vimentin staining appears to correlate with the number of cells positive for the stem cell marker Olfm4+ and ki67+ proliferating cells (Fig. 2D–I and P). Olfm4 is not expressed in the mouse colon, but a similar correlation could be observed in the colon between vimentin and Ki67 (Fig. 2M–O and Q). An IHC staining for cleaved caspase 3 was performed to investigate if the increase in the number of epithelial cells could be due to a decrease in the number of cells undergoing apoptosis. An increase in the number of apoptotic cells was observed in the intestines from BMPR1AΔ−Col1a2 compared to controls, which was significant in the colon (Supplementary Fig. 4 J–K). To investigate whether the increase in proliferation is caused by the BMPR1AΔ fibroblasts, primary fibroblasts were isolated from induced BMPR1AΔ−Col1a2 mice and co-cultured with normal intestinal mouse organoids. BMPR1AΔ fibroblasts significantly enhanced organoid growth compared to fibroblasts isolated from Cre-negative or non-induced or non-induced BMPR1Afl/fl−Col1a2 mice (Fig. 2R–S), indicating a possible paracrine interaction. These data show that intestinal BMPR1AΔ fibroblasts can stimulate epithelial proliferation, potentially explaining the increased proliferation observed in vivo in BMPR1AΔ−Col1a2 mice.
Fig. 2
BMPR1AΔ-fibroblasts support epithelial cell proliferation. In the small intestines, immunohistochemical stainings on BMPR1A∆−Col1a2 tissue showed a significant increase of vimentin-positive stromal cells (A–C), which appeared to correlate with the increase in Olfm4+ stem cells (D–F) and Ki67+ proliferating cells (G–I). A significant increase of vimentin and Ki67 was also observed in the colon of the mice (J–O). P–Q The Pearson R correlation analyses showed a significant correlation between Olfm4 or Ki67 with vimentin in the small intestine and Ki67 and vimentin in the colon. R, S Co-culture experiments of normal intestinal organoids with either BMPR1Actrl−Col1a2 fibroblasts or BMPR1AΔ−Col1a2 fibroblasts showed that the size of the organoids increased significantly when they were co-cultured with BMPR1AΔ-fibroblasts. Bars represent mean ± SD except for P in which bars represent mean ± SEM. P < 0.05 (*), < 0.01 (**), < 0.001 (***) and < 0.0001 (****)
Loss of BMPR1A in fibroblasts results in increased pERK levels and increased TGF-β activityCRC is thought to develop through two main independent pathways. 20–30% of CRC is thought to develop through the serrated pathway, which is associated with increased MAPK/ERK pathway activity and BRAF mutations while the majority, 70–80%, is thought to develop through the adenoma pathway [17]. To investigate if MAPK-ERK signaling was also increased in the BMPR1AΔ−Col1a2 mice, a staining was performed for phosphorylated ERK (pERK). While pERK positive cells were only found to be present in the (lower) crypt region of the intestines from BMPR1Actrl−Col1a2, positive cells were also found to be present in the top of the villi, polyps, and colonic glands at the luminal side of the colon from BMPR1AΔ−Col1a2 mice (Fig. 3A–E). Next, an IHC staining for B-catenin was performed to investigate changes in Wnt pathway activity. Increased cytoplasmic and membranous staining was observed specifically in the polyps but no increased nuclear translocation was observed in both the small intestine and colon of the BMPR1AΔ−Col1a2 mice (Fig. 3F–J).
Fig. 3
Loss of BMPR1A signaling in fibroblasts results in increased pERK levels and increased TGF-β activity. A–E pERK staining in the intestines of BMPR1Actrl−Col1a2 is only observed in the (lower) crypt part of the small and large intestines. However, pERK positive cells were present at the luminal side of the intestines from BMPR1A∆−Col1a2mice. F–J Despite the increased membranous and cytoplasmatic B-catenin staining in the epithelial cells within the polyp, no increased nuclear localization was observed in tissue from BMPR1A∆−Col1a2mice compared to control. K, L Quantification of an immunohistochemical staining for TGF-β showed significant increased TGF-β in the colon of BMPR1A∆.−Col1a2mice. M, N An ELISA for TGF-β confirmed the increased TGF-β observed in the IHC staining. O–U The staining for pSMAD2 verified increased TGF-β signaling.) Bars represent mean ± SD. P < 0.05 (*)
Since the abrogation of BMP signaling in fibroblasts could augment a number of actions in fibroblasts, we performed additional stainings to assess TGF-β signaling. TGF-β is known to be an important growth factor secreted by fibroblasts that regulates fibroblast activation and phenotypic transition into myofibroblasts [18]. Increased TGF-β staining was observed in the intestines of BMPR1AΔ−Col1a2 mice compared to BMPR1Actrl−Col1a2 mice. However, this increase was only found to be significant in the colon of the BMPR1AΔ−Col1a2 mice (Fig. 3K–L and Supplementary Fig. 6A–E). Next, a TGF-β ELISA on whole tissue homogenates was performed and confirmed the increased levels of TGF-β (Fig. 3M, N). Given the fact that TGF-β is secreted as a latent molecule, we performed an additional IHC staining for phosphorylated SMAD2 (pSMAD2), a protein activated upon TGF-β signaling. pSMAD2 staining confirmed increased TGF-β activation in BMPR1AΔ−Col1a2 mice (Fig. 3O–U). Increased TGF- β activation did not result in an increased number of myofibroblasts as no differences in α-SMA expression were detected (Supplementary Fig. 6 J–Q).
