
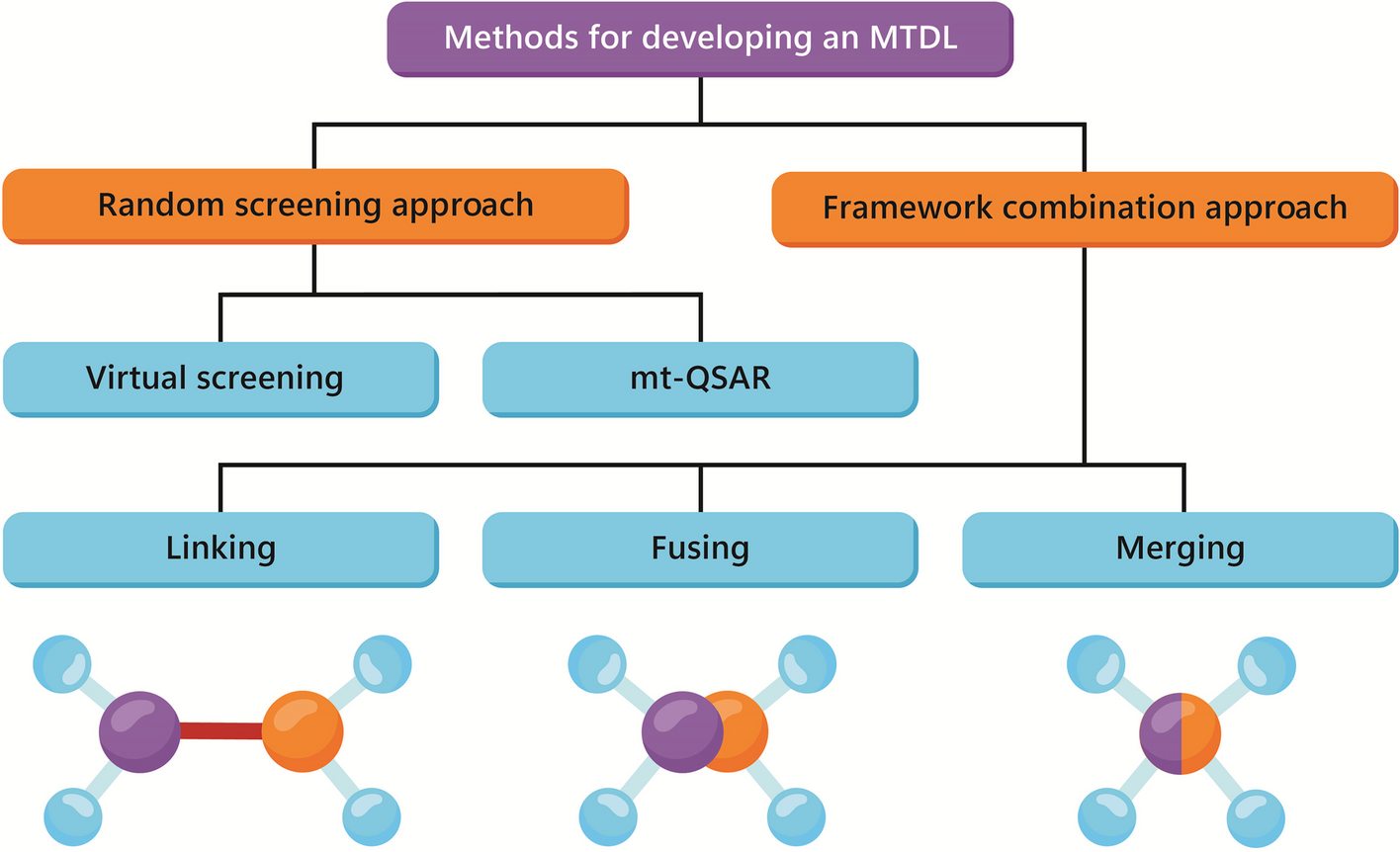
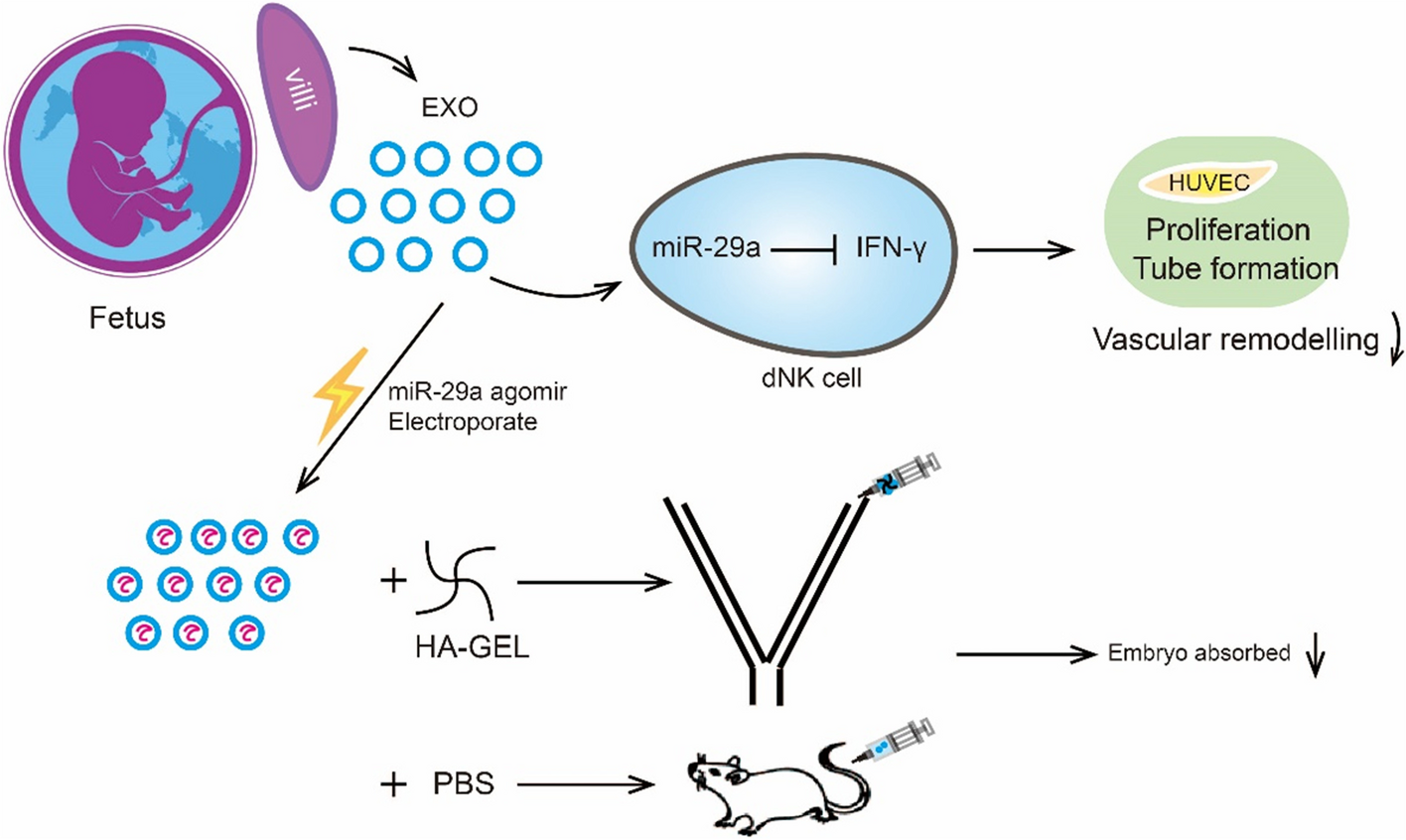
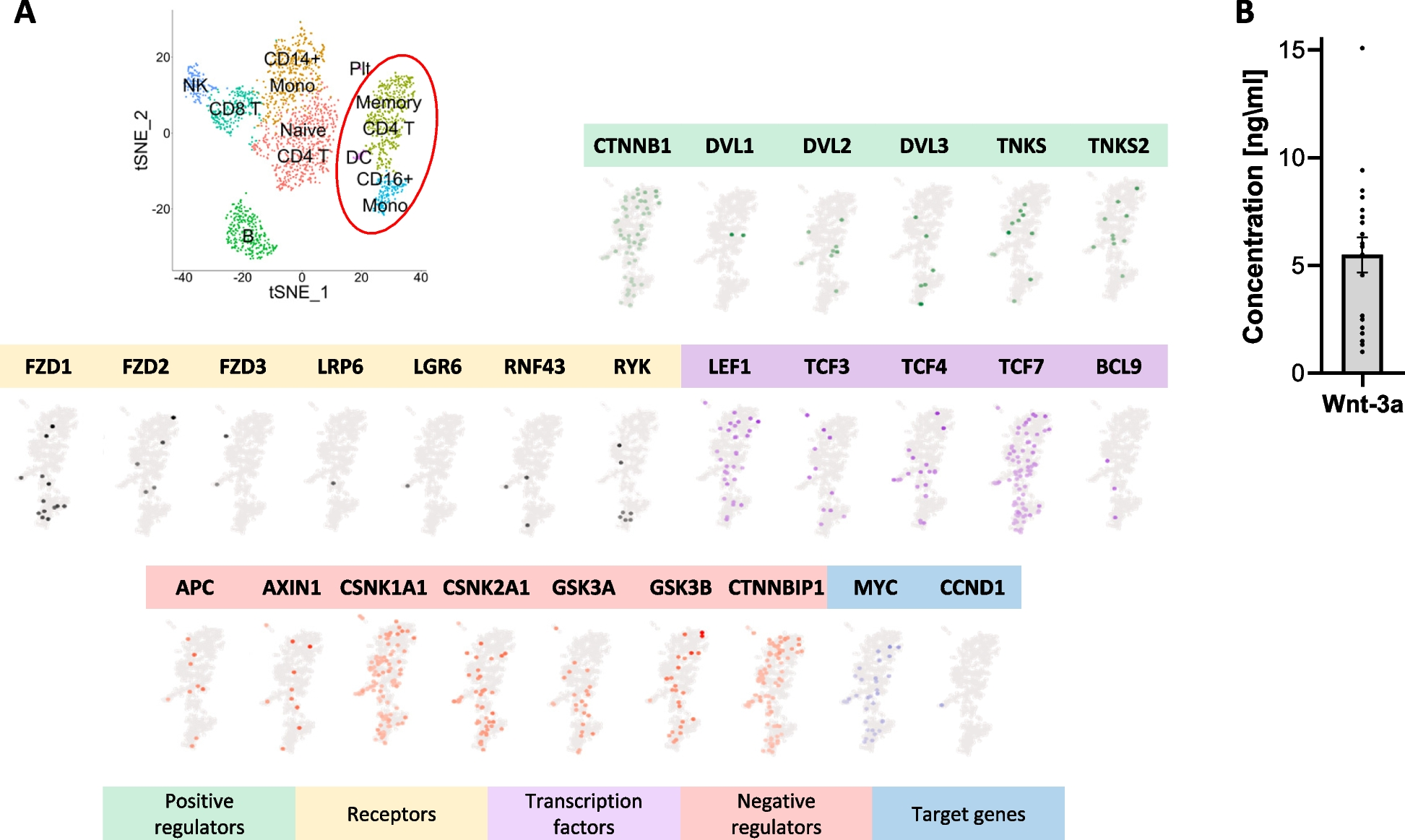
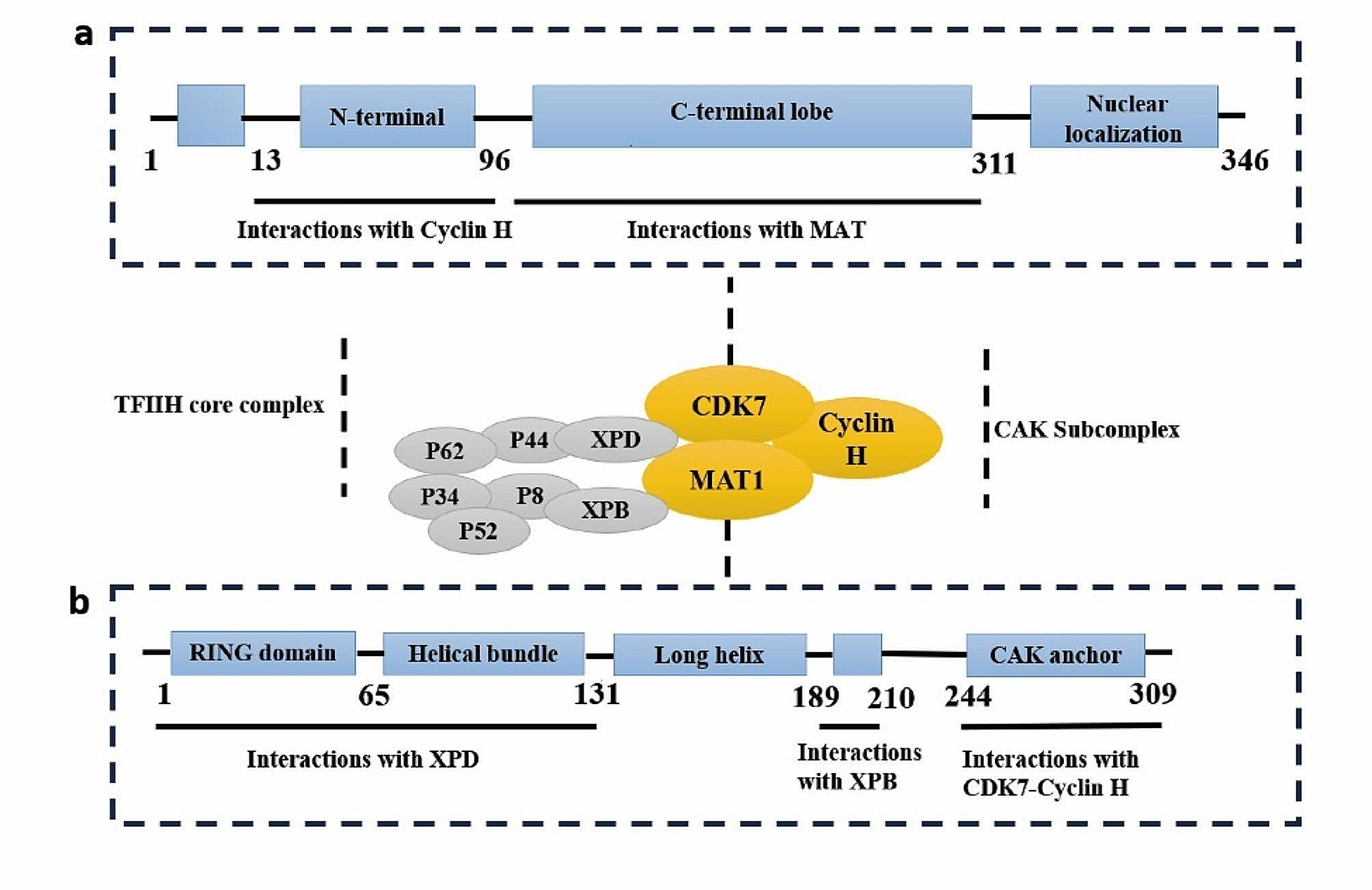
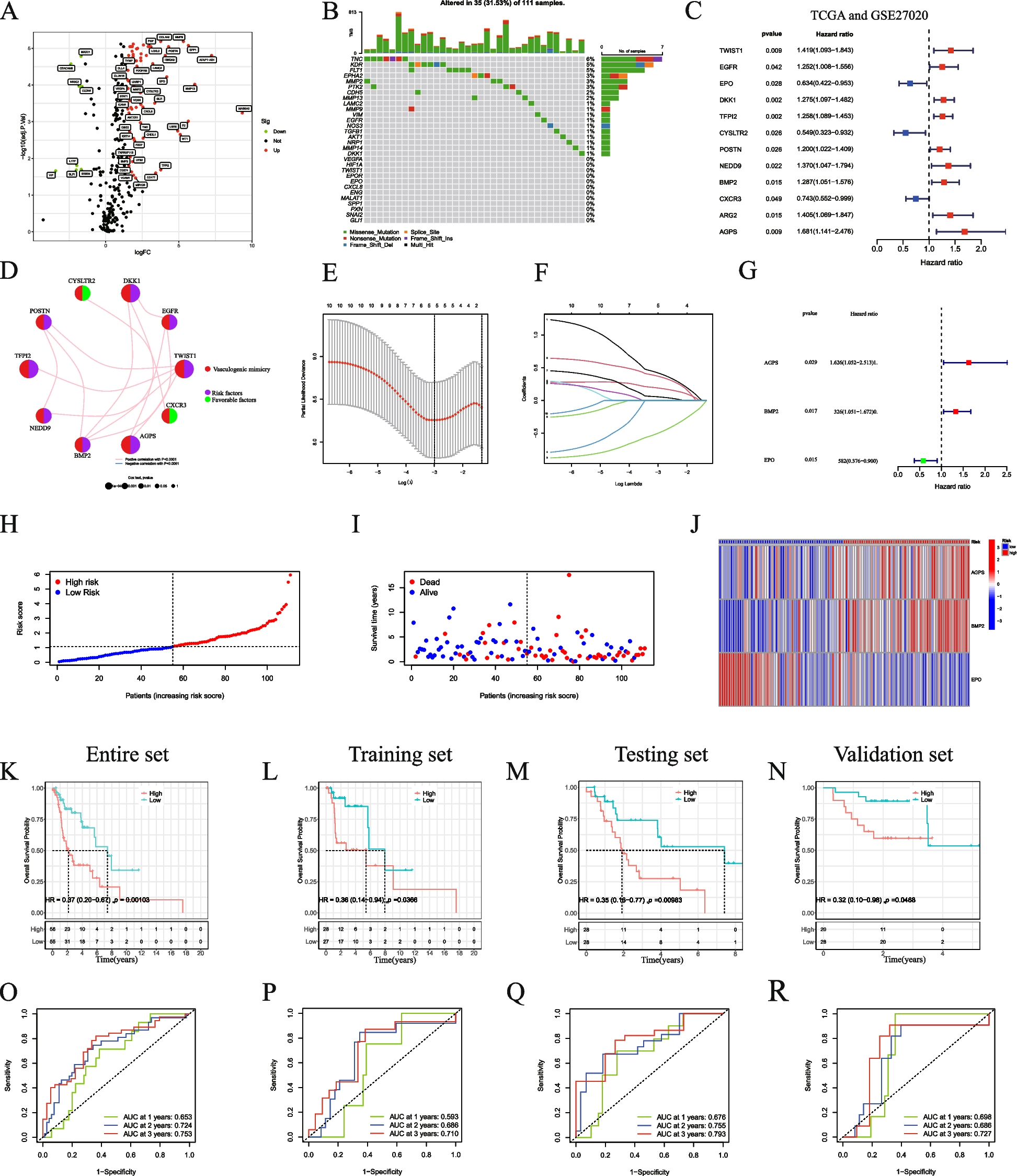
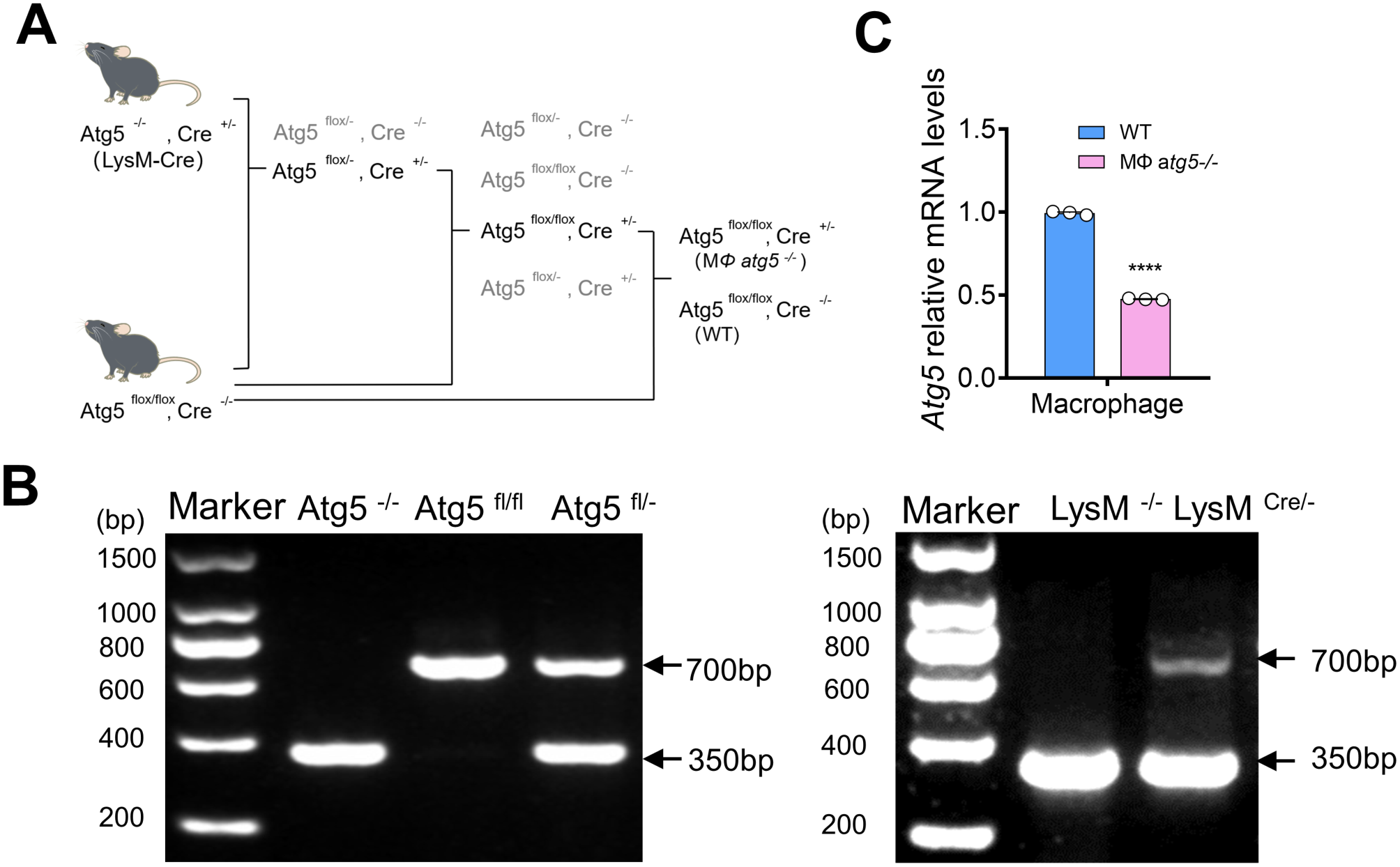
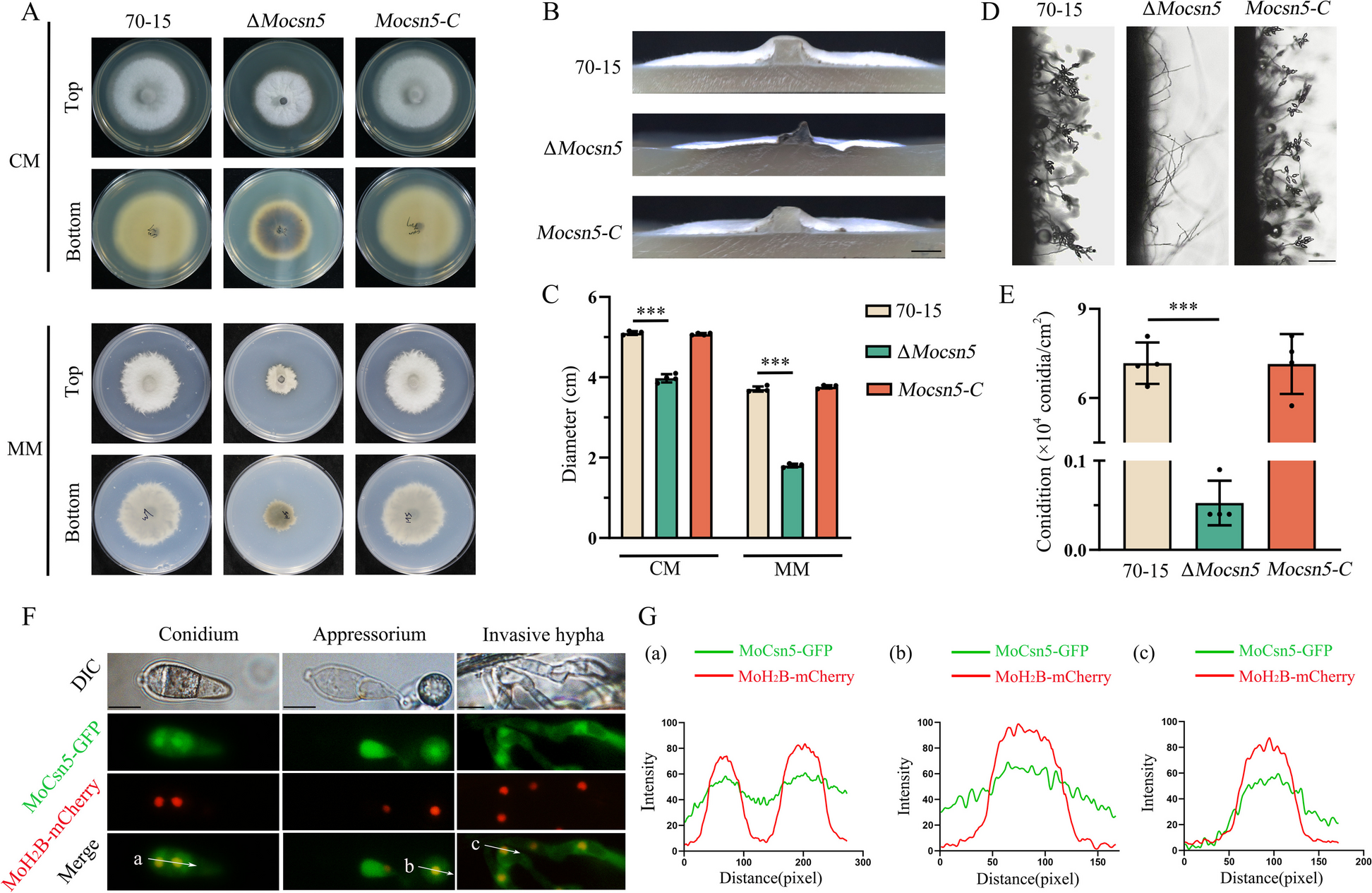
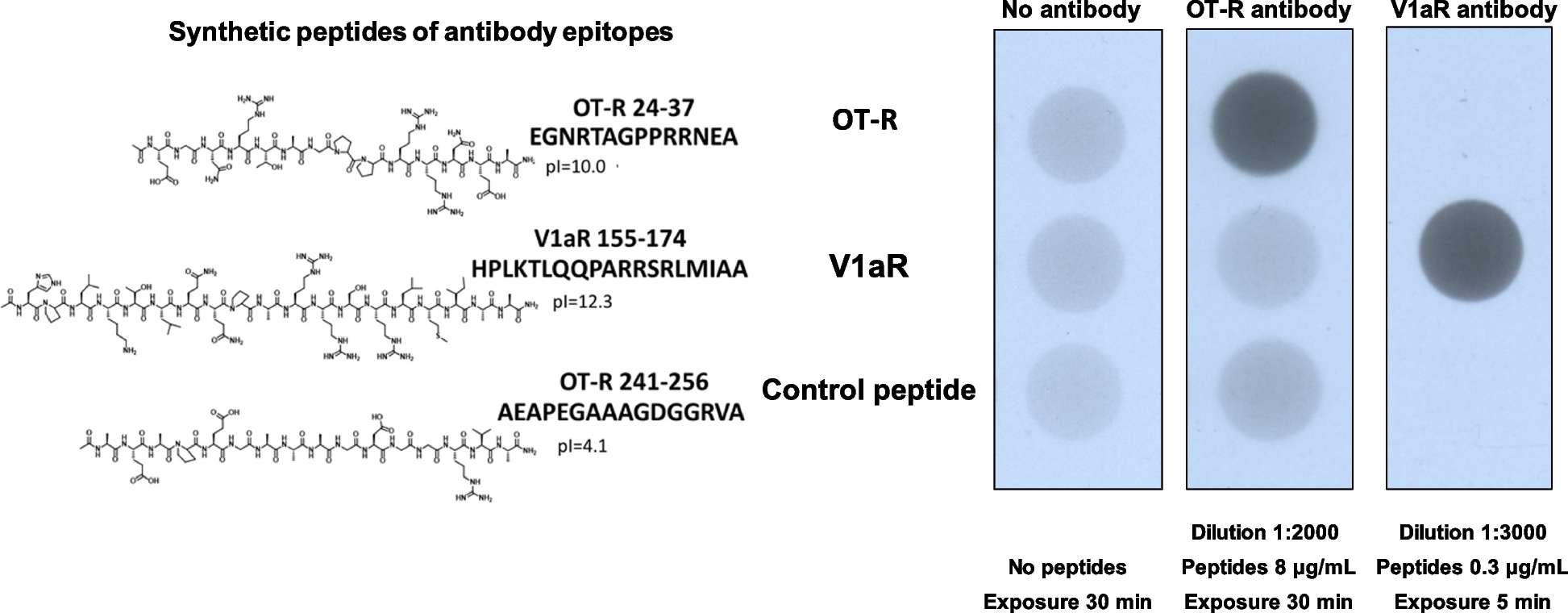
記住我
Since acinar morphology has been shown to be affected by both MRTF activity and tissue stiffness, we wanted to assess whether they cooperate during acini formation. First, we analysed changes in three-dimensional acinar morphogenesis induced by various matrix compositions of different stiffness. We thus cultured human mammary MCF10A epithelial cells on solidified Matrigel, Matrigel/collagen I mixtures (75%/25%, 50%/50%, 25%/75%) and pure collagen I. The increasing collagen I content leads to increasing stiffness of the solidified matrix with an