
記住我










Antibiotic resistance is recognised as a global threat to human health by national healthcare agencies, governments and medical societies, as well as the World Health Organization. Increasing resistance to available antimicrobial agents is of concern for bacterial, fungal, viral and parasitic pathogens. One of the greatest concerns is the continuing escalation of antimicrobial resistance among Gram-negative bacteria resulting in the endemic presence of multidrug-resistant (MDR) and extremely drug-resistant (XDR) pathogens. This concern is heightened by the identification of such MDR/XDR Gram-negative bacteria in water and food sources, as colonisers of the intestine and other locations in both hospitalised patients and individuals in the community, and as agents of all types of infections. Pneumonia and other types of respiratory infections are among the most common infections caused by MDR/XDR Gram-negative bacteria and are associated with high rates of mortality. Future concerns are already heightened due to emergence of resistance to all existing antimicrobial agents developed in the past decade to treat MDR/XDR Gram-negative bacteria and a scarcity of novel agents in the developmental pipeline. This clinical scenario increases the likelihood of a future pandemic caused by MDR/XDR Gram-negative bacteria.
AbstractAntimicrobial resistance continues to rise among Gram-negative bacteria, leading to greater morbidity, mortality, lengths of stay and costs. The level of resistance is approaching pandemic proportions, requiring an urgent call to address this problem. https://bit.ly/3NTnDqK
IntroductionRespiratory infections are among the most common indications for hospitalisation to include admission to an intensive care unit (ICU) [1]. The novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) respiratory virus pandemic is estimated to have accounted for >7 million deaths worldwide [2] and resulted in significant economic distress with the global economy contracting by 3.5% in 2020 [3]. Another “silent” pandemic that has been going on for more than three decades is the increasing prevalence of bacterial infections attributed to multidrug-resistant (MDR) and extremely drug-resistant (XDR) Gram-negative bacteria [4]. The United Nations Interagency Coordination Group on Antimicrobial Resistance Report considers bacterial antimicrobial resistance to be a major threat to human health, and a recent Wellcome Trust report suggests that nearly 300 million individuals will die over the next several decades as a direct result of antimicrobial resistance [5, 6]. Similarly, in the United States, antibiotic-resistant pathogens cause >2 million infections and 23 000 deaths per year, as reported by the United States Centers for Disease Control and Prevention [7]. Despite the introduction of novel antibiotics, the continued escalation of resistance among Gram-negative bacteria suggests that the problem of antibiotic resistance is likely to intensify in the future, leading to antimicrobial inadequacies and a potential full-blown pandemic [8].
The Extended Study on Prevalence of Infection in Intensive Care III (EPIC III) found that among the 15 165 qualifying patients, 8135 (54%) had at least one suspected or proven infection on the study day, with most being respiratory infections [1]. Moreover, antibiotic-resistant pathogens including vancomycin-resistant Enterococcus, Klebsiella species resistant to β-lactam antibiotics or carbapenem-resistant Acinetobacter species were associated with the highest risk of in-hospital death [1]. Given the importance of escalating antibiotic resistance among Gram-negative bacteria as a cause of mortality, morbidity and economic hardship, we brought together in this issue of the European Respiratory Review a multidisciplinary group of authors to discuss this possible next pandemic. The goal of this review is to provide a concise appraisal of the problem of escalating antibiotic resistance among Gram-negative bacteria as an important class of agents for respiratory infections. It is also important to understand that pandemics are not mutually exclusive, as evident by the SARS-CoV-2 pandemic which contributed to outbreaks of MDR/XDR Gram-negative infections including pneumonia [9, 10]. Therefore, as a medical community we must maintain a state of preparedness in order to deal with future pandemics, to which antibiotic resistant Gram-negative bacteria will likely contribute, if not cause outright.
Antibiotic resistant Gram-negative pulmonary infections: key organisms and their epidemiologyAccording to recent estimates, antibiotic-resistant infections were associated with nearly 5 million deaths globally in 2019 alone [11]. The most common site of infection among these was the lower respiratory tract, which accounted for 1.5 million of the deaths and >75 000 000 disability-adjusted life-years [11]. Although not delineated according to infectious syndrome by the authors, the Gram-negative pathogens associated with the most deaths included Escherichia coli, Klebsiella pneumoniae, Acinetobacter baumannii and Pseudomonas aeruginosa, each associated with >250 000 deaths in 2019, although incidence of the pathogens and death rates varied substantially by region [11].
Among individual pathogen–antimicrobial combinations, in 2019, >50 000 deaths each were directly attributable to third-generation cephalosporin-resistant K. pneumoniae and E. coli, carbapenem-resistant A. baumannii and K. pneumoniae, and fluoroquinolone-resistant E. coli (figure 1) [11]. The deaths associated with and attributable to antimicrobial resistance as estimated by the Antimicrobial Resistance Collaborators [11] were derived from a variety of data sources from across the world. However, these estimates were derived from 2019 data among patients who became ill before the onset of the SARS-CoV-2 pandemic. Studies showed that even before the SARS-CoV-2 pandemic, cases of pneumonia due to antimicrobial-resistant Gram-negative bacteria were on the rise in many regions throughout the world [12]. 3 years into the pandemic, we know that patients hospitalised with SARS-CoV-2 receive antimicrobials at very high rates (≥75% in some instances), a situation which will only exacerbate the global antimicrobial resistance crisis that was burgeoning pre-pandemic [13]. In a vicious positive-feedback cycle, patients with SARS-CoV-2 are known to have higher rates of ventilator-associated pneumonia (VAP) [14], a situation which will lead to increased levels of broad-spectrum antimicrobial use, and subsequent development of antimicrobial resistance. In addition, despite the high numbers of antimicrobial-resistant lower respiratory infections found in the aforementioned study [11], this may be an underestimate, as causative pathogens in pneumonia are identified in only ∼30–60% of patients [15–18].
