
記住我
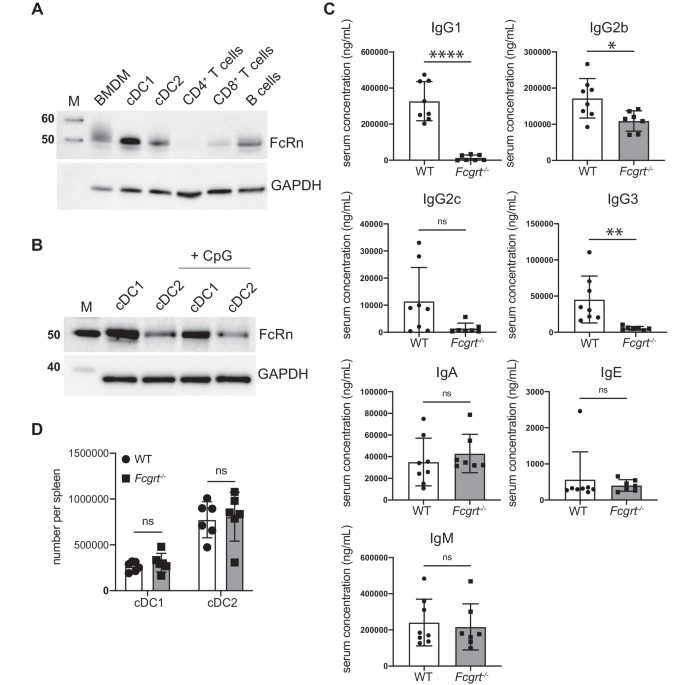
Previous studies11,26,44 in both animals and humans45,46, demonstrated that HA-Fe-nanoparticles elicit antibody responses of significantly enhanced breadth and magnitude relative to currently licensed inactivated influenza vaccines. However, the antigen specificity and magnitude of the CD4 T cell response to HA-Fe-nanoparticle immunization remains poorly understood. We sought to empirically determine the relative CD4 T cell immunodominance hierarchy of the ferritin vs. HA components of the HA-Fe-nanoparticles. To investigate the primary CD4 T cell response to HA-nanoparticle immunization, two mouse stains (BALB/c and CBA/J) were chosen based on their expression of 4 alternate MHC class II proteins of two distinct allelic forms of I-E and I-A, to recruit as diverse a CD4 T cell repertoire as possible. Mice were immunized once with HA-Fe-nanoparticle via a subcutaneous route in the presence of Sigma Adjuvant System® that elicits both IL-2 and IFNγ producing CD4 T cells47. CD4 T cell responses were quantified 10 days post immunization by peptide-stimulated cytokine ELISpot assays. To determine the epitope specificity of the CD4 T cell response at the level of single peptides, a peptide library of 15-mer peptides overlapping by 11 residues covering the sequence of H. pylori ferritin (Table 1) was used. To test CD4 T cell reactivity directed against HA, we tested an array of major peptide epitopes from A/New/Caledonia/99 HA that had been previously defined by overlapping peptide matrices spanning the full sequence of HA in mouse strains expressing different MHC class II haplotypes (Table 2)48,49,50,51,52.
