
記住我



Previously, we demonstrated that hUCs are reprogrammable into iPSCs, neural progenitor cells (NPCs) and cardiomyocytes by defined factors [13, 30, 31], indicating the high cell fate plasticity of hUCs. Hence, we speculated whether hUCs could be reprogrammed into other cell fate states, such as kidney lineage-associated progenitor cells, which are the developmental ancestors of differentiated hUCs. To test this hypothesis, we isolated and cultured hUCs in renal epithelial growth medium (REGM), and nearly 100% of hUCs expressed the tubular epithelial cell marker, PAX8. Furthermore, hUCs were reprogrammed in a series of defined medium. Through screening and optimization, we eventually developed a defined medium including the following chemicals: CHIR99021 (5 μM), EPZ5676 (1 μM) and Actinvin A (10 ng/mL), bFGF (10 ng/mL), and EGF (10 ng/mL) to convert hUCs into U-iIMPLCs within 9 days. Followed by step-wise differentiation under the defined medium, U-iIMPLCs were induced into nephron progenitor cells (U-iNPCs) at 14–16 days and then U-iKOs at 22–24 days (Fig. 1A, B). These intermediate identities were confirmed through gene expression analysis such as T, MIXL1, and TBX6 for primitive streak; WT1 and HOXD11 for nephrogenic intermediate mesoderm; SIX2, WT1, and PAX2 for nephron progenitors; Podocalyxin (PODXL) and WT1 for podocytes, and CDH6 and JAG1 for tubule epithelial cells (Fig. 1C, Additional file 1: Fig. S1A, F). Furthermore, they did not express markers for the pluripotent, definitive endoderm, or ectoderm (Additional file 1: Fig. S1B). We successfully generated organoids from 11 individuals, including 8 healthy volunteers and 3 patients with CKD from both males and females (Additional file 1: Fig. S1C). Patient-derived urine cells underwent a similar process of cellular morphological changes and gene expression profile changes to healthy volunteers (Additional file 1: Fig. S1D, E).
Fig. 1
Generation of 2D kidney organoids from hUCs. A Schematic illustration of 2D kidney organoids starting from hUCs (human urine cells), hUCs-induced intermediate mesoderm progenitor like cells (U-iIMPLCs), hUCs-induced nephron progenitor cells (U-iNPCs), hUCs-induced kidney organoid (U-iKOs) at the indicated time points. B Representative bright-field images during A at day0, day9, day16, and day24. Scale bars, 250 μm. C Gene expression analysis for markers of the primitive streak (T, MIXL1, and TBX6), nephrogenic intermediate mesoderm (WT1 and HOXD11), nephron progenitor (SIX2, WT1, and PAX2), podocytes (PODXL and WT1), tubule epithelial cells (CDH6 and JAG1) during differentiation. The relative expression of each transcript to GAPDH expression is presented as the mean ± SEM (n = 6). D Representative loci for the close-to-open (CO), open-to-close-to-open (OCO), and close-to-open-to-close, (COC) peaks at different stages at hUC (day0), U-iIMPLCs (stage1, day9), early U-iNPCs (stage2, day16) and late U-iNPCs (stage2, day19). E Dynamic CO, OC, and permanently open (PO) chromatin regions are clustered for hUC, U-iIMPLCs, U-iNPCs (E) and U-iNPCs (L). Representative genes are noted for each subgroup on the right side. F Heatmaps for the motif enrichment among the OC, CO and PO subgroup peaks for stage1, and stage2 as shown in E. *p < 1e-30
To further characterize the process of U-iKOs generation, we profiled the chromatin accessibility and transcriptome dynamics using ATAC-seq and RNA-seq respectively at each stage. At the chromatin level, ATAC-seq analysis revealed that genomic loci associated with MIXL1, TBX6, WT1, PAX8, LHX1, CALB1, and PODXL underwent closing or opening as shown in Fig. 1D. Notably, the PAX8 locus closed at the first stage, but then opened upon acquisition of kidney lineage fate (Fig. 1D), whereas the TBX6 locus opened at the first stage but then closed at stage 2 (Fig. 1D). Globally, loci underwent dynamic close-to-open (CO), or open-to-close (OC) remodeling during the entire process as shown in Fig. 1E. At each transition, we identified critical genes up- or down-regulated by time-course RNA-seq (Additional file 1: Fig. S2A), with corresponding biological pathways including mesoderm development, kidney development, and epithelial cell morphogenesis involved (Additional file 1: Fig. S2B–D). Interestingly, loci with CO or OC were enriched with specific transcription factors, such as those from the FORKHEAD, HMG, HOMEBOX, T-BOX, TEAD, and BZIP families (Fig. 1F), reflecting the specific roles of each factor during the two main transitions from U-iIMPLCs to U-iNPCs and then from U-iNPCs further to U-iKOs.