elastic Young's modulus from about 0.1 kPa for soft Matrigel to about 4 kPa for stiffer collagen I [4]. To enable 3D acini formation, the overlay medium contained a constant amount of 2% matrigel in all conditions. The stiffness-dependent effects on the size and shape of the acini were analysed on day 4, 8 and 14 after cell seeding. Immunofluorescence analysis revealed an increasingly irregular shape and single- or multi-cellular outgrowth already in early 3D acinar structures on stiffer matrices (Fig. 1A). The average acini diameter, however, remained comparable, regardless of matrix composition (Fig. 1B). A moderate and rather expected increase of the acini diameter was observed correlating with the cultivation duration.
Fig. 1
Correlation of increasing matrix stiffness with protrusion formation of MCF10A acini. 3D cultures of MCF10A cells on the indicated Matrigel/Collagen I mixtures (100:0, 75:25, 50:50, 25:75, 0:100) were analysed by microscopy. A Phalloidin (green), Laminin V (red) and nuclei (DAPI, blue) staining of acinar structures at day 4 after seeding on matrices with increasing Collagen I percentages. Arrowheads indicate protrusions. The corresponding increase of Young's elastic modulus of the matrix is depicted below. B Diameters of individual acini (diamonds) measured at day 4, day 8 and day 14. Black horizontal lines indicate mean diameter and SEM (whiskers) of three biologically independent replicates. C Quantification of protrusion formation at day 4, day 8 and day 14. At least 11 acini were assessed each day, the percentage of protrusion-positive acini calculated, and the entire experiment repeated 3 times. D Calculated roundness index of individual acini (diamonds). Statistical significance compared to acini formed on pure Matrigel was determined using a one-way ANOVA and the means for each time point. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. Error bars, SEM (n = 3). Scale bars, 50 μm
Quantitative analysis showed that stiffer collagen I-rich matrices (75–100%) promote significant protrusion formation independently of the cultivation time (Fig. 1C), consistent with previous observations [4]. For example, protrusion formation on collagen I increased by 23%, 17.8% and 17.9% compared to Matrigel on day 4, day 8 and day 14, respectively. Protrusion formation was accompanied by a more irregular acini shape as demonstrated by the significantly lower roundness of acini grown on Matrigel compared to those cultured on 75–100% collagen I (Fig. 1D).
Similar morphological changes were observed for human MCF7 breast cancer cells cultured on solidified Matrigel/collagen I mixtures for 4 days (Additional file 1: Fig. S1). The average diameter of the MCF7 spheroids remained constant independent of matrix properties. Also in this cell line, high collagen I content in the culture matrix promoted acinar outgrowth. Compared to soft Matrigel layer, 4.2–13.4% of the spheroids grown on matrix mixtures containing 50%, 75% or 100% collagen I developed protrusions.
Stiffness but not underlying matrix composition governs acini formationIncreasing the percentage of collagen I in the base layer does not only change the stiffness of the matrix, but also its molecular composition and thereby, potentially, the ligands available for cellular receptors such as integrins. Thus we investigated whether the observed morphological changes were mediated by the increased collagen I content or by the increased stiffness of the underlying matrix. For this purpose, we firstly analysed the formation of MCF10A acini on Matrigel (MG) and collagen I (COL1) matrices, which were both thinly precoated with collagen IV (COL4), a constituent component of the basal lamina adjacent to mammary acinar structures.
Collagen IV coating did not significantly alter the mean diameter of MCF10A acini grown on either Matrigel or collagen I matrices for 4 days, as compared to the uncoated matrix (Fig. 2A). As before, protruding acini significantly increased on the stiffer COL1 base layer, but additional precoating with COL4 did not result in a significant reduction of protrusion formation (Fig. 2B). Rather, fluorescence microscopy revealed profound differences of the acini grown on Matrigel or collagen I, even though both were coated with collagen IV (Fig. 2C). This suggests that it is not the presentation of collagen I on the matrix which promotes protrusion formation.