Table 1 MHC Class II-restricted H. pylori ferritin epitopes in H-2d, H-2K, HLA-DR1, and HLA-DR4 mice.Table 2 MHC Class II-restricted A/New Caledonia/99 hemagglutinin epitopes in H-2d, H-2K, HLA-DR1, and HLA-DR4 mice.The relative CD4 T cell immunodominance hierarchy of ferritin and HA in these experiments was scored by using a cutoff of 25 spots per million CD4 T cells as a criterion to define reproducible CD4 T cell specificities, indicated by a horizontal dotted line in Fig. 1. In BALB/c mice, expressing the MHC class II molecules I-Ad and I-Ed, the elicited CD4 T cells exhibited a striking immunodominance bias for ferritin-derived peptides, as measured by IL-2 (Fig. 1A) and IFNγ ELISpot assays (Fig. 1B). When assessing IL-2 production in response to peptide stimulation, ferritin peptides FE49, FE53, FE101, and FE105 elicited robust responses of greater than 100 IL-2 spots per million CD4 T cells (Fig. 1A). In contrast only 4 of the 13 HA-derived peptides, HA120, HA126, HA180, and HA209, elicited a response of greater than 25 spots per million CD4 T cells (Fig. 1A). When assessing IFNγ production in HA-nanoparticle immunized mice, ferritin peptides FE49, FE53, FE101, and FE105 again elicited responses of several hundred IFNγ spots per million CD4 T cells (Fig. 1B). None of the HA peptides tested elicited responses greater than 25 spots (Fig. 1B). In CBA/J mice expressing the MHC class II molecules I-Ak and I-Ek, HA-Fe-nanoparticle immunization elicited a CD4 T cell response that was biased towards the ferritin nanoparticle core, with HA-derived peptides being sub-dominant as measured by IL-2 (Fig. 1C) and IFNγ ELISpot assays (Fig. 1D). CD4 T cells stimulated with ferritin peptides FE37, FE41, FE45, FE101, FE141, FE145, and FE149 each elicited robust responses (Fig. 1C). Of the 8 HA-derived peptides tested, only peptides HA120, HA304, HA358, and HA398 elicited responses of greater than 25 spots per million IL-2 producing CD4 T cells (Fig. 1C), similar to the immunodominance observed when IFNγ producing cells were quantified. Ferritin peptides FE37, FE41, and FE45 elicited responses of greater than 1000 spots per million CD4 T cells, and responses of greater than 25 spots per million for peptides FE101, FE141, FE145, and FE149 (Fig. 1D). None of the HA peptides elicited IFNγ responses of greater than 25 spots per million CD4 T cells (Fig. 1D).
Fig. 1: Defining immunodominance of the CD4 T cell repertoire in BALB/c and CBA/J mice expressing diverse I-A and I-E alleles.
Identification of immunodominant ferritin epitopes was performed by IL-2 and IFNγ cytokine ELISpot following immunization with full length HA-ferritin nanoparticles. Purified CD4 T cells were restimulated with peptides spanning the sequence of H. pylori ferritin and major HA peptides. Experiments were conducted in A, B BALB/C mice (MHC II I-Ad/Ed) and C, D CBA mice (MHC II I-Ak/Ek). The fraction of the cytokine-producing CD4 T cell response specific for ferritin (black) or HA (white) is shown for E BALB/c and F CBA/J mice, respectively. Data are shown as the mean and SD of two independent experiments of five pooled mice per group.
When assessing summed responses by antigen specificity, 85% of the IL-2 response and 98% of the IFNγ response in BALB/c mice were directed against ferritin-derived peptides (Fig. 1E). In contrast, only 15% of the IL-2 response and 2% of the IFNγ response in BALB/c mice were directed against HA (Fig. 1E). When the relative magnitudes of the responses were assessed for CBA/J mice, 88% of the IL-2 response and 99% of the IFNγ response were directed against ferritin-derived peptides (Fig. 1F), while only 12% of the IL-2 response and 1% of the IFNγ response were directed against HA (Fig. 1F). Given the nature of the overlapping peptide library used to define reactivity against H. pylori ferritin, it is possible that adjacent peptides tested by ELISpot represent a single peptide specificity. The peptides tested, listed in Table 1 and Table 2, were overlaid on the sequence of HA-Fe nanoparticles in Supplementary Fig. 1. For both BALB/c and CBA/J mice, regions of overlap between adjacent major peptides, potentially representative of the core binding motif, are underlined.
To confirm that CD4 T cell reactivity to H. pylori ferritin was due to HA-Fe-nanoparticle immunization and not murine Helicobacter infection, splenocytes of naïve CBA/J mice were tested for reactivity to immunodominant ferritin peptides by cytokine ELISpot and compared to HA-nanoparticle immunized controls. Reactivity to ferritin peptides, as measured by IL-2 and IFNγ production, was only observed in HA-nanoparticle immunized mice (Supplementary Fig. 2). In addition, all animals used in these studies were housed in a Helicobacter negative space where mice routinely tested negative for murine Helicobacter by PCR. Thus, the ferritin reactivity detected by ELISpot was due to elicitation of the CD4 T cells after the vaccination.