The surprisingly simple way to generate organoids directly further encouraged us to design methods to generate 3D organoids. As shown in Fig. 2A, we came up with a relatively straightforward procedure to generate 3D in 24 days. As shown in Fig. 1, we picked colonies from a 16-day culture and then either aggregated them directly for 1 day or dissociated them into single cells before aggregation for 5 days. These aggregated cells were then placed in the 3D air–liquid interface (ALI) culture, as illustrated in Fig. 2A. On day 19, cells from colonies grew into organoids, whereas those from single cells remained as spherical cysts (Fig. 2B). Yet, at day 24, both procedures generated organoids with specialized and complex structures (Fig. 2B, right panels). We further characterized these organoids by performing immunostaining for the specific markers as shown in Fig. 2C. We accurately identified kidney-specific structures and cells such as podocytes (PODXL, nephrin, and WT1), proximal tubules (LTL), medial tubules (JAG1), distal tubules (PAX2, PNA and CDH1), and basement membrane (LAM). These results suggested that high-quality 3D organoids can be generated using aggregation and ALI culture conditions. Together, through partial reprogramming and staged differentiation under defined medium, we obtained personalized kidney organoids within a month. To obtain kidney organoids from PSCs in a personalized way, somatic cell reprogramming is required to obtain iPSCs and then proceed to the differentiation process, which typically takes 2–3 months following literature (Fig. 1G) [6, 32].
Fig. 2
Establishment of 3D U-iKOs. A Schematic illustration of 3D U-iKOs. Colonies at day14 in Fig. 1 were picked and then aggregated as pellets or single cells in suspension and air–liquid interface (ALI) culture to form 3D U-iKOs at indicated time points in B. B Representative bright-field images of 3D U-iKOs as in A. Scale bars, 250 μm. C Immunostaining analysis of nephron segment-specific markers for podocytes (PODXL, nephrin, and WT1), proximal tubule (LTL), medial tubule (JAG1), distal tubule (PAX2, PNA and CDH1), and basement membrane (LAM). Scale bars, 50 μm
Long-term expanding U-iIMPLCs for U-iKOs generationDuring daily cell culture, certain hUCs from some individuals or batches proliferated slowly or poorly, which lengthened the time required to collect sufficient hUCs for reprogramming. Therefore, we examined whether any progenitor state during the U-iKOs generation could expand long-term in vitro, which is of vital importance for optimization, functional validation, and future application. By optimizing culture conditions, we succeeded in developing a defined medium including CHIR99021 (5 uM), EPZ5676 (1 μM), and hLIF (10 ng/mL) to support long-term growth and expansion of U-iIMPLCs, but not for U-iNPCs or U-iKOs (data not shown). Under the defined medium, U-iIMPLCs grew in tight clusters and expanded in vitro for more than 20 passages within 80 days (Fig. 3A, B). RNA-seq indicated that the stable transcription profiles of U-iIMPLCs were distinct from those of hUCs and hPSCs (Fig. 3C). In detail, U-iIMPLCs expressed intermediate mesoderm specific markers OSR1 and HOXD11 but not the paraxial mesoderm markers MSGN and MESP1, lateral mesoderm markers FOXF1 and NKX2.5, pluripotent genes NANOG, ectodermal genes SOX1 and PAX6, and endodermal genes SOX17 and FOXA2 (Fig. 3D). Consistently, qRT-PCR analysis showed that U-iIMPLCs from three different cell lines at passages 2, 5, 10,15, 20, and 25 similarly expressed T, MIXL1, OSR1, and HOXD11 similarly (Fig. 3E). To assess the potential of U-iIMPLCs for U-iKOs generation, we chose U-iIMPLCs from the same cell line at passage 5, 10, 15 and 6 different cell lines at passage 10 and seeded at 10‒12.5 × 104 cells/cm2 for U-iKOs induction (Fig. 3F, H). Following a two-stage process, U-iIMPLCs efficiently differentiated into kidney lineages and formed more than 600 2D U-iKOs per well of 24-well cell culture plate (Fig. 3G–I). Taken together, we concluded that U-iIMPLCs can self-renew in vitro under defined medium and possessed the potential to generate U-iKOs efficiently.