To investigate if any epithelial mutations were present in the polyps, next-generation sequencing for common CRC hotspot genes including Apc, Smad4, B-raf, K-ras, N-ras, Pik3ca, Pten and Trp53 was performed. No somatic mutations were detected in 16 polyps that were analyzed. Taken together, these data indicate that the loss of BMPR1A in fibroblasts results in increased MAPK-ERK signaling and is associated with the serrated pathway of CRC development. The absence of a somatic suggests that increased ERK activation is probably induced by soluble factors secreted by the BMPR1AΔ fibroblasts.
CXCL12 is expressed by intestinal fibroblasts and upregulated in mice with fibroblast-specific BMPR1A knockoutAfter observing the increased epithelial cell growth, we assumed that this is probably caused by soluble factors specifically derived from BMPR1A KO fibroblasts. We, therefore, combined two publicly available human mRNA-sequencing data sets to identify factors secreted by the tumor stroma (GSE39395), which are also upregulated in serrated polyps (GSE45270) (Supplementary Fig. 7). 99 genes were found to be upregulated in both data sets. The exclusion of genes encoding for intracellular proteins, membrane proteins, and extracellular matrix proteins resulted in a selection of genes encoding for secreted factors (Supplementary Fig. 7B). Subsequent literature research resulted in a shortlist of 10 possible target genes that could influence epithelial cell growth. CXCL12 and EFEMP1 belonged to the top 3 most differentially expressed genes in both datasets. qPCR analyses on fibroblasts in which BMP signaling was inhibited, revealed upregulation of CXCL12 to be the strongest (Supplementary Fig. 7C, D), which was therefore selected to evaluate further.
To investigate if CXCL12 could indeed be implicated in epithelial cell growth, RNAscope in situ hybridization for Cxcl12 was performed on BMPR1AΔ−Col1a2 mouse tissue. Increased Cxcl12 staining was present in the small intestines, polyps, and colon of BMPR1A∆−Col1a2 mice compared to control mice (Fig. 4A–D). The membrane marker glycoprotein 38 (gp38) was used to visualize intestinal fibroblasts and showed that Cxcl12 colocalized in fibroblast-rich regions [19]. To further confirm that intestinal fibroblasts express CXCL12, we used tissue from Cxcl12-green fluorescent protein (GFP) transgenic mice in which cells that express CXCL12 also express GFP. Immunofluorescent visualization of GFP showed that CXCL12 is exclusively expressed by the stroma of both the small intestine and the colon (Fig. 4E, F). Immunofluorescent staining for cell-specific markers showed clear co-localization of GFP with endothelial cells (CD31) and general fibroblast markers Collagen-I and gp38, but not with immune cells (CD45) or lymphatic endothelial cells (Lyve1) (Supplementary Fig. 8A). These observations were further validated by performing Fluorescence-Activated Cell Sorting (FACS) on both the small intestine and colon for epithelial cells (EpCam+), immune cells (CD45+), endothelial cells (gp38− CD31+), fibroblasts (gp38+CD31−), and lymphatic cells (gp38+CD31+). The gating strategy is indicated in Supplementary Fig. 8B. Real-time quantitative PCR (RT-qPCR) analysis of sorted cells for Cxcl12 mRNA showed that intestinal Cxcl12 is only expressed by fibroblasts and endothelial cells. In the small intestines, the highest Cxcl12 expression was found in fibroblasts, followed by endothelial cells (Fig. 4G). The opposite pattern was found in the colon (Fig. 4H). Interestingly, in colorectal cancer (CRC), CXCL12 is also expressed most abundantly in endothelial cells, followed by fibroblasts (Fig. 4I). These data show that fibroblasts produce CXCL12 and that expression is increased after the loss of BMPR1A signaling, suggesting that CXCL12 might be involved in the observed phenotype in the BMPR1AΔ−Col1a2 mice.