Fig. 2
Acinar protrusions are independent of composition but depend on stiffness of matrix. A, B, C Matrigel (MG) or collagen I (COL1) gels were thinly coated with collagen IV (COL4) or left uncoated (wo COL4) as indicated. Subsequently MCF10A cells were seeded on top and acinar diameter A and protrusion formation B was analysed 4 days post seeding. C Representative micrographs of acini stained with laminin V (red), actin filaments with phalloidin (green) and cell nuclei with DAPI (blue) on day 4 of acini morphogenesis. Arrowheads indicate protrusions. D, E, F MCF10A cells were seeded on pre-cast acrylamide gels with defined stiffness of 0.2 kPa and 4 kPa coated with Matrigel, respectively. Acini diameter D and average percentage of protruding acini (E) were determined 4, 8 and 14 days post seeding as before. F Representative micrographs of acini on day 4 after seeding on the matrix indicated. Statistical significance was determined by ANOVA using the means of 3 independent experiments, each analysing ≥ 22 acini per condition. G, H Relative mRNA expression of ACTA2 G and VIM H normalised to ALAS and GAPDH. Statistical significance according to an unpaired Student’s t-test (n = 4). *p ≤ 0.05, **p ≤ 0.01. Error bars, SEM. Scale bars, 50 μm
In a second experimental approach, we seeded MCF10A cells onto base layers of commercially available pre-cast acrylamide gels with defined stiffness of 0.2 kPa and 4 kPa. Both gels were coated with Matrigel to allow cell adhesion. Diameter and shape of the acini were examined 4, 8 and 14 days after cell seeding. The results confirmed that an increased stiffness of 4 kPa of the underlying matrix did not significantly change the diameter of the acini structures formed (Fig. 2D). However, formation of protrusions was triggered almost exclusively on the pre-cast acrylamide gels with high stiffness (Fig. 2E and F). These results suggest that the role of particular matrix components or diffusable ligands can be neglected for protrusion formation on collagen I matrices under our experimental conditions, but that physical stiffness causes the increased protrusion formation of MCF10A acini.
To corroborate the stiffness-induced malformation of acini with mammary epithelial characteristics, we analysed the expression of the markers smooth muscle alpha actin (ACTA2) and vimentin (VIM). mRNA was collected from acini grown for 4 days on the defined "soft" acrylamide gels with 0.2 kPa, or from those with 4 kPa which permitted protrusion formation in around 15% of acini. Both markers were slightly upregulated in acini grown on the stiffer acrylamide gels with 4 kPa (Fig. 2G and H). Whilst ACTA2, a known MRTF target gene, was only induced by around 20 percent, the mesenchymal marker VIM was non-significantly elevated threefold, suggesting that the cells are changing their luminal epithelial fate when growing on stiffer matrices.
MRTF/SRF is required for stiffness-dependent protrusion formationAfter having validated our model of matrix stiffness, we next analysed MRTF/SRF activity in MCF10A acini grown on soft and stiff ECM gels. Using a promoter-luciferase reporter construct [35] dependent on transcriptional activation by the MRTF/SRF complex, we observed a moderate but significant increase in MRTF/SRF activity in 3D acini grown on stiff collagen I matrices for three days, compared to those grown on soft Matrigel (Fig. 3A). Accordingly, we hypothesised that activation of MRTF-A dependent transcription during acinar formation on stiff matrices might contribute to irregular spheroid formation and development of protrusions. We thus treated 3D acini cultures from day 4–8 during morphogenesis with the Rho/MRTF/SRF pathway inhibitor CCG203971 and analysed the morphological changes on matrices with increasing stiffness. Compared to the untreated control, CCG203971 did not significantly alter the average diameter of the acini regardless of the stiffness of the underlying matrix (Fig. 3B). However, the average protrusion formation of acini treated with CCG203971 was reduced by approximately 50% on stiffer collagen I matrices, exhibiting statistical significance on a Matrigel/collagen I mixture of 25:75 (Fig. 3C). Reduced protrusion formation correlated with reduced MRTF/SRF reporter activity following CCG203971 treatment (Additional file 1: Figure S2). Together, these results suggest a correlation of high MRTF/SRF activity and stiffness-dependent protrusion formation in 3D acini of MCF10A cells.
Fig. 3
Implication of the MRTF-SRF pathway in stiffness-dependent acini morphogenesis. A Relative MRTF/SRF luciferase reporter activity in transiently transfected MCF10A seeded on Matrigel (MG) or collagen I (COLI) and analysed at day 3 of acini morphogenesis (n = 4). B, C Effects of the Rho/MRTF/SRF pathway inhibitor CCG203971 (20 μM, day 4–8) on acini diameter and average percentage of protrusion-positive acini at day 8. Eleven or more acini were analysed per condition in 3 biologically independent replicates. B Individually measured acini diameter (diamond) and mean ± SEM (horizontal black line ± whiskers). C Percentage of protrusion formation. Significance was tested using an unpaired Student’s t-test. *p ≤ 0.05. Error bars, SEM
Matrix stiffness and MRTF activity cooperatively affect primary organoidsIn vivo, the mature lobulo-alveolar system is more complex, and acinar structures are already pre-existing. To partially account for the increased complexity and to further analyse stiffness-mediated effects on the mature acinus, we extended our experimental approach to primary acini from 8 to 12 week old female NMRI mice. Epithelial mammary acini were extracted and cultivated on 3D matrices for 5 days, followed by microscopical analysis of the acinar structures. When grown on soft Matrigel, organoids appeared round and hollow, whereas organoids on stiff collagen I gels lacked regularly shaped acinar morphology (Fig. 4A). As with MCF10A-derived acini, no stiffness-induced alteration of the diameter of the mature murine acini was observable. In contrast, increasing matrix stiffness correlated with significantly increased formation of protrusions (Fig. 4B, C). We also analysed luminal filling of the preformed acinar structures, but quantification of more than 40 organoids revealed only a minor increase of filled acini on stiffer matrices under these experimental conditions (Fig. 4D).