Fig. 3
Efficient maintained U-iIMPLCs retain nephrogenic potential. A Cell morphology of U-iIMPLCs subcultured at different passages. The same strain of U-iIMPLCs 1# with different passages P0, P1, P5, P9, P17, and P21 are presented. Scale bars, 250 μm. B Growth curve of U-iIMPLCs for different strains of U-iIMPLCs 2#, 6#, and 8#. C Correlation analysis showing gene expression profiles of hUCs, hPSCs, and U-iIMPLCs 1# maintained at different passages. D Gene expression analysis for hUCs, intermediate mesoderm, paraxial mesoderm, Lateral plate mesoderm, pluripotent, ectoderm, and endoderm markers of the cells shown in C. E Gene expression analysis for intermediate mesoderm progenitor markers for different strains with different generations. The relative expression of each transcript to GAPDH expression is presented as the mean ± SEM. F Representative stereo microscope images showing 2D kidney organoids differentiated from the same strain of 8# U-iIMPLCs at different passages in a single well of a 24-well plate. Scale bars, 500 μm. G The number of 2D U-iKOs in F. H Representative stereo microscope images showing 2D kidney organoids differentiated from different strains at passage 10. Scale bars, 500 μm. I The number of 2D kidney organoids is shown in H
Characterizing U-iKOs by single cell RNA sequencing (scRNA-seq)Cell type diversities in organoids were further described using scRNA-seq. These results showed that the 10,125 single cells from 2D U-iKOs could be classified into 13 distinct clusters (Fig. 4A), while the 10,739 single cells from 3D U-iKOs into 11 distinct clusters (Fig. 4B) with high quality (Additional file 1: Fig. S3A). Overall, four major cell types were identified, including podocytes, tubules, mesenchyme, and neural cells, which were consistent with the immunostaining results described above. These main types were further classified into 13 and 11 subtypes in the 2D and 3D organoids respectively with specific markers identified (Fig. 4C, D). The heatmap shows similar cell types in 2D and 3D organoids with a high correlation (Additional file 1: Fig S3B). For the 2D culture of U-iKOs, the predominant cell types were mesenchyme related with five distinct subtypes, specifically labeled with PCLAF/UBE2C (subtype1), SFRP2/COL2A1 (subtype 2), PLP1/NPR3 (subtype 4) and H2BC9 and CSKMT (subtype 5) (Fig. 4C). Interestingly, subtype 3 appeared to share markers with subtypes 1 and 2, suggesting that these cells may be intermediates (Fig. 4C). On the other hand, 3D U-iKOs had ~ 57% tubular epithelial cells, suggesting that the 3D condition favors the tubular structure. Reflecting being more mature, 3D U-iKOs featured a collecting duct-like that was absent from the 2D ones (Fig. 4B, D). Furthermore, the various cell types in 3D U-iKOs had more complex markers than their counterparts in 2D, again reflecting a more mature state (Fig. 4D). The difference between 2 and 3D became apparent when cell type diversities were compared directly, as shown in Fig. 4E, F, as 3D organoids were enriched with the loop of Henle, proximal tubule, distal tubule, podocyte, and tubule progenitors, and collecting tubule compared to 2D organoids with podocyte and distal tubule. It was of interest to note that, even with the same origin of cells and the same culture media, culture dimensions, i.e., 2D vs. 3D, they conferred drastically different cell fates (Fig. 4G–I). For instance, 2D generated more distal tubule cells and tubule GDF15 + cells (Fig. 4G–I). Collecting tubule-like cells were unique in only 3D conditions, reflecting the spatial requirement for specific cell types such as collecting tubule lineage (Fig. 3G–I). Hence, our findings demonstrated that 3D is superior to 2D in generating more mature kidney organoids.
