
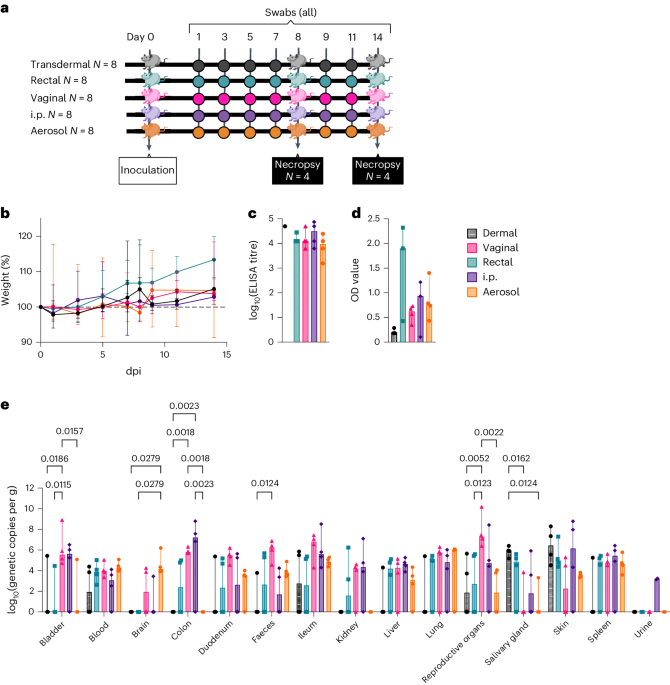
記住我

The coronavirus disease 2019 (COVID-19) pandemic started in December 2019 and has caused 591 million cases and claimed 6.4 million lives as of 19 August 2022. SARS-CoV-2 (ref. 1), which causes COVID-19, is a member of the subgenus Sarbecovirus, as is SARS-CoV-1, which caused the SARS epidemic 19 years ago2. Bats are reservoirs for SARS-related coronaviruses3,4. Multiple sarbecoviruses have been detected in bats5,6,7,8 and, more recently, in pangolins9. SARS-CoV-2 variants of concern (VOC) have emerged since late 2020, probably in response to immune responses in the human population10. So far, five major VOCs have been recognized by the WHO (Alpha, Beta, Gamma, Delta and Omicron). SARS-CoV-2 VOCs have either developed resistance/escape to neutralizing antibodies11,12,13,14,15,16 or acquired mutations that have increased transmission or pathogenicity17,18. SARS-CoV-2 Omicron, which contains 32 amino acid mutations in the spike, was first detected in South Africa and Botswana and has rapidly spread to many countries19,20,21,22.
We applied our SARS-CoV-2 surrogate virus neutralization test (sVNT) platform23,24 which can detect total immunodominant neutralizing antibodies targeting the viral spike (S), and differentiated antibody responses to several human and animal sarbecoviruses in a 20-plex assay that included 15 receptor binding domains (RBDs) of the SARS-CoV-2 clade (clade-1b) viruses and 5 RBDs of the SARS-CoV-1 clade (clade-1a) viruses. Clade-1b viruses included the ancestral SARS-CoV-2 virus (Wuhan-hu-1), variants of concern or interest (Alpha, Delta, Beta, Gamma, Delta plus, Lambda, Mu, Omicron BA.1 and Omicron BA.2) and animal sarbecoviruses (BANAL-52, BANAL-236, GD-1, RaTG13 and GX-P5L). For clade-1a, we included human SARS-CoV-1 and bat sarbecoviruses (Rs2018B, LYRa11, RsSHC014 and WIV1). The phylogenetic relatedness and the amino acid sequence differences of these RBDs and spikes are shown in Extended Data Fig. 1a–d.
All RBDs bind to human ACE2 in a dose-dependent manner in the multiplex Luminex system (Extended Data Fig. 2a). We first tested the 20-plex sVNT using the WHO international standard 20/136 (ref. 25) and found that the neutralization titres were reduced from ancestral SARS-CoV-2 in the following order for the human SARS-CoV-2 variants (from least to most): Alpha, Delta, Delta plus, Lambda, Gamma, Beta, Mu, Omicron BA.2 and Omicron BA.1 (Extended Data Fig. 2b). The calibration of WHO international standards using SARS-CoV-2 ancestral multiplex sVNT was modelled (Extended Data Fig. 2c). From a well-defined panel of 120 sera with varying levels of neutralizing antibodies (NAbs), the data showed a good correlation between sVNT and pVNT (pseudovirus-based VNT) with R2 of 0.83 and 0.73 for the ancestral and Omicron BA.1 virus, respectively, and with good correlation with Omicron BA.1 plaque reduction neutralization test (PRNT) with R2 of 0.79 (Extended Data Fig. 2d–f).