 FIGURE 1
FIGURE 1 Global deaths in 2019 directly attributable to Gram-negative antimicrobial resistance by pathogen–antimicrobial. Data from [11]. 3GCR: third-generation cephalosporin resistance; AGR: aminoglycoside resistance; CR: carbapenem resistance; FQR: fluoroquinolone resistance.
In the study by the Antimicrobial Resistance Collaborators, methicillin-resistant Staphylococcus aureus (MRSA) and drug-resistant E. coli were the two most common organisms causing mortality [11]. For patients with pneumonia, there is good evidence that MRSA nare swabs have a high negative predictive value and can be used in antimicrobial stewardship efforts to stop unnecessary anti-MRSA antimicrobial use [19]. Unfortunately, there is no easy analogue/stand-in to assist in ruling out drug-resistant Gram-negative pneumonia. Some efforts have attempted to understand the relationship between intestinal colonisation with antimicrobial-resistant Gram-negatives, including those producing extended-spectrum β-lactamases and/or carbapenemases, and the development of future infection, but operationalising such practices has proven challenging and may lead to overtreatment with novel antimicrobials, which must be preserved to prevent development of new resistance [20–23]. Even before the SARS-CoV-2 pandemic, the match between prescribed antimicrobials and recovered pathogens was poor, with MRSA and Pseudomonas being treated far more often than they were detected [24].
At the individual level, patients with antimicrobial-resistant Gram-negative pneumonia are more likely to have been treated previously with antibiotics, be bed-bound, have longer lengths of hospitalisation, have a previous microbiology history of resistant organisms, and to have certain acute and chronic medical comorbidities, with variation by hospital case-mix and region [16, 24–29]. Understanding the risk factors for infection with antibiotic-resistant organisms is crucial for effective empiric treatment decision making. One helpful tool is the “PES” score aimed at identifying antibiotic-resistant Pseudomonas, Enterobacterales and MRSA in patients with pneumonia [30] (table 1).
TABLE 1The “PES” score to assess the risk of pneumonia due to Pseudomonas aeruginosa, Enterobacteriaceae with extended-spectrum β-lactamases and methicillin-resistant Staphylococcus aureus pathogens
With continued antimicrobial selection pressure, additional mechanisms of antimicrobial resistance are likely to evolve, as has happened with each new antimicrobial that has been developed and used clinically. Identifying risk factors for the development of antimicrobial resistance and the prospective use of algorithms to predict which hospitalised patients will go on to develop antimicrobial resistance may be helpful in certain instances and/or localities, but rapid detection of pathogens and their antimicrobial resistance profiles is likely to be more sensitive for resistance detection [31]. The implementation of rapid molecular diagnostics, especially for pneumonia and bloodstream infections, may be a key step forward in minimising antimicrobial overuse.
Overview of main resistance mechanismsResistance mechanisms contributing to the development of multidrug-resistant Gram-negative organisms include enzymatic inactivation, efflux pumps, porin mutations and target site modifications [32] (figure 2). Mutations leading to resistance may be intrinsic to the organism or acquired via plasmid-mediated transmission [33]. Each plasmid-mediated conjugation may contain multiple resistance determinants, thus conferring a multidrug-resistant phenotype [32]. Table 2 summarises the relevant resistance mechanisms discussed in this section, in addition to organisms commonly harbouring such mechanisms, the phenotypic result and potential therapeutic approaches.
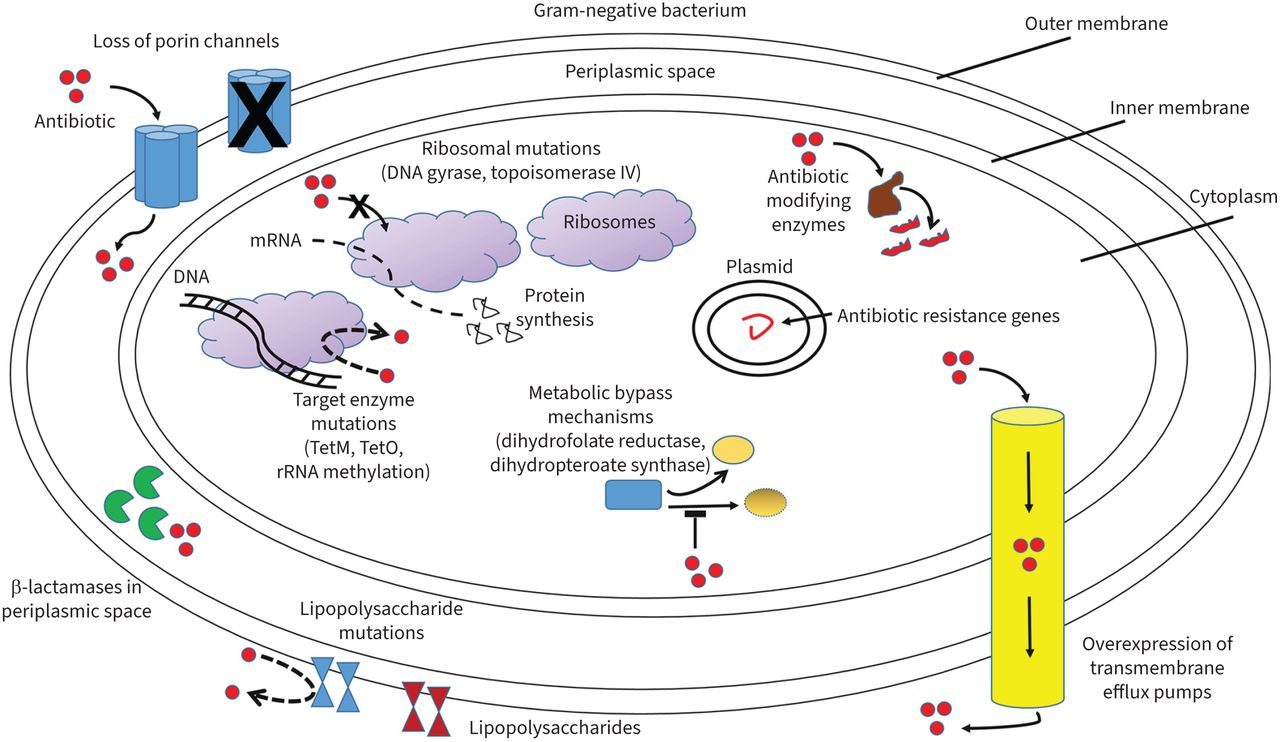 FIGURE 2
FIGURE 2 Mechanisms of antibiotic resistance in Gram-negative bacteria. Shown are loss of porin channels which reduce antibiotic movement across the bacterial membrane; β-lactamases in the periplasmic space inactivating β-lactams; increased transmembrane efflux pump expression expelling antibiotics from within the bacteria; antibiotic-modifying enzymes altering antibiotics so they cannot interact with end targets; antibiotic target and ribosomal mutations interfering with antibiotic actions; metabolic bypass mechanisms allowing alternative enzyme pathways bypassing antibiotic inhibitory effects; and lipopolysaccharide mutations limiting specific antibiotics such as polymyxins from disrupting the cell membrane.