Fig. 4
ScRNA-seq profiling of cell types in U-iKOs. A UMAP projection of 10,125 single cells from 2D U-iKOs into 13 distinct clusters. Four main cell types are circled on the diagram with different colors. B UMAP projection of 10,739 single cells from 3D U-iKOs into 11 distinct clusters. Four main cell types are identified in A. C Heatmap of well-known markers and signature genes in A. D Heatmap of well-known markers and signature genes in B. E Barplot of the tubule, podocyte, neuron, and mesenchyme between 3 and 2D U-iKOs. F UMAP projection of tubule and podocyte from 3 and 2D U-iKOs into 9 clusters. G Signature genes in distal tubule1, distal tubule2, tubule_GDF15 + , and collecting duct (CD). H Barplot of 3D and 2D cells in each cluster in F. I Heatmap of differentially expressed genes between 4 clusters in F, selected signature genes, and GO term with p values in the right
The podocytes were further classified into four different groups, including NPHS2, SUSD3, MKI67, and CLDN6 respectively (Additional file 1: Fig. S3C). These four groups of cells appeared to capture different stages of podocyte development. The NPHS2 + cells are the mature type expressing PODXL, PTPRO, CLIC5, and other genes critical to glomerular structure and function (Additional file 1: Fig. S3C, D, blue). Marked by CLDN6, a tightly clustered group of podocytes (Additional file 1: Fig. S3C, red), expressed genes critical for kidney morphogenesis, nephron development, and epithelium development (Additional file 1: Fig. S3D, bottom). The MKI67 + cells actively proliferated and expressed genes involved in DNA replication, nuclear division, and chromosomal segregation (Additional file 1: Fig. S3C, green). The SUSD3 + cells expressed genes actively involved in metabolic processes and hypoxia response (Additional file 1: Fig. S3D).
The proximal tubule cluster of cells were subdivided into 5 groups, marked by IRX1, CUBN, WT1, SLC2A1, and MKI67 respectively (Additional file 1: Fig. S3E). Remarkably, CUBN + proximal tubule cells reflected the stage of differentiation and also expressed genes related to reabsorption function, such as LRP2, APOM, and DAB. Again, the MKI67 marked the proliferation group that also expressed genes implicated in DNA replication and the cell cycle (Additional file 1: Fig. S3F). The other three groups of cells reflected various stages of development, as exemplified by the mature cell types in the proximal tubule /loop of Henle (Additional file 1: Fig. S3E, F). The proximal tubule is a major kidney function structure and the recapitulation of these cell types further demonstrated the potential of our approach. The loop of Henle and the distal tubule was well represented well in the organoids, including diverse cell types from GATA3 + cells to SLC12A1 + cells, again reflecting the fates of cells undergoing differentiation and maturation (Additional file 1: Fig. S3G, H). Mature cell types marked by SLC12A1, CLDN10, ATP1A2, and other genes depict the function of the loop of Henle with ion homeostasis and regulation of pH. The GATA3 + showed the expression of collecting duct-specific genes SLC14A2, TBX3, and SLC12A2, which produced progenies with branching morphogenesis of an epithelial tube and ureteric bud morphogenesis. (Additional file 1: Fig. S3G, H).