With our 20-plex sVNT platform, we observed significantly more neutralization escape by Omicron BA.1 and BA.2 than by any of the other variants, and this was consistent for all 20 serum panels tested in this study (Fig. 1a) (extent of neutralization in decreasing order: Ancestral > Alpha > Delta > Delta plus > Lambda > Gamma > Beta > Mu > Omicron BA.2 > to lowest Omicron BA.1). Vaccinated individuals who received a third dose of BNT162b2, mRNA-1273 or AZD1222, but not inactivated vaccines, increased overall NAb titre to all viruses. Even in these individuals, a significant reduction in NAb level to Omicron was evident in all serum panels, including those with hybrid immunity (Fig. 1a). Even in those with Omicron-breakthrough infections, the NAb titre to Omicron remained lower than for other SARS-CoV-2 variants (Fig. 1a).
Fig. 1: sVNT and pVNT assay.
a, A heat map of sVNT GMT50 of 20 serum panels derived from convalescent, vaccinated, boosted and hybrid individuals for immunity against 20 sarbecoviruses using multiplex sVNT. b, Illustration of the clade-1b (SARS-CoV-2, RaTG13 and GX-P5L) and clade-2 (RmYN02 and SL-ZC45) RBDs used in rabbit immunization. The receptor binding motif is highlighted in blue. The percentage of amino acid sequence identity to SARS-CoV-2 RBD is indicated. c,d, The NAb level of the hyperimmune rabbit sera to different sarbecoviruses were determined by 20-plex sVNT (c) and pVNT (d) of different sarbecovirus spike proteins indicated above the panels.
Next, we tested a panel of well-defined hyperimmune rabbit sera raised against different recombinant RBD proteins of clade-1b and clade-2 sarbecoviruses, including human SARS-CoV-2, bat CoV RaTG13, pangolin CoV GX-P5L (clade-1b) and two non-ACE2-binding RBDs from bat CoV RmYN02 and bat CoV SL-ZC45 (clade-2)5. Both RmYN02 and SL-ZC45 RBD have major deletions in the receptor binding motif and shared 62.8% and 65.6% amino acid sequence identity to SARS-CoV-2, respectively (Fig. 1b). Consistent with our data from human sera, Omicron BA.1 and BA.2 are the only SARS-CoV-2 viruses that escape NAbs from all rabbit sera in the panel. Sera raised against RaTG13 and GX-P5L RBDs had 6.4-fold and 3.8-fold NAb titre reduction, respectively, to SARS-CoV-2, but Omicron BA.1/BA.2 showed an almost complete NAb escape with a 58-fold/38-fold and 18-fold/16-fold reduction, respectively, against the RaTG13 and GX-P5L hyperimmune sera (Fig. 1c). This was confirmed by pVNT with a 135-fold reduction of neutralizing activity to Omicron BA.1 in GX-P5L hyperimmune sera (Fig. 1d). The hyperimmune sera raised against the non-ACE2-binding RBDs of RmYN02 and SL-ZC45 had the lowest cross-NAbs to SARS-CoV-2 (Fig. 1c,d). As a negative control, no NAb was detected in the rabbit hyperimmune sera raised against a non-sarbecovirus bat CoV HKU1 RBD (Fig. 1c,d).
When comparing NAb escape between human VOCs and animal sarbecoviruses, we observed that Omicron had greater neutralization escape than was observed in animal sarbecovirus RaTG13 and GX-P5L using multiplex sVNT, although there were only 15–16 amino acid mutations in Omicron RBDs compared with 22 and 30 mutations in RaTG13 and GX-P5L, respectively (Extended Data Figs. 1a,b and 2a,b). In addition to Omicron, we further observed that VOCs Beta and Mu had higher NAb escaping ability than their most genetically related animal sarbecoviruses bat BANAL-52 and pangolin GD-1 (Fig. 2a,b), even though these animal viruses are phylogenetically more distant from SARS-CoV-2 (Extended Data Fig. 1a,c). Using pVNT, we further confirmed that the SARS-CoV-2 Beta variant is more potent in NAb evasion than animal sarbecoviruses BANAL-52 and GD-1 despite the animal sarbecoviruses containing 20 and 127 mutations (Extended Data Figs. 1d and 3a and Fig. 2c,d), respectively, in the spike protein. SARS-CoV-2 Omicron BA.1 and BA.2 have comparable pVNT 50% neutralization titre (NT50) with GX-P5L (Fig. 2c).
