
記住我

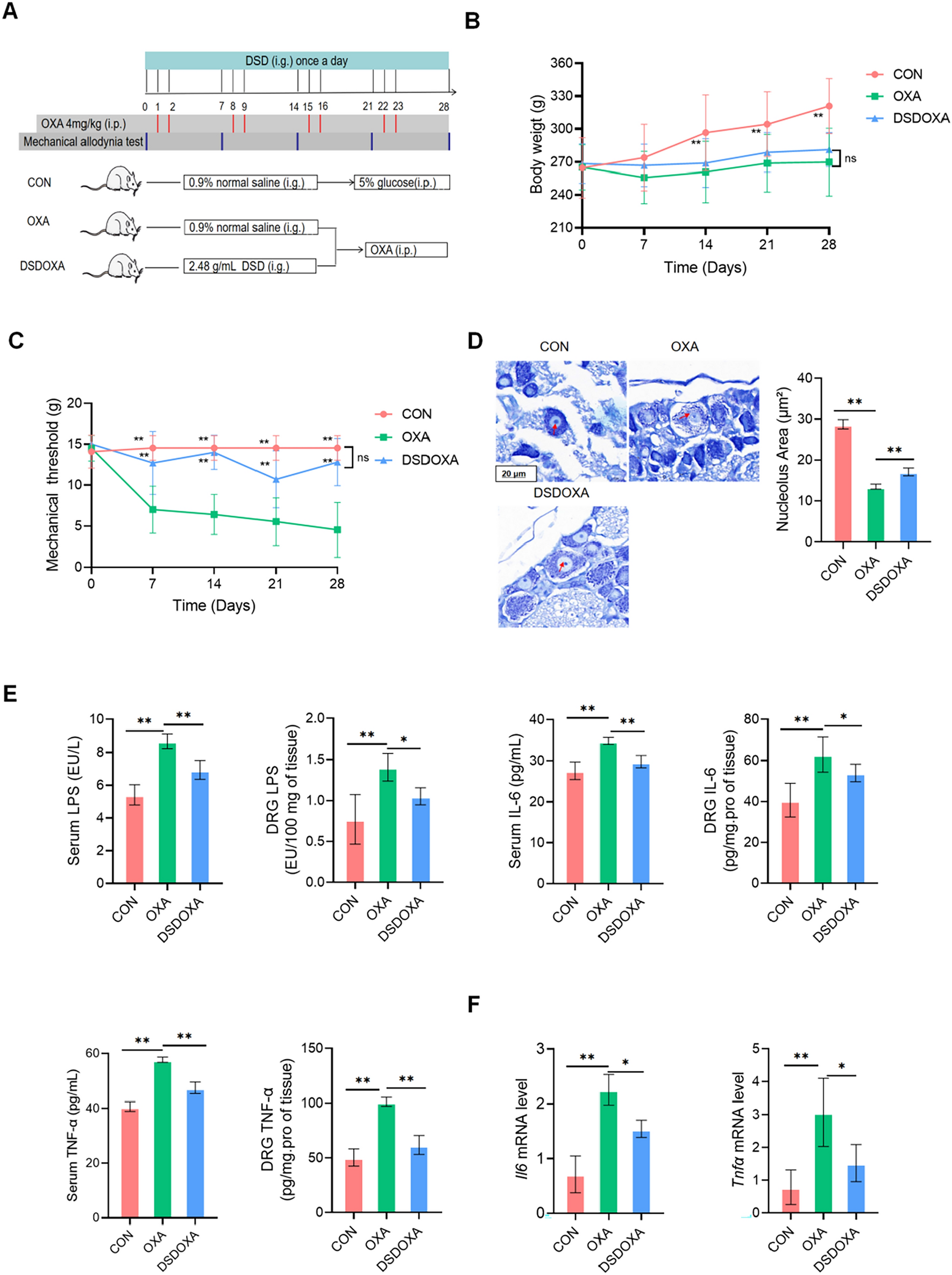


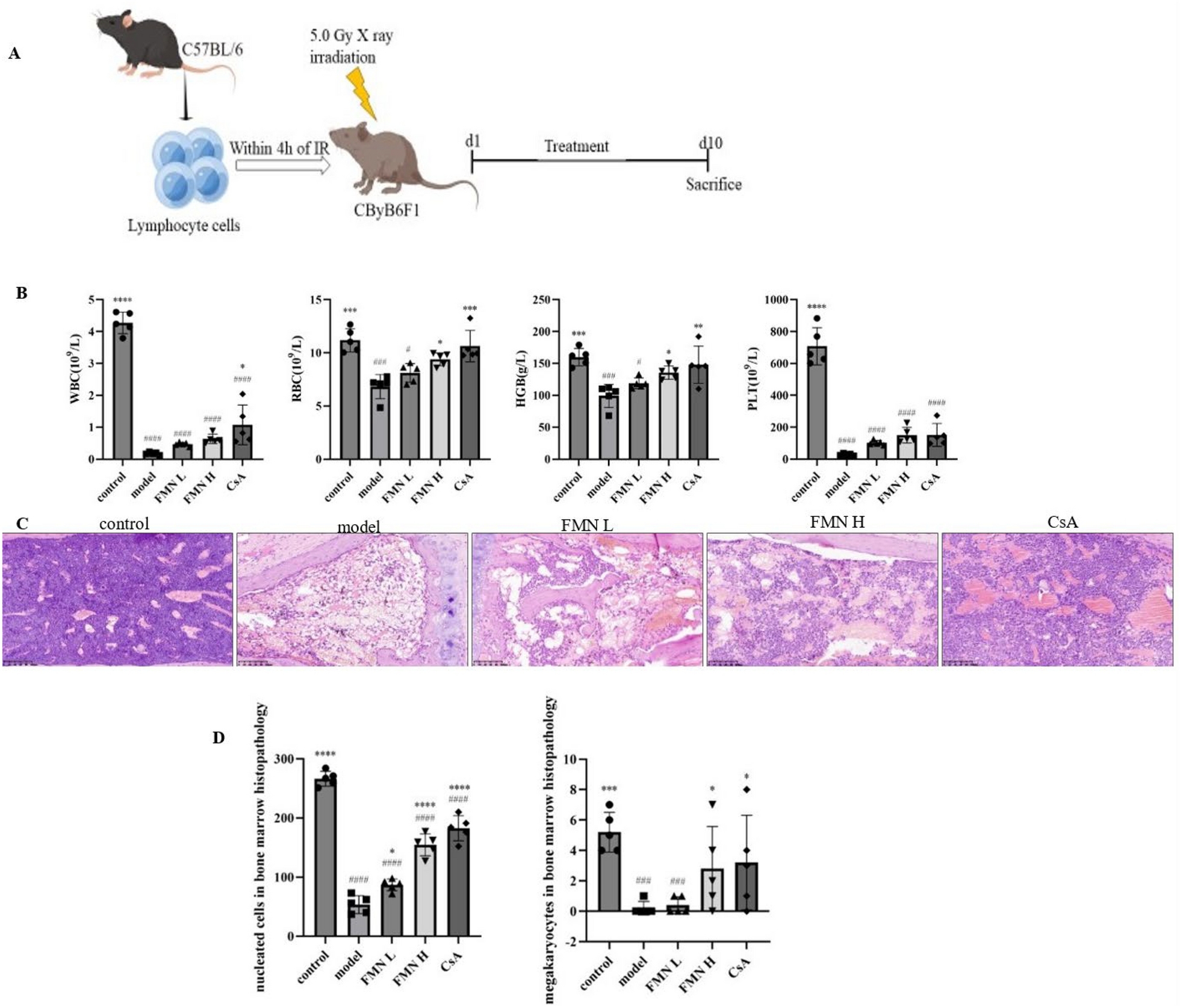
Compared with the NOR group, rats treated with DSS alone exhibited severe diarrhea and rectal bleeding with no weight gain, leading to high DAI during 7 days (Fig. 1A, B). DSS induction also caused remarkably decreased colon length/weight ratio and elevated colonic MPO activity (Fig. 1C, D). Both tonic herbs (GS, AR) and SR slightly increased body weight, alleviated the increases of DAI and MPO, and prevented the decrease of colon length/weight ratio induced by DSS (Fig. 1A–D), while DSS-induced symptoms were aggravated by RB as evidenced by higher DAI mainly attributed to more severe body weight loss, and lower colon length/weight ratio than the UC group. Very interestingly, colonic MPO activity, a sensitive indicator of the influx of inflammatory cells into tissue [34], was also significantly decreased in RB-treated rats.
DSS induction caused the loss of intestinal crypts and goblet cells, severe tissue damage of the epithelial layer, and inflammatory cell infiltration in colon tissues (Fig. 1E, F). The overall tissue damages were attenuated in all herb-treated rats.