Assessing HLA-DR restricted CD4 T cell responses to HA-nanoparticle vaccination and ferritin-specific CD4 T cell responses in healthy human donorsGiven the unexpected immunodominance of ferritin-specific CD4 T cells in two common laboratory mouse strains with distinct MHC class II molecules expressed, we next sought to determine the immunodominance hierarchies of ferritin and HA in the context of human MHC class II molecules. Transgenic mice expressing HLA-DR1 or HLA-DR4, class II molecules commonly expressed in humans, were immunized with HA-Fe-nanoparticles, and CD4 T cell responses were assayed using cytokine ELISpot assays. Cells were stimulated with overlapping peptides spanning the sequence of H. pylori ferritin or previously defined major peptide epitopes from A/New/Caledonia/99 HA48. Even in HLA-DR1 transgenic mice, where we previously discovered an exceptionally high number of CD4 T cell epitopes in HA51, strong reactivity to ferritin-derived peptides was observed, where ferritin peptides FE1, FE5, and FE97 elicited readily detectable responses (Fig. 2A). Reactivity of CD4 T cells post vaccination was also observed against HA peptides HA162, HA203, and HA440 (Fig. 2A). These results suggest that ferritin maintains its CD4 T cell immunodominance even in the context of a robust HA-specific response in HLA-DR1 mice. In HLA-DR4 transgenic mice, robust CD4 T cell reactivity in the primary responses was observed in response to stimulation with ferritin peptides FE12, FE25, FE29, FE37-FE41, FE97-FE105, FE141, and FE149 (Fig. 2B). Responses to defined HA peptides HA203 and HA328 were modest (Fig. 2B) indicating that the HLA-DR4 restricted CD4 T cell response is highly biased towards ferritin. When ELISpot results were summed by antigen specificity across the murine MHC class II and the human MHC class II alleles sampled, a consistent pattern of ferritin-biased immunodominance is readily apparent (Fig. 2C).
Fig. 2: Defining immunodominance of the CD4 T cell repertoire in human MHC-II haplotypes.
Identification of immunodominant ferritin epitopes was performed by IL-2 cytokine ELISpot following immunization with full length HA-ferritin nanoparticles. Purified CD4 T cells were restimulated with peptides spanning the sequence of H. pylori ferritin and major HA peptides. Experiments were conducted in transgenic mouse models expressing A HLA-DR1 and B HLA-DR4 human MHC class II alleles. Data are shown as the mean and SEM of three independent experiments of five pooled mice per group. C Summed cytokine response by antigen specificity across murine and human MHC class II molecules used in Figs. 1 and 2. D PBMC from 36 healthy human donors with unknown history of H. pylori infection were stimulated with a peptide pool spanning the entire sequence of H. pylori ferritin and plated in IL-2 and IFNγ cytokine ELISpot. Information about human donors is presented in Table 3. Data are shown as the mean and SD of the subjects assayed.
We then extended our analysis to sample healthy human donors that express a diverse array of HLA class II molecules. Because humans can be exposed to H. pylori antigens via infection53,54, we asked whether circulating CD4 T cells displayed detectable reactivity to ferritin-derived epitopes that could theoretically be called into the response to immunization with vaccine constructs containing H. pylori ferritin. Peripheral blood mononuclear cells (PBMC) from human donors (Table 3) with unknown exposure to Helicobacter were depleted of CD8 + cells and CD56 + cells, and PBMCs were stimulated with pooled ferritin peptides spanning the sequence of H. pylori ferritin in IL-2 and IFNγ ELISpot assays (Tables 1 and 2). These experiments revealed that for both IL-2 and IFNγ, responses of greater than 25 spots per million depleted PBMC are detectable in approximately 15% of the human donors assayed (Fig. 2D). This reactivity to H. pylori ferritin, likely due to previous infection with Helicobacter, supports the hypothesis that a subset of human donors has H. pylori ferritin-specific CD4 T cells that have the potential to be recalled in response to immunization with ferritin-based vaccine constructs. We observed intra-donor variability in the relative production of IL-2 and IFNγ in response to ferritin epitopes. This may be due to heterogeneity in the responding CD4 T cell phenotypes, where H. pylori infection has been described to elicit Th1, Th17, and regulatory CD4 T cells53,54. These data suggest that both murine and diverse human MHC class II molecules select for and present H. pylori ferritin-derived peptides to ferritin-specific CD4 T cells.