Fig. 2: The effect of RBD and spike mutations on NAb escape.
a, Effect of RBD mutations examined by multiplex sVNT is shown by neutralization titres (n = 75) for SARS-CoV-2 ancestral, Delta, Beta, Mu, Omicron BA.1, Omicron BA.2, Bat CoV BANAL-52, RaTG13, Pangolin CoV GD-1, GX-P5L and SARS-CoV-1. b, The effect of the number of RBD mutations on NAb escape (n = 89). The effect of all mutations in the full spike protein was measured using pVNT and is shown in c. c, Neutralization titres (n = 75) for SARS-CoV-2 ancestral, Delta, Beta, Omicron BA.1, Omicron BA.2, Bat CoV BANAL-52, Pangolin CoV GD-1, GX-P5L and SARS-CoV-1. d, The effect of the number of spike mutations on NAb escape (n = 69). Statistical analysis in a and c was performed using two-sided Wilcoxon rank sum test in GraphPad Prism 8 (in a: Beta versus GD-1 P < 0.0001, Beta versus BANAL-52 P < 0.0001, Mu versus GD-1 P < 0.0001, Mu versus BANAL-52 P < 0.0001, Omicron BA.1 versus RaTG13 P < 0.0001, Omicron BA.1 versus GX-P5L P < 0.0001, Omicron BA.1 versus Omicron BA.2 P < 0.0001, Omicron BA.2 versus RaTG13 P < 0.0001, and Omicron BA.2 versus GX-P5L P < 0.0001; in c: Beta versus GD-1 P < 0.0001, Beta versus BANAL-52 P < 0.0001, Omicron BA.1 versus GX-P5L P = 0.0035, Omicron BA.1 versus GD-1 P < 0.0001, Omicron BA.1 versus Omicron BA.2 P = 0.0063). Each dot denotes the NT50 value of a sample, while the box shows the interquartile range with median at the centre, and the whiskers represent the maximum and minimum. Linear regression analysis in b and d was performed using GraphPad Prism 8. Error bars in b and d indicate standard deviation of the geometric mean.
Furthermore, SARS-CoV-1 survivors who received two doses of BNT162b2 vaccine displayed broad NAbs against all known sarbecoviruses before the emergence of Omicron. However, sera from this panel also had some degree of NAb escape to Omicron (Fig. 1a). We showed that amino acid mutation G339D, S371L/F, S373P, S375F, D406N and R408S located at the conserved regions are unique to Omicron but are not present in animal sarbecoviruses or SARS-CoV-1 (Extended Data Fig. 3b). Interestingly, S371, S373, S375, D406 and R408 are located in the pan-sarbecoviruses neutralizing epitopes of class VI antibodies26,27. In our mutagenesis data, we observed that mutations at the receptor binding motif (amino acids 417–508) had little-to-no impact on escape from NAbs derived from BNT162b2-vaccinated SARS-CoV-1 survivors, but sera from this panel showed 2.9-fold reduction in geometric mean neutralizing titer 50% (GMT50) against ancestral RBD in RBD with S371L, S373P and S375F mutations (Extended Data Fig. 3c).
Finally, data from antigenic cartography confirmed that Omicron VOCs are antigenically more distinct from the ancestral SARS-CoV-2 than animal sarbecoviruses (Extended Data Fig. 4a). Consistent with our previous findings, we found that the overall antigenic breadth of BNT162b2-vaccinated SARS-CoV-1 survivors is significantly broader than that of BNT162b2-vaccinated infection-naïve individuals (Extended Data Fig. 4b). More recently, Omicron BA.5, which carries three additional mutations compared with BA.2 in the RBD (L452R, F486V and R493Q), emerged and is overtaking Omicron BA.2 in South Africa and the United States. Using the same serum panels, we demonstrated even more potent NAb escape of mRNA vaccine-induced neutralizing antibodies by Omicron subvariants BA.2.11 and BA.5 with the additional L452R mutation and L452R/F486V/R493Q mutations, respectively (Extended Data Fig. 5).
Combining our sVNT detection platform with a collection of 20 different human serum panels, we have shown that the degree of NAb escape in VOCs is greater than that of distantly related animal sarbecoviruses. We propose that the SARS-CoV-2 Omicron variant emerged under immune selection imposed during 2 years of virus transmission in humans. On the other hand, the sarbecoviruses in animals, mainly in bats, seem to be involving at a slower rate probably due to two possibilities. First, ACE2 may not be the only or main entry receptor in bats. Second, antibody-mediated immune responses are not as strong in bats as in humans28.
留言 (0)