Together, the identification of individual cell types within the organoids suggested that the 3D condition recapitulated various aspects of in vivo development and generated cell types that are available under physiological condition.
U-iKOs are rich in mitochondriaDue to the unique way of U-iKOs generation, we hypothesized that U-iKOs might possess some different features from the developed P-iKOs recently. In addition, shortage of the research on the similarity between organoids and human fetal kidney also inspired us to make comparisons. To address this, we collected similar scRNA-seq datasets generated from P-iKOs and published human fetal kidney at 16 weeks, and then made comparisons, showing that U-iKOs and P-iKOs shared most tubule epithelial cells and mesenchyme cells (Fig. 5A). In detail, U-iKOs contained collecting duct-like cells which were not generated in P-iKOs, and a higher percentage of the loop of Henle/distal tubule (Fig. 5B). In contrast, we verified that P-iKOs and fetal kidney contained the endothelial cells subpopulation that was absent in U-iKOs. Off-target cells, such as muscle cells and melanocytes, were generated in P-iKOs but not in U-iKOs (Fig. 5B), suggesting cell fate commitment of U-iIMPLCs for U-iKOs induction. The differences in cellular composition between U-iKOs, P-iKOs and fetal kidney inspired us to make an in-depth analysis in molecular level. GO term analysis revealed that U-iKOs were specifically enriched in mitochondria ATP synthesis and metabolic process, proton transmembrane transport, and kidney development (Fig. 5C), whereas differentially expressed genes in P-iKOs were associated with Golgi vesicle transport, RNA splicing, and cytoplasmic translation (Additional file 1: Fig. S4A–D). Compared to organoids, human fetal kidney was enriched in response to unfolded protein and response to temperature stimulus with higher expression level of HSPA1A, HBG2 and GPC3, consistent with those previously described [28]. Consistently, we found that U-iKOs expressed a higher level of kidney function associated genes, such as LRP2 (megalin; macromolecules and numerous ligands reuptake), CUBN (endocytic receptor), AGT (pre-angiotensinogen), PRCP (a serine exopeptidase that acts on angiotensin II), PKM (glycolysis), and LDHA (oxidoreductase) (Fig. 5D). The results revealed that membrane transporters and mitochondria metabolism-related genes were specifically expressed in U-iKOs but not in P-iKOs, such as SLC12A1 (sodium, potassium, and chloride ion cotransporter), SLC5A3 (sodium transport), SLC25A1 (tricarboxylate transporter), SLC25A3 (phosphate carrier protein, mitochondrial), and ATP1A2 (sodium/potassium-transporting ATPase) (Fig. 5D), suggesting that cells in U-iKOs may contain abundant mitochondria. The expression of these genes was also validated by qRT-PCR detection (Fig. 5E). Nephron function maker, such as SLC12A1, LRP2 were validated in 3D U-iKOs (Additional file 1: Fig. S4E). To further confirm these results, we examined cellular organelles in hUCs and U-iKOs using TEM and showed that both proximal and distal tubule epithelial cells consisted of a number of mitochondria in the cytoplasm (Fig. 5F). Taken together, these results demonstrated that U-iKOs are rich in mitochondria, validating our hypothesis that mitochondria-rich kidney organoids may be easily induced from mitochondria-rich human urine cells through partial reprogramming.