Table 3 Demographics and characteristics of subjects in study.Ferritin-specific CD4 T cells participate in the germinal center response to HA-nanoparticle immunizationIn the context of limiting CD4 T cell help, such as vaccination during the primary response lacking CD4 T cell helper memory cells or after vaccination with proteins with low abundance of CD4 T cell epitopes, we hypothesized that the direct linkage of ferritin to HA could contribute to anti-HA B cell responses. To test this hypothesis, BALB/c and CBA/J mice were immunized with HA-Fe-nanoparticles via a subcutaneous route, as before, and CD4 T cell and B cell responses were sampled 15 days post primary vaccination in the draining popliteal lymph node (LN). In these studies, we compared the responses to parallel vaccination of a separate cohort of mice with an early-generation vaccine construct consisting of an HA trimer stabilized with a fold-on domain55 that lacked the ferritin core. Post-vaccination, CD4 T cells were stimulated in IL-2 ELISpot assays with peptide pools spanning the complete sequence of HA or FE in order to broadly capture the responding cells. Robust cytokine producing cells were detected in response to stimulation with ferritin peptides in both BALB/c and CBA/J (Fig. 3A, B), in agreement with our earlier data. When the abundance of TFH (Supplementary Fig. 3A, defined as CD4+ CD44++ PD1++ CXCR5++) was assessed, HA-Fe-nanoparticle immunized mice had a significantly increased abundance of TFH per LN relative to HA-trimer immunized mice (Fig. 3C, D). BALB/c and CBA/J mice immunized with HA-FE-nanoparticles had 3.0 and 4.9-fold increases, respectively, in absolute TFH abundance relative to HA-trimer immunized mice (Fig. 3C, D). To determine the antigen-specificity of the TFH from the draining LN, cells were stimulated with HA or FE peptide pools and a non-cytokine dependent method to quantify the antigen-reactive CD4 T cells was utilized. Activation Induced Marker (AIM) assays that track upregulation of the markers CD154 (CD40L) and CD69 (Supplementary Fig. 3B) were used as readouts of antigen-dependent CD4 T cell activation56,57,58. These analyses revealed ferritin-specific TFH to be the most abundant subset in HA-nanoparticle immunized BALB/c and CBA/J mice (Fig. 3E, F). As a fraction of activated TFH detected by the AIM assay, ferritin-specific CD4 TFH represented 85% of the responding cells in BALB/c and 87% of responding cells in CBA/J (Fig. 3G-H). While activation of HA-specific TFH was detectable, the immunodominance of ferritin was maintained in the TFH repertoire (Fig. 3E–H). Given that non-TFH can also provide help to B cells through CD40L signals41,59, the specificity of the antigen-experienced CD44 high CD4 T cell repertoire was assessed. These studies indicated that the immunodominance of ferritin among CD44 high CD4 T cell is preserved both in terms of absolute abundance and fraction of the antigen-specific responses (Fig. 3I–L). Collectively, these data on CD4 T cell reactivity and specificity indicate that the immunodominance of ferritin is maintained among cytokine-producing cells (Fig. 1), CD4 TFH repertoire (Fig. 3C–H), and antigen-experienced CD4 T cells with the potential to upregulate the essential co-stimulatory molecule CD40L (Fig. 3I–L)60.
Fig. 3: Ferritin-specific CD4 T cells participate in the germinal center response to HA-nanoparticle immunization.
BALB/c and CBA/J mice were immunized with equimolar quantities of HA in the form of soluble HA trimers or HA-nanoparticles. Draining lymph nodes were harvested D15 post immunization. Antigen-specific CD4 T cell responses were quantified by IL-2 ELISpot for A BALB/c mice and B CBA/J mice following stimulation with total HA or total ferritin peptide pools. Absolute numbers of CD4 TFH per lymph node were quantified for C BALB/c and D CBA/J. Antigen specificity of the TFH response was quantified by activation induced marker (AIM) assay by scoring upregulation of CD154 and CD69 in response to peptide stimulation for E BALB/c and F CBA/J. The fraction of the AIM+ TFH specific for ferritin (black) or HA (white) is shown for G BALB/c and H CBA/J. Quantification of antigen experienced non-TFH that upregulate CD154 and CD69 in response to peptide stimulation for I BALB/c and J CBA/J. The fraction of the AIM+ antigen experienced CD4 T cell responses specific for ferritin (black) or HA (white) is shown for K BALB/c and L CBA/J. Data are shown as the mean and SD three individual mice per group from three independent experiments, for a total of 9 individual mice per group. In A, B, E, F, I, J, significant differences between HA-trimer and HA-nanoparticle immunized mice were determined by two-way ANOVA with Tukey’s correction for multiple comparisons. In C, D, significant differences between HA-trimer and HA-nanoparticle immunized mice were determined by unpaired, two-tailed Mann-Whitney test.