TABLE 2Antibiotic mechanisms of resistance of Gram-negative organisms
β-Lactamasesβ-Lactamases are categorised molecularly with the Ambler classification based on the enzymatic active site structure and amino acid sequences, dividing them into classes A (narrow and extended-spectrum β-lactamases), C (AmpC cephalosporinases) and D (oxacillinases), which all utilise serine for hydrolysis, and B (metallo-β-lactamases), which utilise zinc ions for hydrolysis [34]. Functionally, β-lactamases are categorised into Bush–Jacoby groups, based upon their hydrolysis and inhibition profiles [34–36]. Phenotypically, β-lactamases include extended-spectrum β-lactamases (ESBLs), AmpC cephalosporinases and carbapenemsases [36]. A summary of β-lactamase classification is displayed in figure 3.
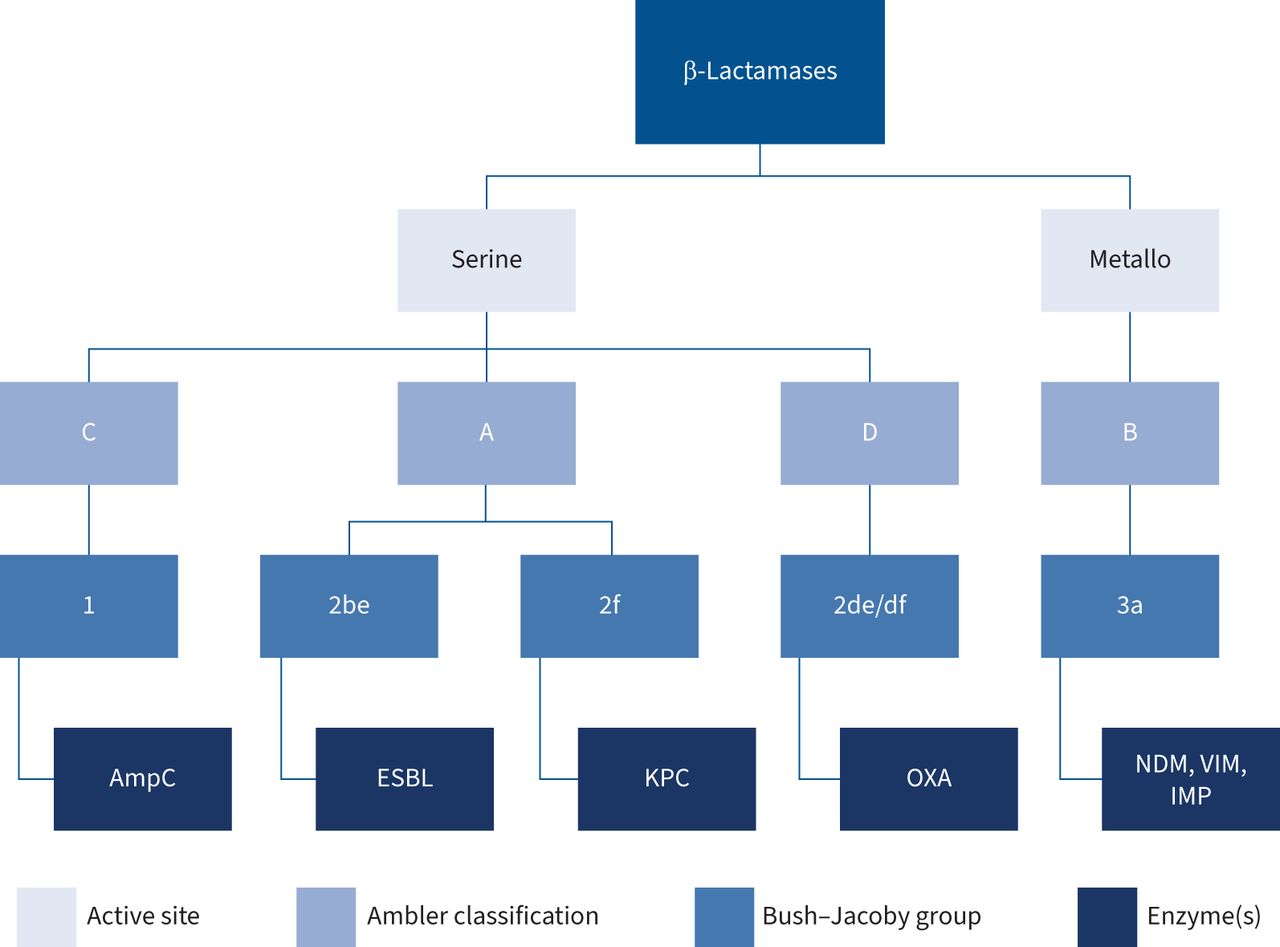 FIGURE 3
FIGURE 3 Molecular, functional and phenotypic classification of β-lactamase enzymes. ESBL: extended-spectrum β-lactamase; KPC: Klebsiella pneumoniae carbapenemases; OXA: OXA-β-lactamases; NDM: New Delhi metallo-β-lactamase; VIM: Verona integron-encoded metallo-β-lactamase; IMP: imipenem's metallo-β-lactamase.