Fig. 4
Fibroblast CXCL12 is upregulated in BMPR1A∆−Col1a2. Combining two online available mRNA-expression data sets (GSE45270 and GSE39395) identified CXCL12 as a soluble factor upregulated in CRC tumor stroma and serrated polyps. A–D RNAscope in situ hybridization for Cxcl12 and staining for the fibroblast marker gp38 showed an apparent increase of Cxcl12-expressing fibroblasts in BMPR1A∆−Col1a2 intestinal tissue compared to tissue from BMPR1Actrl−Col1a2. E, F Immunofluorescent visualization of GFP expressed in intestinal tissue from CXCL12-GFP mice showed that it was only present in the stroma of the small intestine and colon. G-H) qPCR for Cxcl12 on cell sorted epithelial cells, immune cells, fibroblasts, lymphatic endothelial cells, and endothelial cells isolated from wild-type B6 mice showed that Cxcl12 is exclusively expressed by fibroblasts and endothelial cells. I Human CRC expression data from GSE39396 showed that CXCL12 is mainly expressed by fibroblasts and endothelial cells and follows the same expression pattern as observed for mouse colonic tissue. Bars represent mean ± SD. P < 0.05 (*)
BMP signaling regulates CXCL12 expressionTo investigate how the BMP pathway affects fibroblast CXCL12 expression, several experiments were performed using the human colon fibroblast CCD-18co. Firstly, stimulation of 18co fibroblasts for 24 h with recombinant BMP2 resulted in a significant decrease of CXCL12 expression (Fig. 5A, P = 0.002). This could be restored by adding LDN-193189, a selective BMPR1A inhibitor. To mimic the long-term effect of loss of BMP signaling, cells were exposed to LDN-193189 for up to 96 h. CXCL12 expression gradually increased over time, reaching a 10.5-fold increase after 96 h (Fig. 5B). Expression of the BMP target gene ID1 was downregulated after 24 h and remained downregulated up to 96 h (Fig. 5C), showing that BMP signaling was successfully inhibited over the time course of the experiment. Stimulation of human primary colonic fibroblasts (pFibro) isolated from normal colon tissue for 96 h with LDN-193189 also resulted in increased CXCL12 expression (Fig. 5D), confirming the findings with the CCD-18co fibroblasts. Besides CXCL12, inhibition of BMP signaling was also found to significantly alter expression of WNT2A, TGF-β2 and TGF-β3, but not TGF-β1 and A-SMA,(Supplementary Fig. 6H–K and S).
Fig. 5
CXCL12 expression is regulated by BMP activity and stimulates epithelial proliferation. A Stimulation of CCD-18co fibroblasts with 100 ng BMP2 for 24 h resulted in a significant downregulation of CXCL12 expression. Downregulation was prevented when 200 nM LDN-193189, a BMPR1A inhibitor, was present in the medium. B Stimulating CCD-18co fibroblasts or primary colonic fibroblasts for up to 96 h with 200 nM LDN-193189 showed a gradual increase of CXCL12 expression over time. C Expression of ID1, a BMP target gene, decreased, showing that BMP signaling was successfully inhibited. D Similar findings were observed when human primary fibroblasts were treated with LDN-193189. E, F The presence of 200 nM LDN-193189 was found to lower the invasive capacity of fibroblast while also significantly hindering the migratory capacity of the fibroblasts. G–J Immunohistochemical staining of BMPR1A∆−Col1a2 intestinal tissue for Gremlin showed a clear increase compared to the intestines from control mice. K–L An ELISA for Gremlin confirmed the increased levels in intestines from BMPR1A∆−Col1a2 mice. M, N Treatment of normal intestinal organoids with recombinant CXCL12 showed that it CXCL12 stimulates organoid growth significantly. Bars represent mean ± SD except for (I) in which error bars represent mean ± SEM. P < 0.05 (*), < 0.01 (**), < 0.001 (***) and < 0.0001 (****)
To further evaluate how inhibition of BMP signaling would influence the functional behavior of fibroblasts, transwell invasion assays were performed. These data showed that inhibition of BMP signaling in fibroblasts with LDN-193189 resulted in decreased migratory and invasive capacities of the fibroblasts (Fig. 5E, F).