Given the dependence of germinal center formation on TFH, we hypothesized that if CD4 T cell help was in fact a limiting element in the B cell response, the HA-Fe-nanoparticle vaccinated mice would display an increased abundance of GC B cells relative to HA-trimer vaccinated mice. We found that in both BALB/c and CBA/J mice, the vaccine-draining LN of the HA-Fe-nanoparticle vaccinated mice had significantly increased overall abundance of GC B cells (Supplementary Fig. 3A, defined as B220+ FAS+ GL7+) relative to that exhibited by the HA-trimer vaccinated mice (Fig. 4A, B), which ranged from 2.8-fold in BALB/c and 4.6-fold in CBA/J mice. Linear regression analyses demonstrated the presence of a strong, positive correlation between the overall abundance of GC B cells and TFH. The slope of the regression line was significantly positive, suggesting that increased TFH recruitment contributes to enhanced GC B cell responses in both BALB/c and CBA/J mice (Fig. 4C, D).
Fig. 4: HA-nanoparticle immunization elicits enhanced antigen-specific B cell responses that correlate strongly with abundance of TFH.
Total abundance of FAS+ GL7+ germinal center B cells per lymph node was assessed in A BALB/c and B CBA/J mice immunized with equimolar quantities of HA in the form of soluble HA trimers or HA-nanoparticles from the same individual mice shown in Fig. 3. The relationship between germinal center B cell abundance and CD4 TFH abundance was assessed by linear regression analysis for C BALB/c and D CBA/J mice immunized with HA trimers or HA-nanoparticles. When cohorts vaccinated with HA-trimers and HA-Fe-nanoparticles are separately analyzed by linear regression analyses, the same trend is observed. Antigen-specific GC B cell responses were assessed by staining with fluorophore conjugated HA probes, where the B cell population of interest was defined as FAS+ GL7+ IgM- IgD- HA+ for E BALB/c and F CBA/J mice. HA-specific ASCs were assessed by B cell ELISpot. The frequency of HA-specific IgG isotype B cells was assessed for G BALB/c and H CBA/J mice. Data are shown as the mean of three individual mice per group from three independent experiments, for a total of 9 individual mice per group. In (A, B, E, F, G, H), significant differences between HA-trimer and HA-nanoparticle immunized mice were determined by unpaired, two-tailed Mann-Whitney test.
In order to address the issue of antigen specificity in the GC B cells, fluorescently labelled full-length HA probes were used to detect HA-specific B cells55 (Fig. 4E, F). HA-Fe-nanoparticle immunized BALB/c and CBA/J mice have significantly increased overall abundance of HA-specific germinal center B cells (Supplementary Fig. 3A, defined as B220+ FAS+ GL7+ HA-probe+) relative to the HA-trimer immunized cohort (Fig. 4E, F). Increased HA-specific germinal center B cell abundance ranged from 3.7-fold in BALB/c and 3.8-fold in CBA/J (Fig. 4E, F). Finally, we assessed the ASC response elicited by HA-trimer and HA-Fe-nanoparticle immunization with antigen-specific B cell ELISpot. There was a significantly increased frequency of HA-specific IgG isotype ASCs in HA-nanoparticle immunized mice relative to HA-trimer immunized mice, where there was an 11-fold increase in BALB/c and 31-fold increase in CBA/J mice (Fig. 4G, H). Recent studies have demonstrated that a robust isotype switched ASC response is dependent on CD40L signaling from CD4 T cells59, such as those depicted in (Fig. 3E–L), implicating the increased abundance of ferritin-specific helper CD4 T cells in enhanced magnitude of the B cell response.
Equivalent vaccine responses to HA-trimers and HA-Fe-nanoparticles in H-2b MHC II haplotype mice lacking major CD4 T cell epitopesAlthough the results above show an enhanced magnitude of B cell and CD4 T cell responses in BALB/c and CBA/J mice immunized with HA-Fe nanoparticles relative to HA-trimers may be attributed to the recruitment of ferritin-specific CD4 T cells, there are other advantages of the HA-Fe-nanoparticle, including the multivalent nature of 8 HA trimers available to engage a broad diversity of HA specific B cells11,35,36,37,61. To address whether the primary advantages of the Fe-nanoparticle based vaccine in the developing germinal center response was related to its multimeric state, we sampled an additional inbred mouse strain C57BL/6 (B6) mice expressing the I-Ab class II molecule. Using a vaccination regimen as described above for BALB/c and CBA/J mice, we found that B6 mice elicited minimal CD4 T cell reactivity to H. pylori ferritin. No epitopes elicited a response of greater than 25 spots per million CD4 T cells in HA-nanoparticle immunized C57BL/6 mice by either IL-2 or IFNγ cytokine ELISpot (Supplementary Fig. 4). Restimulation of cells with complete peptide pools spanning the sequences of HA and ferritin elicited nearly undetectable CD4 T cell responses (Fig. 5A), in line with previous data showing very limited I-Ab restricted CD4 T cell reactivity to A/New Caledonia/20/99 HA62.