Fig. 5
U-iKOs are rich in mitochondria. A The UMAP shows the integration of 3D U-iKOs, M-iPSC, T-iPSC, C-iKOs, and human fetal kidney, with cell types on the right. M-iKOs, iPSC-induced organoids with protocol by Morizane; T-iKOs, iPSC-induced organoids with protocol by Takasato; C-iKOs, iPSC-induced organoids from Combes’s study; Fetal_kidney, human fetal week16 kidney from Lindström’s study. B Comparison of the proportions of cell types in 3D U-iKOs, M-iKOs, T-iKOs, C-iKOs and human fetal kidney. C GO analysis of differentially expressed genes in 3D U-iKOs. D Dotplot shows differentially expressed genes of 3D U-iKOs related to nephron function (top panel), membrane transporters (middle panel), and mitochondia metabolism (bottom panel) compared with with other data. E Gene expression analysis for differentially expressed genes in U-iKOs in D of the mature tubule (CUBN, LRP2, and SLC12A1), AGT (angiotensinogen), and mitochondria metabolism (ATP1A2, ATP5G1, MUC1, and MRPL48). The relative expression of each transcript to GAPDH expression is presented as the mean ± SEM (n = 5). *P < 0.05 vs. the U-iIMPLCs; **P < 0.01 vs. the U-iIMPLCs; ***P < 0.001 vs. the U-iIMPLCs; ****P < 0.0001 vs. the U-iIMPLCs. (F) TEM of hUCs (upper panel) and 3D U-iKOs (botttom panel). Abundant mitochondria indicated with red asterisks. Scale bars, 1 μm
Structure and function of developed 3D U-iKOsScRNA-seq provided evidence that the 3D organoids possessed the cellular diversity to perform kidney functions. To further characterize the structure and anatomy of the organoids, we observed the uptake of dextran under 100 kDa and the exclusion of those above 2000 kDa (Fig. 6A). The results showed that these organoids function similarly to the kidneys. Additionally, we showed that the ultrastructure of the organoids resembled those of the kidneys. Within U-iKOs, the podocytes had typical macronuclei and primary and secondary foot processes. The putative proximal tubule epithelium formed lumen-like structures with dense apical microvilli at the brush border and abundant mitochondria in the cytoplasm. The putative distal tubule epithelium showed sparse and short microvilli and tight junctions (Fig. 6B).
Fig. 6
Structure and function of 3D U-iKOs. A Immunofluorescence analysis of fluorescence-labeled dextran uptake assay in 3D U-iKOs for 10 kDa, 70 kDa, and 2000 kDa of dextran. Scale bars, 50 μm. B TEM of 3D U-iKOs (day 24). Podocyte (p) with characteristic large nuclei and primary processes (pp) and secondary processes (sp) in the upper panel. A putative proximal tubule cells with enriched mitochondria (mit) and a lumen filled with closely packed microvilli and the brush border (bb) in the middle panel. A putative distal tubule with relatively sparse short microvilli (m) and tight junctions (tj) in the bottom panel. Scale bars, 2 μm. C H and E staining of implanted kidney organoids in the CAM for 6 days. Magnified views of glomerular (G) and tubular (T) cells are shown. Scale bars, 100 μm. D Confocal microscopy images of implanted organoids in C stained for parietal epithelial cells maker (PAX8), bowman’s capsule basement membrane (LAM), and tubular markers (LTL, CDH1, and PAX8). Scale bars, 50 μm
We then implanted the organoids into the CAM and found that they further matured structurally within 7 days (Fig. 6C, Additional file 1: Fig. S5A–C), with glomerular and tubular cells as marked. Finally, we performed confocal microscopy on grafts (Fig. 6D) and showed that glomerular markers PODXL, nephrin, WT1, PAX8, and LAM and tubular markers LTL, CDH1, and PAX8 were present in the developed organoids. We also noticed that PAX8 + parietal epithelial cells were present on the wall of Bowman's capsule, indicating that glomeruli acquired a much more mature architecture in the CAM transplantation. Together, these results suggested that a rather comprehensive recapitulation of kidney function could be established in vitro through our reprogramming approaches.
