
記住我


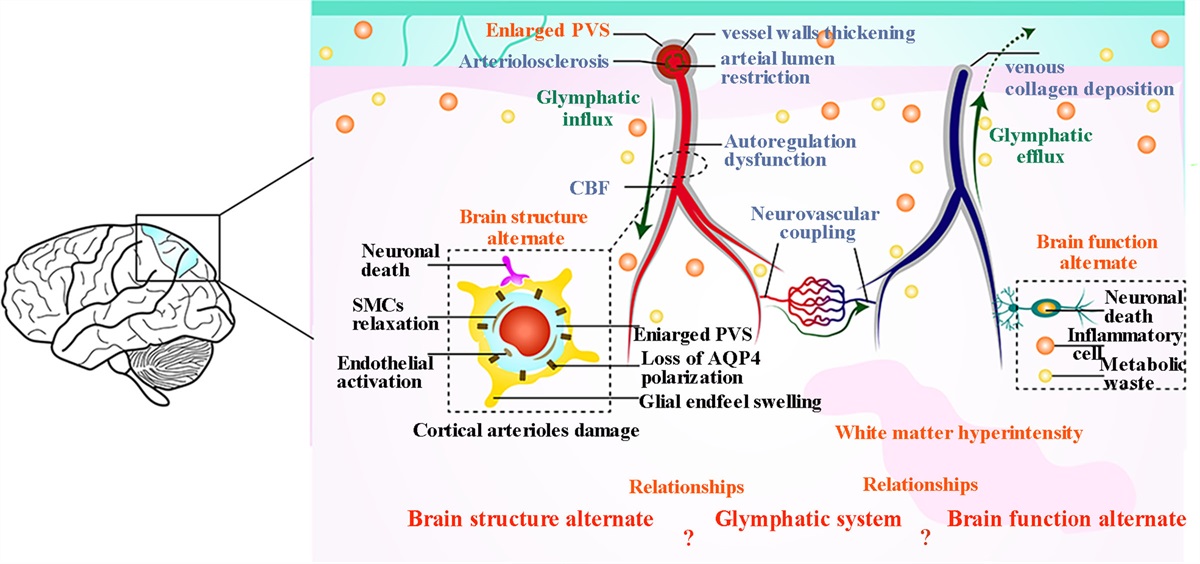
The premotor cortex is traditionally known to be involved in the preparation and execution of movements [1–4]. On the other hand, while some evidence exists for its involvement in visuomotor association learning [5,6], not much is known about its involvement in other types of motor learning or motor learning in general. More recently, the dorsal premotor cortex (PMd) is shown to have modulatory interactions with the cerebellum during visuomotor adaptation such that inhibition of PMd by the cerebellum becomes weaker, which in turn increases excitatory modulation of the cerebellum by PMd [7]. This raises a possibility that PMd is also involved in sensory error-based motor adaptation, a process in which the cerebellum is heavily involved [8,9].
If motor adaptation entails changes in the activity of PMd and its interaction with the cerebellum, it is possible that neuromodulation of PMd can influence adaptation behavior. One recent study has confirmed this in monkeys, showing attenuated trial-by-trial visuomotor adaptation by intracortical micro-stimulation to disrupt PMd at go-signal right before experiencing a sensory error [10]. However, whether motor adaptation can be modulated by targeting PMd in humans has not been tested. Also, since invasive stimulation cannot be readily used by humans, it is important to achieve modulation by a safer and more convenient stimulation method.
The purpose of this study is to test this by transcranial magnetic stimulation (TMS), a noninvasive method used to modulate motor behavior [11]. The task is modified from the original task [10] for humans while keeping the stimulation timing the same, applying TMS just before experiencing a sensory error. Given the substantial overlapping between monkeys and humans in brain structures and functions, we predicted that TMS can modulate human motor adaptation.
We also examine if modulation is region-specific by comparing stimulation on PMd with the ventral premotor cortex (PMv), a region whose roles are known to be different from PMd in motor behavior [12–14]. In particular, PMd plays a role in selecting and preparing movements, especially ones associated with visual cues, whereas PMv does in preparing and guiding movements in extrinsic visual space. Since the original study observed attenuated disruption of adaptation by stimulation on the motor cortex compared to PMd, we predict modulation of motor adaptation by TMS is also region-specific, showing attenuated/nullified effects when PMv is stimulated.