Fig. 5: Equivalent vaccine responses to HA-trimers and HA-nanoparticles in H-2b MHC II haplotype mice lacking major CD4 T cell epitopes.
C57BL/6 mice were immunized with equimolar quantities of HA in the form of soluble HA trimers or HA-nanoparticles. Draining lymph nodes were harvested D15 post immunization. Antigen-specific CD4 T cell responses were quantified by IL-2 ELISpot for A following stimulation with total HA or total ferritin peptide pools. Absolute numbers of CD4 TFH per lymph node were quantified B. Antigen specificity of the TFH response was quantified by activation induced marker (AIM) assay by scoring upregulation of CD154 and CD69 in response to peptide stimulation C. Quantification of antigen experienced non-TFH that upregulate CD154 and CD69 in response to peptide stimulation D. Total abundance of FAS+ GL7+ GC B cells per lymph node was assessed in E mice immunized with equimolar quantities of HA in the form of soluble HA trimers or HA-nanoparticles from the same individual mice shown in Fig. 5. The relationship between GC B cell abundance and CD4 TFH abundance was assessed by linear regression analysis F. Antigen-specific GC B cell responses were assessed by staining with fluorophore conjugated HA probes, where the B cell population of interest was defined as FAS+ GL7+ IgM- IgD- HA+ G. HA-specific ASCs were assessed by B cell ELISpot H. Data are shown as the mean and SD of three individual mice per group from two independent experiments, for a total of 6 individual mice per group. In A, C, D, significant differences between HA-trimer and HA-nanoparticle immunized mice were determined by two-way ANOVA with Tukey’s correction for multiple comparisons. In B, E, G, H, significant differences between HA-trimer and HA-nanoparticle immunized mice were determined by unpaired, two-tailed Mann-Whitney test.
We hypothesized that if the recruitment of Fe-specific CD4 T cells was a key factor in the overall immunogenicity of HA-Fe-nanoparticles, the HA-Fe-nanoparticle and HA-trimer based vaccines should elicit a similar magnitude of HA-specific B cell responses, whereas if the multimeric state was the critical advantage provided by HA-Fe-nanoparticles, this advantage would persist in B6-vaccinated mice. The analyses of CD4 T cells isolated from HA-trimer and HA-Fe-nanoparticle immunized B6 mice contained similar overall numbers of CD4 TFH per draining lymph node (Fig. 5B). Quantification of activated CD4 TFH and antigen-experienced CD4 T cells by AIM assay indicated very modest CD4 T cell responses (Fig. 5C, D). The number of GC B cells per lymph node was not significantly different between HA-trimer and HA-Fe-nanoparticle immunized B6 mice (Fig. 5E). Linear regression analyses showed a weak positive relationship between the abundance of GC B cells and CD4 TFH, but the slope was not significantly different than zero (Fig. 5F). HA-trimer and HA-Fe-nanoparticle immunized mice did not differ in their overall abundance of HA-specific isotype-switched GC B cells (Fig. 5G), nor in their frequency of HA-specific ASCs (Fig. 5H). These results indicate that B6 mice expressing the MHC II molecule I-Ab elicit limited CD4 T cell responses specific for HA or ferritin, and under these conditions, HA-trimer and HA-Fe-nanoparticle vaccine formulations elicited responses of equivalent, albeit lower overall magnitude, likely due to limited CD4 T cell help. Overall, these results suggest that a key component of the enhanced immunogenicity of HA-Fe-nanoparticles and the germinal center response is due to the abundance of CD4 T cells recruited after vaccination and that the Fe core typically contributes substantially to elicitation of the cognate CD4 T cells help needed for the robust germinal center response observed in the analyses of this vaccine platform.
留言 (0)