ESBLsESBLs confer resistance to penicillins, cephalosporins including extended-spectrum (third and fourth generations) and monobactams, and are categorised as Ambler class A and Bush–Jacoby group 2 [35, 37]. This group of β-lactamases is genetically diverse, with the most common mutations including TEM, SHV and CTX-M isolated in Enterobacterales species [32, 38]. CTX-M-15 is the most common variant worldwide, but there are nearly 500 recognised variants of the various mutations [35]. Contemporary ESBLs are reliably inhibited by newer β-lactamase inhibitors, including avibactam, relebactam and vaborbactam; however, older-generation β-lactamase inhibitors including clavulanate, sulbactam and tazobactam may succumb to the inoculum effect or may be rendered ineffective against isolates harbouring inhibitor-resistant β-lactamases [35, 39, 40]. OXA-type ESBLs (Ambler class D) are less common, but include derivations of OXA-10 and OXA-2, commonly detected in isolates of P. aeruginosa [35].
AmpCAmpC β-lactamases are classified as Ambler class C and Bush–Jacoby group 1 and are often isolated in Enterobacterales species [32, 41]. These enzymes result from either the inducible or stable derepression of ampC regulatory protein production or from transmission of plasmid-mediated ampC genes [42, 43]. Susceptibility testing of an organism with stable derepressed AmpC production will demonstrate resistance to third-generation cephalosporins and cephamycins, while inducible AmpC producing organisms will initially test susceptible to third-generation cephalosporins. These inducible organisms become resistant upon exposure to certain β-lactams, which causes disabling of the negative regulator protein, AmpR, resulting in increasing production of AmpC [42]. Potent inducers of AmpC production include the aminopenicillins, first-generation cephalosporins, and cephamycins, while weak inducers include piperacillin-tazobactam, aztreonam, and third-generation cephalosporins, all of which may be hydrolysed in varying levels of AmpC production. Cefepime and carbapenems are inducers of AmpC production, but withstand its hydrolysis [42].
CarbapenemasesCarbapenemases include β-lactamases from Ambler class A (K. pneumoniae carbapenemases (KPC)), class B (New Delhi metallo-β-lactamases (NDM), Verona integron-encoded metallo-β-lactamases (VIM) and imipenem's metallo-β-lactamases (IMP)) and class D (OXA-48 variants) [36]. Carbapenemase enzymes are isolated in Enterobacterales, Pseudomonas and Acinetobacter species [44].
KPCs are the most clinically relevant of the class A carbapenemases and have been identified across the globe [44–46]. Despite their nomenclature, these genes are found on transferable plasmids and are isolated in other Enterobacterales species, in addition to K. pneumoniae [47].
The three subclasses of metallo-β-lactamases (MBLs) (NDM, VIM and IMP) render resistance to carbapenems via hydrolysis utilising zinc, and they are not inhibited by available β-lactamase inhibitors [44, 48].
OXA-β-lactamases, specifically OXA-48 variants, have been isolated in Enterobacterales species. Because of their heterogeneity, the global prevalence of OXA-48 β-lactamases may be underestimated [44].
In addition to carbapenems, most carbapenemases also hydrolyse penicillins and cephalosporins, while only KPCs hydrolyse monobactams [45, 46]. Similar to ESBLs, KPCs are generally inhibited by traditional β-lactamase inhibitors, while OXA carbapenemases are not [45, 46]. KPCs are additionally susceptible to all of the newer-generation β-lactamase inhibitors. OXA carbapenemases are not reliably inhibited by vaborbactam, but demonstrate variable susceptibility to avibactam and relebactam [49]. Gram-negative organisms may also produce aminoglycoside-modifying enzymes (AMEs), including acetyltransferases, nucleotidyltransferases and phosphotransferases, all of which inactivate aminoglycosides [50, 51]. Less commonly, fluoroquinolone-, macrolide- and rifampin-inactivating enzymes may be produced by various Enterobacterales, Pseudomonas and Acinetobacter species [33].
Target site modificationAlthough alterations in penicillin-binding proteins are commonly observed with Gram-positive organisms, Pseudomonas species may harbour such mechanisms, leading to reduced β-lactam susceptibility [33]. Modification of fluoroquinolone target proteins occurs by mutations of gyrA/gyrB and parC/parE genes [32, 52], while aminoglycoside binding site alteration occurs by mutation of the 30S ribosomal subunit or by production of 16s rRNA methyltransferases via organisms harbouring armA and various rmt genes [50, 51]. Additionally, resistance to colistin is observed with alterations in the negative charge of the bacterial cell membrane, which prevents binding and insertion of the positively charged drug at its target site [33].
Efflux pumpsIn nonfermenting Gram-negative bacteria, such as Pseudomonas species, resistance to β-lactams and fluoroquinolones is often caused by overproduction of the MexAB-OprM efflux pump, whereas overproduction of MexXY-OprM transporters results in resistance to aminoglycosides [32, 52, 53]. This overproduction is mediated by mutations in the nalB, nfxB and nfxC genes [52].