BMP antagonists are known to regulate the BMP signaling amplitude in vivo. To investigate whether BMP antagonists can also influence CXCL12 expression, CCD-18co cells were stimulated for 96 h with the BMP antagonist Noggin. This resulted in ID1 downregulation and concomitant CXCL12 upregulation (Supplementary Fig. 9). To explore if the overexpression of BMP antagonists could explain increased Cxcl12 expression in BMPR1A∆−Col1a2 mice in vivo, immunohistochemical stainings were performed for the BMP antagonist Gremlin. Gremlin binds the same BMP ligands, but unlike Noggin, it is almost exclusively expressed by fibroblasts (Supplementary Fig. 10A, B). A substantial increase in Gremlin expression (Fig. 5G–J) in the small and large intestines of the BMPR1AΔ−Col1a2 was observed. This was also confirmed in polyps of the BMPR1A∆−SM22 mice (Supplementary Fig. 11). ELISA analysis for Gremlin confirmed the increased Gremlin expression in BMPR1AΔ−Col1a2 mice (Fig. 5K, L). These data show that the loss of BMPR1A signaling and increased BMP antagonist expression might be instrumental for increased CXCL12 expression.
To investigate whether CXCL12 indeed stimulates cell growth, we stimulated intestinal mouse organoids with recombinant CXCL12. The organoids were significantly larger from day 2 onwards, compared to unstimulated organoids (Fig. 5M, N). We examined if treatment of organoids with CXCL12 regulated expression of serrated-associated genes Anxa1, Pdx1 (shown to be upregulated in serrated polyps) and Cdx1 (downregulated in serrated polyps). Treatment of organoids with CXCL12 did not result in significant changes in expression of Anxa1, Cdx1 and Pdx1 (Supplementary Fig. 12A). Together these data show that loss of BMP signaling can increase CXCL12 expression, and in turn, CXCL12 can drive epithelial cell growth.
Treatment of mice with the BMP activator FK-506 or CXCL12 neutraligand LIT-927 abrogates polyp developmentNext, we investigated if polyp development in the BMPR1AΔ−Col1a2 mice could be abrogated by restoring BMP activity with FK-506, a ligand-independent activator of the BMP pathway. Restoring BMP signaling significantly reduced polyp formation by 45% from 20.8 polyps per mouse in the vehicle control group to an average of 11.4 per mouse in the FK-506 treated group (P = 0.0013, Fig. 6A, B and E). To confirm the role of CXCL12 in polyp development, BMPR1AΔ−Col1a2 mice were treated with LIT-927, a CXCL12 neutralizing ligand. A substantial 75% reduction in the number of polyps from 30.3 per mouse to 7.5 per mouse was found for mice treated with LIT-927 compared to mice treated with vehicle (P = 0.0018, Fig. 6C, D and F). Next to the decreased number of polyps in the small intestine, LIT-927 normalized the increased the crypt length in the large intestine of treated mice (mean = 817.9 µm, P = 0.0054, Fig. 6L) compared to vehicle-treated mice (mean = 1125 µm), while FK-506 was not capable of restoring the increased crypt length (Fig. 6K). These data suggest that inducing BMP activity with FK-506 or inhibiting CXCL12 signaling due to neutralization of CXCL12 can significantly prevent polyp development in BMPR1AΔ−Col1a2 mice.
Fig. 6
Treatment of mice with the BMP activator FK-506 or the CXCL12 neutralizer LIT-927 results in fewer polyps. A–D Treatment of mice with FK-506 or LIT-927 resulted in less polyps. E 1.8-fold fewer polyps were observed in FK-506 treated mice (mean = 11.4) compared to vehicle-treated mice (mean = 20.8). F fourfold fewer polyps were observed in LIT-927 treated mice (mean = 7.5) compared to vehicle control mice (mean = 30.3). G–L While the FK-506 treatment did not restore the crypt length, a significant decrease in crypt length was observed in mice treated with LIT-927. M, N Treatment of mice with the compounds resulted in a decreased number of olfm4 positive cells per crypt. This was however only significant in mice treated with FK-506. O–R The number of Ki67 positive cells showed a decreased trend in mice treated with LIT-927 (not significant). S–V Treatment of mice with one of the compound did not result in an increase of cells that have undergone apoptosis as assessed with an IHC for cleaved caspase 3. Bars represent mean ± SD. P < 0.05 (*) and < 0.01 (**)
Next, we investigated the effect of the treatments on the number of Olfm4, Ki67, CD45 and cleaved caspase 3 positive cells. The number of Ki67 positive cells and percentage of CD45 positive cells seemed to decrease in the treated mice, although this was not significant (Fig. 6O–R and Supplementary Fig. 13 and 14). In contrast, significantly less Olfm4 positive cells per crypt were observed in mice treated with FK-506 (Fig. 6M). A similar decrease was observed for mice treated with LIT-927 but due to the increased variation between mice this was found to be not significant (Fig. 6N). No significant changes in the number of apoptotic cells were observed based on the cleaved caspase 3 staining (Fig. 6S–V).