Moreover, DSS insult resulted in elevated production of pro-inflammatory (TNF-α, IL-1β, and IL-6) cytokines in serum (Fig. 1G). The levels of anti-inflammatory (IL-4 and IL-10) cytokines were also increased but to less extents. All herbal treatments reversed the increases of the pro-inflammatory cytokines, but further potentiated the elevation of the anti-inflammatory cytokines (Fig. 1G). Similar changes at mRNA levels of these pro- and anti-inflammatory cytokines were observed in the colon tissues. It’s interesting to find that, both tonic herbs also further enhanced the mRNA levels of TGF-β1 which is a multifunctional set of peptides controlling the immune system, while RB reversed its slight increase induced by DSS. SR can suppress the colonic TNF-α elevation to the normal level as efficiently as the tonic herbs (Fig. 1G). Meanwhile, both tonic herbs also mitigated the elevation of the mRNA levels of colonic IFN-γ, MCP-1, iNOS, COX-2, and iCAM-1 induced by DSS.
In general, the two tonic herbs were more effective than the detoxifying SR and/or RB in alleviating clinical colitis symptoms and colonic damages and modulating the aforementioned immune imbalance.
Fig. 1
Therapeutical evaluation of four different medicinal herbs on DSS-induced acute colitis rat model. Dynamic changes of A body weight and B DAI; C Rat colon length vs. weight ratio; D MPO activity in colon; E HE-stained sections of the distal colon, photographs were taken under ×200 magnification. *: mucosa tissue damage; #: inflammatory cells infiltration; →: reduction of goblet cells. F HE scores. Data were expressed as mean ± SD. The significance of differences was determined using one-way ANOVA. *p < 0.05 vs. NOR; #p < 0.05 vs. UC. DSS dextran sulfate sodium, DAI disease activity index, MPO myeloperoxidase; HE Hematoxylin and Eosin
Herbal interventions elicited faster and greater changes in gut microbial structure than urinary metabolic profileCompared with the NOR group, the richness and diversity of the microbial community, as indicated by the estimators Chao1 and Shannon, respectively, were gradually reduced in the UC group (Fig. 2A, B), which were altered by herbs of both types in different manners. Specifically, RB abolished DSS-induced changes in both measurements. SR treatment mitigated the decrease in the richness which was finally restored to the original level, and even ended with significantly higher diversity at the end of the treatment. Similar to SR, AR prevented the decrease of the richness and significantly enhanced the diversity. Interestingly, although GS showed a more potent immune-regulatory effect and the strongest alleviation effect on DSS-induced colitis, it only elevated the diversity at the early stage of DSS insult (day 3) while slightly decreased Chao 1.
The alterations in the gut microbial compositions were further compared at phylum level. More than 99% of the total bacteria were assigned into seven dominant phyla (Fig. 2C), namely Firmicutes, Bacteroidetes, Verrucomicrobia, Proteobacteria, Actinobacteria, Candidatus Saccharibacteria (TM7), and Tenericutes. An increase of Bacteroidetes was observed in the UC group, leading to a decreased Firmicutes/Bacteroidetes (F/B) ratio at the early stage (day 3, Fig. 2D). RB treatment resulted in a decrease of Firmicutes, corresponding to lowered F/B ratios, while SR generally increased the abundance of both Firmicutes and Bacteroidetes and as a consequence maintained the F/B ratio throughout the experimental period. In contrast, GS intervention resulted in markedly decreases in Firmicutes and increases in Bacteriodetes, causing significantly decreased F/B ratios. AR altered the two main phyla in the same manner as GS but to a much lesser extent (Fig. 2C, D). The abundance of Verrucomicrobia in the two groups receiving the detoxifying herbs was much higher than those in the tonic herb groups, which was eliminated by GS treatment. All the four herbs can maintain relatively higher abundance of Proteobacteria which was minor in the UC group, while only RB recovered the abundance of Actinobacteria which was significantly reduced by DSS insult.