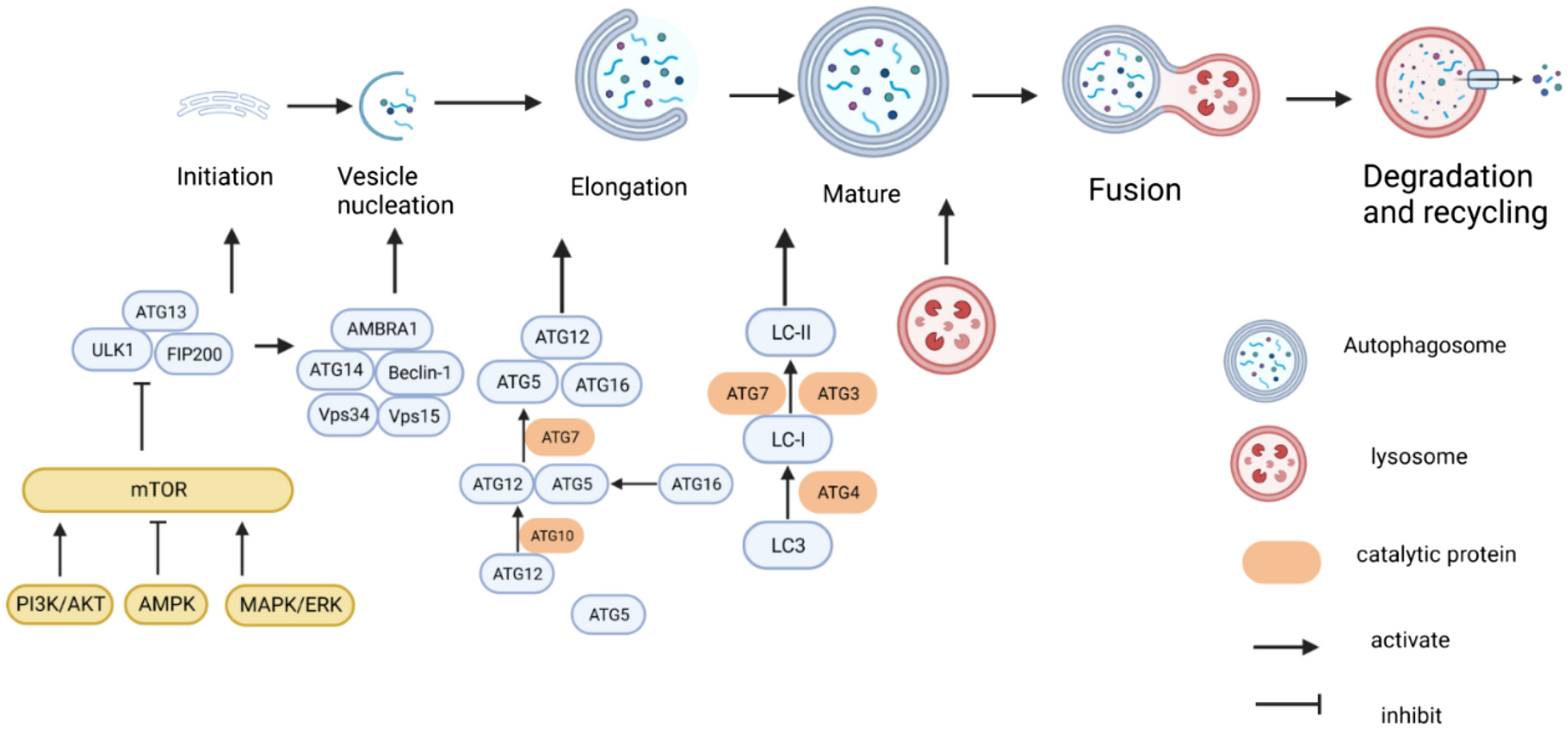
Generation of kidney disease models with 3D U-iKOsThe structure and function of U-iKOs further inspired us to evaluate whether these organoids be used to model kidney diseases. To this end, we treated the U-iKOs with cisplatin for 24 h, which is a nephrotoxic drug used in the chemotherapy of malignant tumors [33]. Interestingly, cisplatin treatment resulted in the up-regulation of kidney injury molecule-1 (KIM-1), a major marker for acute kidney injury (Fig. 7A). Hematoxylin–eosin (HE) staining showed that tubular epithelial structures were nearly destroyed following cisplatin treatment (Fig. 7B). Meanwhile, we collected U-iKOs samples with or without cisplatin treatment for RNA-seq. GO analysis further indicated that cisplatin treatment triggered upregulation of genes that are strongly enriched in cell cycle DNA replication, signal transduction by p53 class mediator, apoptotic signaling pathway, and mitochondrial depolarization while downregulated genes were closely related to glomerular epithelial cells differentiation and mitochondria respiratory chain assembly (Fig. 7C, D). In addition to chemotherapeutic drugs, antibiotics such as gentamicin, are another kind of nephrotoxic drugs [34]. Thus, we treated the U-iKOs with gentamicin for 24 h and observed that U-iKOs displayed similar cellular morphological changes as cisplatin treatment and upregulated KIM-1 expression (Fig. 7E, F). RNA-seq data showed that gentamicin treatment increased the expression of genes involved in unfolded protein, apoptosis, negative regulation of cell growth and mitochondria autophagy, and the downregulated genes related to nephron tubule morphogenesis and ion homeostasis (Fig. 7G, H). On the other hand, forskolin or FSK, a chemical agonist to activate intracellular cAMP levels, is often used for cystogenesis induction in P-iKOs from patients with polycystic kidney disease (PKD) or non-PKD iPSCs [8, 35, 36]. Consistently, we observed that U-iKOs formed enlarged cyst structures after FSK treatment for 4 days, as validated by HE staining and immunostaining with tubule epithelial cell markers (Additional file 1: Fig. S6A–C). Interestingly, we found that FSK treatment enhanced renin, AGT, and PRCP expression which was confirmed in P-iKOs recently [37] (Additional file 1: Fig. S6D, E). Compared with untreated U-iKOs, FSK treatment led to unregulated genes enrichment in the regulation of cell morphogenesis and