Porin mutationsPorin channels play an important role in antibiotic uptake into Gram-negative bacteria [32]. Mutations resulting in inactivation or decreased expression of porin proteins, most notably OprD, leads to decreased permeability of several antibiotic classes including carbapenems, aminoglycosides and fluoroquinolones [32, 51, 52].
Gram-negative bacteria can harbour multiple resistance mechanisms that manifest clinically as a multidrug resistance phenotype [32]. Multiple β-lactamases may coexist, such as the combination of carbapenemases with an AmpC or ESBL, which, along with the presence of an AME-, ESBL- and AmpC-producing organism also having porin mutations results in a slowed rate of bacterial penetration, facilitating enzymatic hydrolysis of β-lactams, including carbapenems [44]. Additionally, the presence of such β-lactamases in combination with various drug efflux pumps or alterations in target sites can also confer resistance to other classes of antibiotics, and thus the manifestation of multidrug resistant organisms [44]. The growing prevalence and global impact of such multidrug resistance is of great concern.
Impact of Gram-negative antibiotic-resistant pulmonary infection on outcomesThere is a consensus that timely and appropriate antibiotic treatment, defined as an antibiotic regimen with in vitro activity against the causative pathogens, is a necessary first step to optimise the outcomes of patients with serious infections, especially in the ICU setting [54–58]. The guiding principle of timely appropriate antibiotic treatment of serious infections is endorsed by the most recent 2021 Surviving Sepsis Campaign International Guidelines for the management of sepsis and septic shock with a strong recommendation [59]. Kumar et al. [58] demonstrated that for every hour's delay until appropriate antibiotic administration, crude mortality increased by more than 10%. Vazquez-Guillamet et al. [60] studied >1000 subjects with septic shock and calculated that appropriate therapy enhanced the likelihood of survival at least three-fold. More importantly, this converted into a number needed to treat (NNT) to save one life of only 4, and the prevalence-adjusted pathogen-specific NNT to prevent one patient death was lowest for infections caused by MDR bacteria (NNT=20) [60]. Thus, both appropriate antibiotic selection and their timely administration are necessary to optimise patient outcomes.
Bassetti et al. [61] summarised the data regarding appropriate early therapy of serious infections in a meta-analysis of 114 studies of appropriate therapy in severe bacterial infections including pneumonia. Appropriate initial antibiotic therapy not only significantly reduced in-hospital mortality (OR 0.44, 95% CI 0.38–0.50), but also reduced length of stay by >2.5 days [61]. Delayed appropriate antibiotic therapy has also been shown to be associated with greater mortality in VAP [62]. Moreover, several nosocomial pneumonia registration trials for agency drug approval have demonstrated that when inactive antibiotic therapy is administered to patients, primarily inactive due to inadequate dosing or antibiotic exposure, mortality is increased [63–66]. These studies confirm the importance of delivering an appropriate and adequately dosed antibiotic regimen to patients with bacterial lung infections in order to improve patient outcomes, especially hospital survival.
The strongest evidence supporting early appropriate antibiotic therapy for severe infections comes from a randomised prospective trial examining this issue. The MERINO trial compared piperacillin-tazobactam to meropenem in patients with bloodstream infection caused by ceftriaxone-nonsusceptible E. coli or K. pneumoniae; in essence, it was a trial of appropriate versus inappropriate therapy [57]. The source of infections was primarily urinary tract and intra-abdominal with <5% being pneumonia. Noninferiority of the piperacillin-tazobactam arm could not be established, with 12.3% of 187 patients randomised to piperacillin-tazobactam dying at 30 days compared with 3.7% of 191 patients randomised to meropenem [57]. Microbiological failures were also more common among patients randomised to receive pipercillin-tazobactam when infected with Amp-C overexpressing Gram-negative bacteria [67].
MDR and XDR Gram-negative bacterial infections are more likely to initially receive inappropriate antibiotic treatment, resulting in worse outcomes. Zilberberg et al. [68] studied 1064 patients with Gram-negative bacteraemia, of whom 351 (29.2%) did not survive hospitalisation. Nonsurvivors were significantly more likely to have infection with an MDR isolate and to have received inappropriate initial antibiotic therapy. A multivariate analysis demonstrated that presence of infection with an MDR isolate was strongly associated with the receipt of inappropriate initial antibiotic therapy (adjusted OR 13.05, 95% CI 7.00–24.31). The same group of investigators also evaluated 1423 patients from United States hospitals with pneumonia or sepsis due to A. baumannii [69]. Harbouring MDR A. baumannii increased the risk of receiving inappropriate initial antibiotic therapy more than five-fold, and inappropriate initial antibiotic therapy nearly doubled hospital mortality. More recently, Martinez-Nadal et al. [70] evaluated 1615 episodes of bacteraemia associated with neutropenia, of which 394 (24%) received inappropriate initial antibiotic therapy. Patients with MDR Gram-negative bacteria, accounting for 221 (14%) of all isolates, were more likely to receive inappropriate initial antibiotic therapy (39% versus 7%, p<0.001). Overall mortality was also higher in patients with Gram-negative bacteraemia who received inappropriate initial antibiotic therapy (36% versus 24%, p=0.004). Thus, it can be seen that the presence of antibiotic resistance among Gram-negative bacteria is associated with greater initial administration of inappropriate antibiotic therapy, which in turn leads to greater mortality.