CMS4 colorectal cancers express high levels of BMP antagonists and CXCL12To investigate if decreased BMP signaling is also implicated in the development of human serrated polyps, publicly available human mRNA-sequencing data sets were used in which the expression profile of serrated polyps are compared to tubular adenomas (GSE45270). GREMLIN1 expression was significantly upregulated, while the inhibitor of DNA binding (ID) proteins ID1, ID2, ID3 and ID4, all major downstream transcription targets of BMP signaling, were found to be significantly downregulated in serrated adenomas compared to tubular adenomas. This suggests that BMP activity is indeed decreased in serrated adenomas (Fig. 7A–E).
Fig. 7
CXCL12 and the BMP antagonists GREMLIN1 and NOGGIN are significantly upregulated in CMS4 tumors. A–E Serrated adenomas express GREMLIN1 compared to tubular adenomas. Several ID proteins were found to be lower expressed in serrated adenomas. F–G Analyzing a colon adenocarcinoma TCGA cohort (n = 155) for GREMLIN1 and CXCL12 showed that patients with high GREMLIN1 or CXCL12 had a significantly poorer survival compared to patients with low expression. H Stratifying the 155 samples for both CXCL12 and GREMLIN1, showed that particularly patients with high expression of both CXCL12 and GREMLIN1 had a significantly worse overall survival. I High GREMLIN1 expression showed a significant association with higher CXCL12 expression J–L Stratification of colorectal TCGA cases (n = 240) into the four CMS subtypes showed that GREMLIN1, NOGGIN and CXCL12 are significantly upregulated in CMS4 tumors. M, N High GREMLIN1 and NOGGIN were significantly associated with higher CXCL12 levels. Bars represent mean ± SD. P < 0.05 (*), < 0.01 (**), < 0.001 (***) and < 0.0001 (****)
Next, to study the relation between BMP antagonist expression and CXCL12 in humans, a publicly available the cancer genome atlas (TCGA) mRNA-expression data set from a colon adenocarcinoma cohort with survival data consisting of 155 patients was used. High GREMLIN1 and high CXCL12 expression were associated with a significantly poorer overall survival (P = 0.04 and P = 0.024 respectively, Fig. 7F–G). While a similar pattern was observed for NOGGIN in the first 24 months of follow-up, this did not reach statistical significance (P = 0.116, Supplementary Fig. 15A). High GREMLIN1 levels were associated significantly (P ≤ 0.0001) with higher CXCL12 expression while NOGGIN was not (Fig. 7I and Supplementary Fig. 15B). Moreover, patients with tumors expressing high levels of both GREMLIN1 and CXCL12 had a significantly worse survival (P = 0.021) compared to patients with high expression of only CXCL12, GREMLIN1, or NOGGIN (Fig. 7H). Similar trends were observed for NOGGIN, but these did not reach statistical significance (P = 0.095, Supplementary Fig. 15C).
Since the BMP antagonists are expressed by stromal cells in the colon, we hypothesized that the BMP-CXCL12 signaling axis would be of particular significance in tumors with an abundant stromal compartment. The Consensus Molecular Subtype (CMS) classification system subdivides CRCs into four subgroups based on the transcriptional signature to which both cancer cells and the tumor stroma contribute [16]. The CMS4 tumors are known as the mesenchymal subtype because of gene signatures consistent with activated tumor stroma, such as active TGF-β signaling. We used TCGA RNA sequence data from sporadic CRC patients to look at the expression of GREMLIN1, NOGGIN, and CXCL12 between the 4 different CMS subgroups. Both the BMP antagonists and CXCL12 were more highly expressed in CMS4 compared to CMS1-CMS3 (Fig. 7J–L). Dividing patients into GREMLIN1 low and GREMLIN1 high expressing subgroups or NOGGIN low and NOGGIN high expressing subgroups showed that patients with high GREMLIN1 or NOGGIN also had significantly higher CXCL12 expression (Fig. 7M, N). These data together support the existence of a tumor-promoting BMP-CXCL12 signaling axis.
留言 (0)