At genus level (Fig. 2E), 14 genera showed differential relative abundance among six experimental groups. Specifically, DSS insult resulted in the increases of Enterorhabdus and Flavonifractor as well as decreases of six genera (Prevotella, Rhodococcus, Vibrionimonas, Clostridium XIVa, Clostridium IV, and Lachnospiracea incertae sedis). All four herbs mitigated DSS-induced decreases of Rhodococcus and Vibrionimonas with the tonic herbs more potent than the detoxifying herbs. Both tonic herbs displayed similar effects on five genera with GS more effective, specifically, suppressed DSS-induced elevation of Enterorhabdus, potentiated the increase of Flavonifractor, while reversed the suppression of Rhodococcus and Vibrionimonas and enhanced the levels of Acetatifactor which was unaffected by DSS insult. Additionally, AR potentiated the levels of three extra genera (Clostridium IV, Odoribacter, and Lachnospiracea incertae sedis) which were unaltered by GS treatment. The impact of SR on the gut microbial structure is largely similar to the tonic herbs, except for potentiating DSS-induced elevation of Ruminococcus and restoring the reduced Lachnospiracea incertae sedis. Interestingly, in addition to the reversal of the decreases of Prevotella, Rhodococcus, and Vibrionimonas and the increase of Flavonifractor triggered by DSS, RB intervention also elicited distinct microbial alterations, resulting in marked increases of Parasutterella and Akkermansia which were unaltered in DSS-induced colitis and by other herb treatments. Further hierarchical clustering analysis of gut microbiota structure on day 5 and day 7 revealed a clear separation between the Normal and UC group (Additional file 1: Fig. S1). Agreeing with the genus-level changes, SR and AR were always clustered together, while GS gradually moved away from SR and AR and is clustered into a separate group on day 7. Interestingly, RB displays a distinct pattern in gut microbial structure, making it well separated from all other treatments on both day 5 and day 7.
Fig. 2
Effect of four different medicinal herbs on the gut microbial structure of DSS-induced acute colitis rat model. The microbial community A richness estimator Chao 1 and B diversity estimator Shannon index; C bar chart of the bacterial community composition at phylum level, D the Firmicutes to Bacteroidetes ratio, and E the heatmap of distinct bacteria regulated by UC pathology and tested herbs. Data were expressed as mean ± SD. Significance of differences was determined using one-way ANOVA. *p < 0.05 vs. NOR; #p < 0.05 vs. UC.
Further PCA analysis revealed that the microbial structure at both OTU and genus levels of the UC group markedly shifted away with changes at OTU level essentially separated from the normal status throughout day 3 to day 7 (Fig. 3). In presence of herb intervention, the trajectories of microbial changes drifted away from the colitis group to different extents and in different directions as well. Specifically, the bacterial OTU profile of the RB group on days 5 and 7 was well separated from those of the other three herbal groups in the PC1 and positioned on opposite sides of the normal group (Fig. 3). Although the other three herb-treated groups, including the tonic GS and AR and the detoxifying SR, clustered together and separated from the UC group, they also well separated from the normal group (Fig. 3). The gut microbial structures at genus level were generally less distinguishable than that at OTU level. Only the RB group was still well separated from all other groups, agreeing with the clustering analysis (Additional file 1: Fig. S2). GS moved away from the UC group on day 7, while SR and AR are still partially overlaid with the UC group.
The PCA plot of the urinary metabolome showed high intra-group variations and could not separate different groups from each other (Fig. 3). Supervised PLS-DA plots (Additional file 1: Fig. S2) showed similar patterns to those of the PCA analysis (Fig. 3). The microbiome at OTU level was generally more distinguishable than that at the genus level. The PLS-DA plots revealed high intra-group variations in the urinary metabolome and could not separate different groups from each other on days 3 and 5. Although the colitis group was still slightly overlaid with NOR on day 7, both groups receiving the tonic herbs shifted away from the colitis group and partially merged with NOR.
Fig. 3
Principal component analysis (PCA) of bacterial OTUs, genus and host urinary metabolome on days 3, 5, and 7. Groups in the plot are shown with 95% confidence ellipses
Perturbations of the gut microbiome and host urinary metabolome displayed increasing associationsTo decipher whether there was an overall association between the perturbed gut microbiome and host metabolome during the development of colitis, linear correlation analysis was performed between PC1s of PLS-DA plots of bacterial OTUs and those of host urinary metabolome on days 3, 5, and 7 (Additional file 1: Fig. S3). The linear coefficient R was 0.439, 0.508, and 0.661 on days 3, 5, and 7, respectively, indicating increasing associations between host urinary metabolome and bacterial OTUs. Pairwise Spearman correlation analysis further revealed that the numbers of significant correlations (p < 0.05) between differential genera and metabolites increased dramatically (6, 48 and 69 on days 3, 5 and 7, respectively) (Fig. 4A). The correlations were all positive on day 3, while the numbers of negative correlations increased dramatically from 6 on day 5 to 26 on day 7 (12.5% and 37.7% of total correlations, respectively).