The problem of antimicrobial resistance appears to be escalating. Data from the National Healthcare Safety Network from 2015 to 2017 show that among P. aeruginosa isolates in ICU patients, 26.3% were resistant to carbapenems, 26.5% were resistant to extended-spectrum cephalosporins and 27.1% were resistant to fluoroquinolones, with 18.6% classified as MDR [71]. Increasing resistance has led clinicians to employ antimicrobial “cocktails” in the hopes of salvaging patients with MDR/XDR infections [72, 73]. Verona integron-encoded metallo-β-lactamase-positive P. aeruginosa (VIM-PA) has emerged as one of the most resistant Gram-negative pathogens, along with carbapenem-resistant A. baumannii. In a study from the Netherlands, VIM-PA bacteraemia was associated with greater mortality compared to carbapenems-susceptible infection [74]. Thus, it is evident that increasing future infections with MDR/XDR pathogens will be associated with greater morbidity and mortality, particularly in respiratory infections. Figure 4 offers an approach to the clinical management of antibiotic-resistant Gram-negative infections in the hospital setting.
 FIGURE 4
FIGURE 4 Clinical approach to the treatment of microbiologically confirmed or suspected multidrug-resistant (MDR) Gram-negative bacterial infection. XDR: extremely drug-resistant; CTZ-TAZ: ceftolozane/tazobactam; CEF-AVI: centazidime-avibactam; CEFID: cefiderocol; IMI-REL: imipenem-cilastatin; AG: aminoglycoside; RIF: rifampicin; ESBL: extended-spectrum β-lactamase producing; MER: meropenem; FQ: fluoroquinolone; ERT: ertapenem; AMO-CLA: amoxicillin-clavulanate; FOS: fosfomycin; NTF: nitrofurantoin; TMP-SMX: trimethoprim-sulfamethoxazole; CRAB: carbapenem-resistant Acinetobacter baumannii; ERA: eravacycline; PLZ: plazomicin; COL-PMX: colistin-polymyxin B; TIG: tigecycline; CRE: carbapenem-resistant Enterobacterales; AZT: aztreonam; MER-VAB: meropenem-vaborbactam. #: high risk of mortality is considered >15%.
Novel and pipeline antibioticsSeveral antibiotics have been introduced to combat Gram-negative resistance and expand coverage against MDR Gram-negative infections, including respiratory tract infections (table 3) [75]. Their precise role in empirical and definitive treatment is beyond the scope of this review; however, well-written reviews and guidelines highlight the importance of pertinent risk factors, current and prior culture and susceptibility data, local resistance rates, available clinical and/or in vitro data and other factors [75–77]. Moreover, the use of novel antibiotics as combination therapy provides a unique approach to the treatment of MDR Gram-negative infections. For example, ceftazidime/avibactam has broad activity against serine β-lactamases; however, it is hydrolysed by MBL. In contrast, aztreonam is resistant to MBL hydrolysis, but susceptible to serine β-lactamases. Available in vitro and in vivo data have demonstrated synergistic activity with this combination against MBL-producing Gram-negative organisms [78–81].
TABLE 3Novel and pipeline antibiotic treatment options for Gram-negative respiratory infections
Pipeline antibiotics will provide extended Gram-negative coverage and may become useful options for the treatment of MDR Gram-negative respiratory infections (table 3). For example, the addition of the novel β-lactamase inhibitor zidebactam to the β-lactam cefepime improves activity against KPC, ESBL, OXA-48 and MBLs [82, 83]. The expanded spectrum provided by and mechanism of β-lactam/β-lactamase inhibitor combinations differs among agents [49, 82]. Avoidance of aminoglycoside-inactivating enzymes affords the novel synthetic aminoglycoside, arbekacin, improved activity against MDR Gram-negatives and Gram-positives [84]. Murepavadin highlights a novel drug class of outer membrane protein targeting antibiotics by binding to the lipopolysaccharide transport protein D present on the outer membrane of MDR P. aeruginosa [85]. Initially encouraging phase I and II data have been stifled by the termination of the phase III PRISM-MDR trial due to safety concerns surrounding adjunctive use of murepavadin in patients with MDR P. aeruginosa VAP [86]. These data emphasise the need for ongoing study of all novel antimicrobials for MDR Gram-negative respiratory infections.
Monoclonal antibodiesThe success of monoclonal antibodies (mAbs) for treatment and/or prevention of various diseases has reinvigorated consideration in treatment and prevention of bacterial infections. Antibacterial mAbs may disrupt bacterial infections via several mechanisms; however, targeting polysaccharides, proteins or toxins associated with proliferation, adhesion, host immunity protection, host cell damage and/or increased virulence are most common (table 4) [87, 88]. Encouraging results in prevention or treatment of Clostridioides difficile, inhalational anthrax and MRSA infections have been reported; however, data in Gram-negative respiratory infections remain preliminary [89–91].
TABLE 4Monoclonal antibody treatment options for Gram-negative respiratory infections
The mAb KB001-A targets the PcrV protein on P. aeruginosa, reducing levels of toxin transport and lung injury/inflammation in animal models [92]. In cystic fibrosis patients, KB001-A was not associated with decreased time to need for antibiotics, but increased forced expiratory volume in 1 s and reduced inflammatory markers [92]. A randomised phase II trial of mechanically ventilated patients colonised with P. aeruginosa reported no difference in incidence of treatment-related adverse events, but reduced incidence of P. aeruginosa pneumonia development in patients receiving KB001-A [93]. The EVADE trial is assessing the ability of MEDI3902, a bivalent/multi-target mAb, to prevent P. aeruginosa infections in patients mechanically ventilated colonised with P. aeruginosa [94]. Panobacumab and AR-105 target polysaccharides produced by P. aeruginosa and are under investigation as adjuncts with standard-of-care antibiotics for nosocomial P. aeruginosa infections [95, 96]. Continued advancements in molecular modelling and bioinformatics are likely to yield additional targets for pathogens associated with MDR Gram-negative respiratory infections [97].