organelle fission while downregulated genes affected kidney epithelium development and pattern specification process (Additional file 1: Fig. S6F, G). Finally, although all these three models affected mitochondria homeostasis, we assessed and compared the expression of genes involved in mitochondria dynamics. Cisplatin, gentamicin, and FSK treatment all affected mitochondrial function associated genes such as BID, ATG13, BNIP3, AMBRA1, SURF1, and COX5A (Fig. 7I, left), which were previously demonstrated to regulate mitochondria-related kidney pathologies in vivo [3, 38,39,40,41,42]. The expression of BID, AMBRA1, and ATG13 was also validated by qRT-PCR (Fig. 7J–L). On the other hand, mitochondria homeostasis-associated genes were specifically expressed in these three models respectively, suggesting that cisplatin, gentamicin, and FSK treatment resulted in kidney injury in a pathology-specific manner (Fig. 7I, right panel). Then, we also validated differentially altered genes expression in each disease model by qPCR (Additional file 1: Fig. S6H). Taken together, these results demonstrated that U-iKOs can be used as an in vitro model to mimic kidney injury conditions related to mitochondria homeostasis in a personalized manner.
Fig. 7
3D U-iKOs disease models. A Immunofluorescence analysis of 3D U-iKOs treated with cisplatin (5 μM) for 24 h. Scale bars, 50 μm. B H and E staining of U-iKOs treated with cisplatin in A. Scale bars, 100 μm. C Scatterplot showing the up-regulated genes (red) and down-regulated genes (blue) between control and cisplatin treatment groups. D GO analysis of C. E Representative bright-field images of control and gentamicin treatment cells. Scale bars, 250 μm. F Immunofluorescence analysis of 3D U-iKOs treated with control and gentamicin (5 mg/mL) for 24 h. G Scatterplot showing the up-regulated genes (red) and down-regulated genes (blue) between control and gentamicin treatment groups. H GO analysis of G. I Heatmap shows common differentially expressed genes of FSK, cisplatin, and gentamicin treatment groups compared to control and their respective differentially expressed genes. J–L Gene expression analysis for differentially expressed genes after cisplatin, gentamicin, and FSK treatment in I. The relative expression of each transcript to GAPDH expression is presented as the mean ± SEM (n = 4). *P < 0.05 vs. the control group; **P < 0.01 vs. the control group; ****P < 0.0001 vs. the control group
留言 (0)