On day 7, 30 distinct metabolites (mainly including nine acyl glycines, nine nucleotide metabolites, four tryptophan metabolites, two steroidal glucuronides, and two acylcarnitines) displayed significant association with 17 differential bacterial genera (including eight from Firmicutes, four from Proteobacteria, two from Actinobacteria, two from Bacteroidetes and one from Verrucomicrobia) with 69 correlations (26 negative and 43 positive) reaching statistical significance (Fig. 4B). Generally, the associations among the acyl glycines (hydroxyhippuric acid (HPA) isomers, phenylbutyrylglycine (PBG), hydroxyphenylacetylglycine (HPAG) and HPAG isomer) and five bacteria at different phylogenetic classification levels, namely the genus Clostridium sensu stricto and the family Clostridiaceae of phylum Firmicutes, the genus Parasutterella and the order Burkholderales of phylum Proteobacteria and the genus Akkermansia (Akkermansia muciniphila) of the phylum Verrucomicrobia, make the biggest contribution to the positive correlation (total 17 positive correlations). The three genera Acetatifactor, Intestinimonas and Clostridium XVIII under phylum Firmicutes made the second contribution with 13 positive correlations among metabolites spanning over all different chemical groups, namely the nucleotides (acetylcytidine and methylcytidine), acyl glycines (HPA), steroidal glucuronides (dehydropregnenolone glucuronide (DPG), estriol glucuronide (EG)), the tryptophan metabolites (N-acetyltryptophan isomer), and others (β-alanine and riboflavin). For the negative correlations, four bacteria, including the two genera Lactobacillus and Flavonifractor from Firmicutes, the family Coriobacteriaceae from Actinobacteria and the order Rhodospirillales from Proteobacteria, are the main contributors and established 17 significant correlations with many metabolites in the nucleotide group (trimethylguanosine (TMG), methyladenosine, succinyladenosine, pterin, acetylcytidine), acyl glycines (hydroxyphenylpropionylglycine (HPPG) and two isomers), and others (l-histidine and creatinine). It’s interesting to note that the association analysis revealed the distinct relationships of different groups of bacteria to one specific chemical type, for instances, the three genera Acetatifactor, Intestinimonas and Clostridium XVIII were positively associated to most nucleotide metabolites, while the six bacteria (the three genera Lactobacillus, Flavonifractor, and Clostridium sensu stricto, the two families Clostridiaceae 1 and Coriobacteriaceae, the order Rhodospirillales) displayed negative correlations. Notably, the same group of bacteria can correlate distinctly (e.g., HPA and its two isomers, N-acetyltryptophan and its isomer) or in similar profiles (e.g., the steroidal glucuronides DPG and EG, the acylcarnitines acylcarnitine (C6:1) and phenylacetyl-carnitine (PAC); deoxylnosine and deoxylinosine isomer) with chemical analogs.
Further inspection of the results found that the tonic herbs, especially GS, made a higher contribution to the positive correlations between genus Rhodococcus and the two metabolites β-alanine and guanine (Additional file 1: Fig. S4A). RB is the sole contributor to the strong correlations between the acyl glycines HPAG and PBG and the genera Akkermansia and Parasutterella (Additional file 1: Fig. S4B), while SR is the one accounting for the correlation between the genus Acetatifactor and both steroidal glucuronides (Additional file 1: Fig. S4C).