Antibacterial mAbs show great promise for the treatment and prevention of Gram-negative bacterial respiratory infections. Noteworthy advantages include target specificity, reduced risk of resistance emergence, minimal disturbance of gut microbiota and unique pharmacokinetic profiles. Still, limitations related to target specificity, cost, antibody development, administration, availability and clinical data hamper use in routine clinical practice [87]. Results of ongoing clinical trials are awaited to properly assess their efficacy, safety and appropriate place in therapy.
Bacteriophage therapyBacteriophages (phages) are the most abundant viruses/organisms on Earth and can infect and replicate within bacterial cells, causing lysis [98]. Leveraging phages to target pathogenic bacteria has seen a resurgence due to threats of antimicrobial resistance and advancements in molecular engineering (table 5). Bacteriophages offer several presumed advantages related to their specificity and ubiquity, including minimal effects on microbiota, safety profile, administration flexibility, and self-limiting properties. However, there is a paucity of data in these regards, combined with unknowns surrounding monitoring, toxin release, antibody production and inevitable development of resistance. Considerations of the unique interplay with bacteria, antibiotics and the immune system are necessary to optimise delivery and production, minimise or leverage resistance and understand their safety.
TABLE 5Bacteriophage therapies for Gram-negative respiratory infections
Clinical evidence for phages is limited to case series and reports. A systematic review highlighted the potential of phages in difficult-to-treat respiratory infections [99]. One such case described success of adjunctive phage therapy for the treatment of a P. aeruginosa VAP and bronchopleural fistula after thoracotomy, resulting in both clinical improvement and bacterial eradication at 6-month follow-up [100]. Phage therapy has exceptional potential for patients with MDR Gram-negative respiratory infections; however, well-designed clinical trials are still necessary.
OligonucleotidesAntimicrobial oligonucleotides are synthetic nucleic acid sequences able to silence genes vital to bacterial survival (table 6) [98]. In vivo and in vitro data suggest that oligonucleotides may serve as a future treatment modality of severe Gram-negative respiratory infections [98]. Oligonucleotides targeting the genes MexB, blaNDM-1, and CTX-M-15 have resulted in improved minimum inhibitory concentration (MIC) profiles of common antimicrobials [101, 102]. Like mAbs and phages, oligonucleotides offer a target-specific method with potentially longer-lasting effects and reduced resistance development; however, limitations surrounding their adequate delivery, use with antibiotics and unestablished safety and efficacy profile highlight the need for continued study.
TABLE 6Oligonucleotides for Gram-negative respiratory infections
Role of stewardship in curbing MDR spreadAntimicrobial stewardship programmes (ASPs) are an essential tool in helping prevent the spread of MDR respiratory infections, and involve a multidisciplinary approach to ensure that patients receive appropriate initial antibiotic therapy, appropriate dosing and route of administration, as well as sufficient duration of antimicrobial therapy [103, 104]. The goal of ASPs is to improve clinical outcomes of patients, decrease costs associated with antimicrobial use and control the spread of resistant organisms [105–107]. It is increasingly recognised that ASPs are an essential part of management of hospitalised patients, especially those in the ICU, and its use is recommended by the Infectious Diseases Society of America and international sepsis guidelines [59, 104, 108].
The ICU is an essential area for ASPs, due to the frequent use of antibiotics, high levels of MDR organisms and implications for poor clinical outcomes in patients treated with inappropriate antibiotics [105–107, 109]. Despite more than half of patients in the ICU having suspected or confirmed infection, antibiotic use is in the ICU is still frequently overly broad or too narrow [1, 106]. The consequence of overly broad antimicrobial use in the ICU leads to the development of antimicrobial-resistant organisms, which is associated with increased mortality [1, 110, 111]. Infection with an MDR organism is also associated with increased all-cause mortality and increased likelihood of hospital readmission [112].
Respiratory infections in critically ill patients represent an important area for antimicrobial stewardship, and pneumonia is the most common infection found in the ICU [1]. Initial antibiotic therapy that is inappropriately narrow has been shown to be associated with an increased risk of mortality, while initial antibiotic therapy that is inappropriately broad-spectrum has also been associated with increased risk of mortality [113]. Appropriate duration of antibiotic therapy is also an important aspect of ASPs, as unnecessarily prolonged courses of antibiotics are associated with adverse patient outcomes and the development of MDR organisms [111, 113]. Teshome et al. [111] showed that each additional day of broad-spectrum antibiotic use directed against P. aeruginosa was associated with an increased risk of development of MDR infection. The benefits of shorter courses of antibiotics for pneumonia were shown in a systematic review and meta-analysis of >1000 patients with hospital-acquired pneumonia (HAP) and VAP found that a shorter course of antibiotics resulted in increased 28-day antibiotic-free days as well as a reduction in recurrent VAP due to MDR organisms [114].
In a retrospective cohort analysis of >17 000 patients admitted to the hospital with sepsis and positive cultures, Rhee et al. [113] found that overly broad empirical antibiotic therapy was associated with increased mortality, with an estimated 20% increase in the odds of death. The adverse effects of overly broad antibiotic therapy were also seen in a retrospective study of over 1900 patients admitted to the hospital with pneumonia, which showed that the use of broad-spectrum antibiotics in community-onset pneumonia was associated with longer hospital stays, increased rates of Clostridium difficile infection and increased mortality [115]. Unnecessarily broad antibiotic therapy has the unfortunate consequence of disrupting the microbiome, which refers to the numerous bacteria, fungi and viruses that exist in a symbiotic relationship with a human body [116, 117]. The use of broad-spectrum antibiotics has been shown to alter the beneficial bacteria that make up the microbiome, leading to dysbiosis and placing the patient at risk for colonisation with MDR organisms and subsequent opportunistic infections [116]. ASPs can be beneficial in preserving the host microbiome by limiting unnecessary antimicrobial use and limiting length of therapy to an appropriate duration. The importance of appropriate timing, duration and spectrum of antimicrobial activity is detailed in figure 5.