Fig. 4
Matrix stiffness and MRTF/SRF activity cooperate on protrusion formation in mature murine acini. Acinar organoids were extracted from the mammary gland of 8–12 week old female NMRI mice. Organoids were seeded on matrices with increasing stiffness, and morphologies were analysed 5 days after seeding by microscopy. A Representative images of primary murine acini stained for nuclei (DAPI, blue) and actin (phalloidin, green) on matrigel or collagen I matrices 5 days after seeding. Arrowheads indicate protrusions. Scale bars, 50 μm. Individual acini diameter B, percentage of protrusion-positive acini C and percentage of filled acini D were quantified (≥ 42 individual acini per condition, one-way ANOVA). (E) Representative images of acini after 4 days of treatment with DMSO or 20 μM CCG203971. Scale bars, 25 μm. Individual acini diameter F, percentage of protrusion-positive acini G and percentage of filled acini H (≥ 24 individual acini per condition, two-way ANOVA). *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. Error bars or whiskers, SEM
Subsequently we analysed whether the MRTF/SRF pathway and matrix stiffness functionally cooperate in protrusion formation, using the inhibitor CCG203971. As before, fluorescence microscopy revealed increasing protrusions and irregular morphology after mature acini were grown for 5 days on matrices with gradually increased stiffness. Vehicle control acini showed protrusions already on 50:50 mixtures of matrigel and collagen, which was partially reverted by CCG203971 treatment at this stiffness (Fig. 4E). No profound change in the diameter of CCG203971-treated organoids was measured compared to the vehicle control acini (Fig. 4F). Quantification confirmed a strong trend towards a CCG203971-mediated reduction of protrusion formation of preformed acini on matrices with intermediate stiffness. However, this inhibition seems to be overcome by cultivating the organoids on 100% collagen. Regarding filling of the acinar lumen, only a slight and non-significant effect of CCG203971 treatment was observed upon quantification of hollow structures under these conditions (Fig. 4H).
Overactivation of MRTF-A causes luminal filling regardless of matrix stiffnessFinally, we utilised a genetically encoded mouse model for an inducible gain-of-function of MRTF-A to analyse how MRTF-A affects primary murine acini cultured on soft and stiff matrices. The Tamoxifen-inducible CreERT2/loxP system was used to generate a conditional knockin of constitutively active MRTF-A in C57BL/6NCrl mice (A.K.S. and G.P., manuscript in preparation). The encoded MRTF-A lacks the RPEL1/2 motifs required for G-actin binding and repression. As described above, organoids were extracted from mammary glands of 8–12 week old transgenic mice which allow (tg) or not allow (wt, negative for CreERT2) for the expression of activated MRTF-A upon treatment with 4-hydroxy-tamoxifen (4OHT).
Organoids were treated with a vehicle control or 4OHT 24 h post seeding to initiate expression of activated MRTF-A. After 4 days of incubation, acini were fixed, stained and morphological changes were analysed. The stiff collagen I matrix lead to irregularly shaped organoids and the formation of protrusions, compared to round control acini cultured on the soft Matrigel (Fig. 5A, B). Strikingly, overactivation of MRTF-A by 4OHT treatment of tg organoids strongly promoted luminal filling, irrespective of the matrix stiffness. Quantification revealed that acini roundness was reduced on stiff matrices, whilst the average diameter of the organoids was largely unaffected, as before (Fig. 5B, C). The stiffer matrix slightly elevated the rate of luminal filling also in control organoids, consistent with previous reports [4], but this was strongly exacerbated in 4OHT-treated tg organoids showing a profound loss of hollow acinar structures (Fig. 5D). The number of protrusion-forming organoids was again around three times higher on the stiffer matrix (Fig. 5E). Interestingly, however, MRTF-A activation did not induce protrusion formation on matrigel, and even inhibited it on the stiff matrix, compared to untreated tg and wt acini. These results suggest that high MRTF-A activity is sufficient to cause luminal filling of preformed mammary acini, but that it is insufficient to promote protrusion formation.
Fig. 5
High MRTF-A activity causes luminal filling in primary mammary acini. Acinar organoids were extracted from the mammary gland of 8–12 week old female wildtype (wt) and transgenic (tg) mice carrying a LSL-MRTF-A cassette and CreERT2. Organoids were seeded on matrices with increasing stiffness and treated for 4 days with 0.25 μM Hydroxy-Tamoxifen (4OHT) or vehicle control. A Representative images of primary murine acini stained for nuclei (DAPI, blue) and actin (phalloidin, green) on matrigel/collagen I matrices 5 days after seeding. Arrowheads indicate protrusions, and asterisks the acinar lumen. B, C Distribution of individual organoid roundness and diameters. D, E Percentage of acini with filled lumen and protrusions. More than 17 acini were analysed per condition. Significance was determined by two-way ANOVA. F Genotyping of 4OHT-treated ( +) and vehicle treated ( − ) tg acini using primer flanking the loxP STOP loxP cassette (LSL, 1500 bp) in comparison to an untreated positive control sample (Ctrl). *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. Error bars or whiskers, SEM. Scale bars, 25 μm
Although the increase of MRTF-A activity could not be quantified due to the minute amount of cell material available, genotyping of tg acini after 4 days in the presence or absence of 4OHT confirmed the removal of the STOP cassette via the ERT2-controlled Cre recombinase (Fig. 5F). Primary murine tg and wt acini were also cultured on matrices of different stiffness for three days to allow recovery and subsequently treated with 4OHT for two days before microscopic examination. Here, staining of HA-labelled MRTF-A provided evidence of expression of the activated MRTF-A transgene in acini after 4OHT treatment (Additional file 1: Fig. S3A). Moreover, even the shorter expression of activated MRTF-A was sufficient to significantly promote luminal filling without causing protrusion formation on soft matrix (Additional file 1: Fig. S3B, C). On the stiff matrix, however, MRTF-A overactivation rather inhibited protrusion formation, in line with a previously proposed uncoupling from mechanotransduction [23].
留言 (0)