Fig. 4
Gut microbiome and host urinary metabolome displayed increasing associations. A The number of significant correlations between distinct genera and metabolites on days 3, 5 and 7. B heatmap of significant correlations between differential genera and metabolites on day 7. #: linear correlation coefficient R > 0.5 and p < 0.05. TMG trimethylguanosine, HPPG hydroxyphenylpropionylglycine, HPA hydroxyhippuric acid, PBG phenylbutyrylglycine, HPAG hydroxyphenylacetylglycine, DPG dehydropregnenolone glucuronide, EG estriol-glucuronide, ICAG 3-indole carboxylic acid glucuronide, 5-HTP 5-hydroxytryptophan, PAC phenylacetyl-carnitine
RB and SR triggered significant alterations in steroidal glucuronides and acylglycines, respectivelyThe differential metabolites induced by DSS and those by each herbal intervention were further analyzed (Fig. 5). DSS insult elicited gradual decreases of guanine, deoxyinosine, N-acetyltryptophan and isomer, PAC and acylcarnitine (C6:1) throughout the experimental period. In contrast, pterin, TMG, HPPG, 5-hydroxy-tryptophan (5-HTP), and EG were significantly increased in DSS-induced colitis, peaking at day 5 while maintaining or decreasing on day 7. The effects of the two tonic herbs on the metabolic shifts are largely similar: both could abolish (guanine, N-acetyltryptophan) or mitigate (HPPG, 5-HTP, PAC, acylcarnitine (C6:1)) most aforementioned changes. Although the detoxifying herb SR shared some common features with RB, such as potentiating DSS-induced decrease of deoxyinosine, its actions on DSS-induced changes more resemble those of the two tonic herbs. Particularly, both tonic herbs and SR increased the level of alanine, which was unchanged after DSS insult or RB intervention; both AR and SR caused significant increases in HPA, a microbial aromatic acid metabolite, which was not observed in DSS induction. It is interesting to find that the two detoxifying herbs elicited distinct metabolic alterations. Specifically, RB abolished DSS-induced elevation of the acyl glycine HPPG, did not alter the level of HPA which was elevated by SR and AR, but induced significant and unique increases of PBG, HPAG and its isomer. RB also significantly decreased the level of 3-indole carboxylic acid glucuronide (ICAG), a glucuronide of indole pathway of tryptophan metabolism, which was not affected by DSS insult or any other herbal intervention. In contrast, SR treatment resulted in the elevation of EG and DPG, although the level of ICAG was unaltered. The changes of the two steroidal glucuronides showed a high correlation with that of genus Acetatifactor with sole contribution by SR (R values 0.916 and 0.977) (Additional file 1: Fig. S4C).
Fig. 5
Relative abundance of distinct metabolites. A nucleotide metabolites: guanine, deoxyinosine, pterin, trimethylguanosine (TMG); B acyl glycines: hydroxyphenylpropionylglycine (HPPG), hydroxyhippuric acid (HPA), hydroxyphenylacetylglycine (HPAG), phenylbutyrylglycine (PBG); C tryptophan metabolites: N-acetyltryptophan and isomer, hydroxytryptamine (5-HTP), 3-indole carboxylic acid glucuronide (ICAG); D steroidal glucuronide conjugates: dehydropregnenolone glucuronide (DPG) and estriol-glucuronide (EG), E carnitines: phenylacetyl-carnitine (PAC) and acylcarnitine (C6:1), and F β-alanine in different groups. Data were expressed as mean ± SD. *p < 0.05
Crosstalk among gut microbiome-metabolome-UC axisTo further assess whether there exist associations among gut microbiome, host metabolome and therapeutical effect of herbs, an interaction network was constructed with data from 3 dimensions: 17 differential gut microbial genera, 30 distinct metabolites and 9 phenotypic measurements (Fig. 6). It is interesting to find that the phenotype data were integrated into the network. In particular, two anti-inflammation cytokines (IL-4 and IL-10) showed similar correlation profiles. They simultaneously showed strong positive correlation with three genera i.e., Acetatifactor, Clostridium, and Intestinimonas, as well as negative correlation with the family Coriobacteriaceae. Meanwhile, they also positively correlated with two nucleotides (methyladeosine and acetylcytidine), l-histidine and β-alanine, while negatively with ICAG. Three pro-inflammatory cytokines (IL-1β, TNF-α and IL-6) and MPO displayed most negative associations with metabolites guanine, PAC, acylcarnitine (C6:1), and bacteria including genera Vibrioniomons and Rhodcoccus, family Brucellaceae. Other UC indices, including DAI, HE scores and MPO, negatively related to guanine, while the ratio of colon length to weight showed a positive correlation with guanine.
Fig. 6
Correlation network among discriminative gut microbial taxa, differential metabolites and UC indices (Spearman coefficient > 0.5 and p < 0.05). The red line represents the positive correlation, and the blue line represents the negative correlation. The stronger the interaction, the thicker and darker a line is. The node size represents degree. The more correlations with other elements, the bigger a node is. TMG trimethylguanosine, HPPG hydroxyphenylpropionylglycine, HPA hydroxyhippuric acid, PBG phenylbutyrylglycine, HPAG hydroxyphenylacetylglycine, DPG dehydropregnenolone glucuronide, EG estriol-glucuronide, ICAG 3-indole carboxylic acid glucuronide, 5-HTP 5-hydroxytryptophan, PAC phenylacetyl-carnitine, DAI disease activity index, MPO myeloperoxidase, HE Hematoxylin and Eosin
留言 (0)