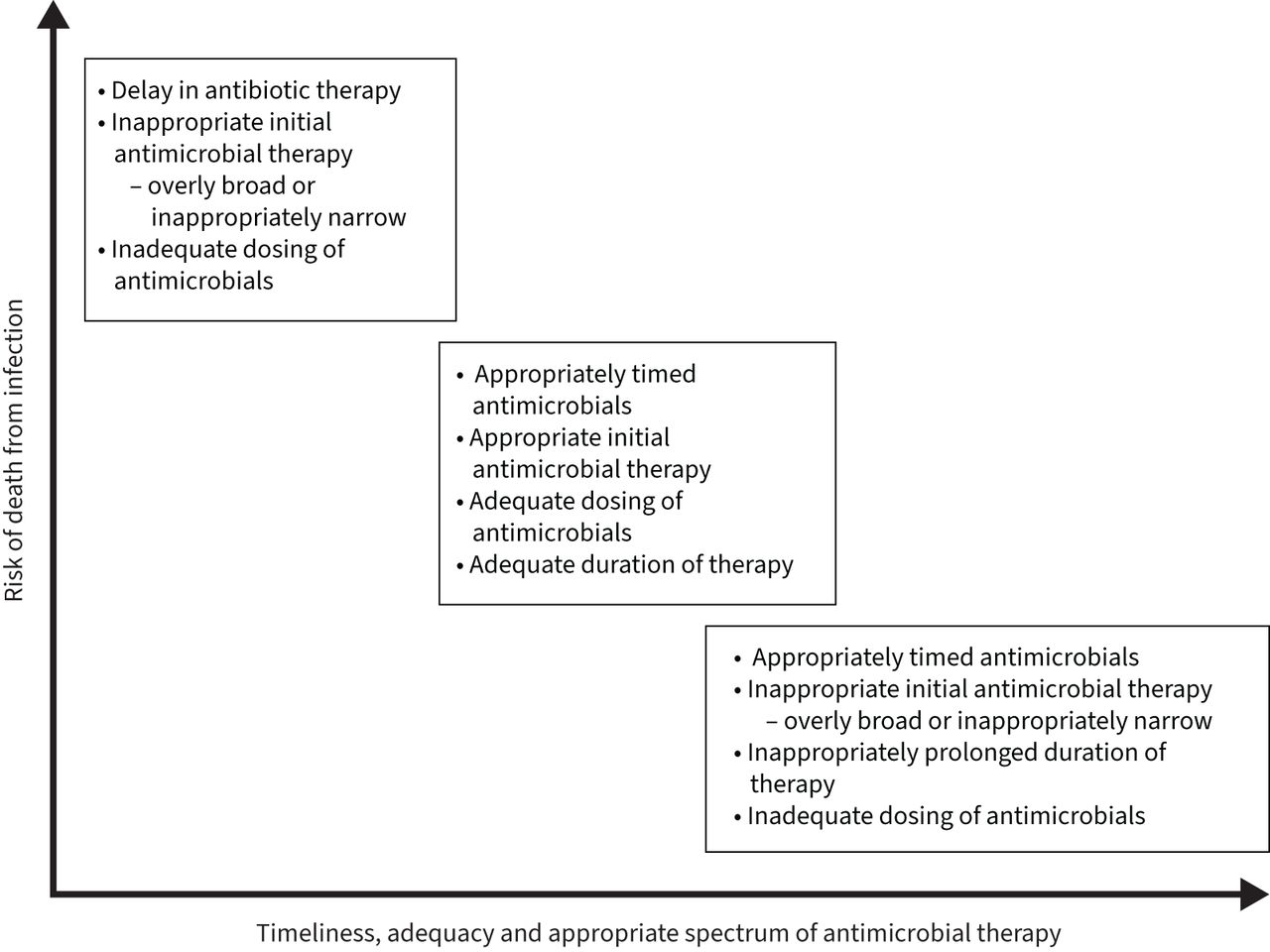 FIGURE 5
FIGURE 5 The importance of appropriate timing, duration and spectrum of antimicrobial activity.
The success of an ASP is dependent on several key elements and must include a multidisciplinary approach that utilises the expertise of ICU physicians, infectious disease physicians, pharmacists and microbiologists [104, 106]. Several of the key elements that are recommended by the United States Centers for Disease Control and Prevention for a successful ASP are detailed in table 7 [104]. Prospective audit and feedback of antibiotic use is an effective aspect of an ASP, which functions as a review of broad-spectrum antibiotic therapy leading to antibiotic de-escalation when appropriate [103, 104]. It has been shown to be associated with a reduction in use of broad-spectrum antibiotics in critically ill patients, reduction in resistance of Gram-negative organisms, with no adverse consequences on mortality or ICU length of stay [103, 108, 118].
TABLE 7Key components of an effective antimicrobial stewardship programme
Clinical decision support systems are another important part of an effective ASP, which are designed to incorporate numerous clinical and patient-specific data in order to provide more information to clinicians so that they can make more appropriate decisions regarding antimicrobial therapy. Previous studies have shown that clinical support tools that make antibiotic recommendations using patient data and antibiogram information can decrease overall use of antibiotics [119] as well as improve initial antibiotic therapy in patients with sepsis [120].
Rapid diagnostic testing to identify a source of infection remains an area of need for ASPs in the ICU. Earlier identification of causative organisms, as well as rapid diagnostics that are sensitive in ruling out specific organisms that are treated with broad-spectrum antimicrobials can lead to rapid de-escalation of antibiotics [
留言 (0)