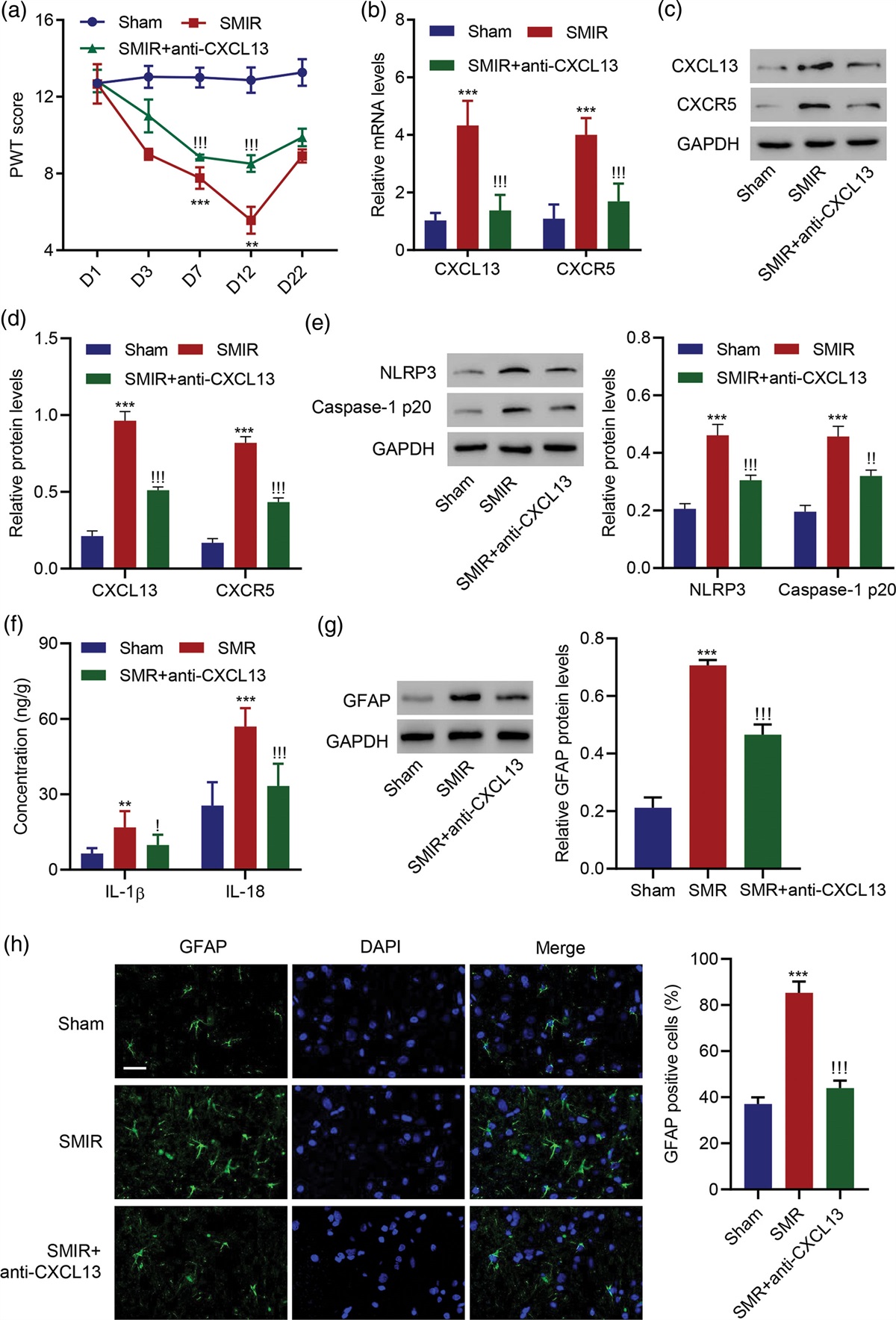
Methods Participants and experimental apparatusTwenty-six right-handed participants [aged 18–28 years (µ = 22.2), 17 males] without known neurological/motor disorders gave written informed consent before participating in the study approved by the Institutional Review Board at the University of Tsukuba. All the participants had a laterality quotient score of +40 or higher on the Edinburgh Handedness Inventory. The participants were recruited across departments without specific motor skill requirements, representing the university’s general population. They were paid 2150JPY for their participation. We terminated an experiment session before the motor task for one participant who reported disrupting facial muscle twitch when PMd/v was stimulated. The remaining 25 participants were included in the data analyses.
A typical apparatus for motor adaptation was used. The participants manipulated a robot manipulandum handle that moved in the horizontal 2D plane with the right hand. The participant’s arm and the handle were occluded by a flat mirror. A computer monitor above the mirror displayed visual stimuli that were reflected on the mirror. The vertical gaps between the monitor, mirror, and handle were matched so that the visual stimuli appeared at the same height as the handle to the participants. Magstim 2002 and a figure-eight coil were used. Brainsight version 2.3.12 tracked the coil position, the participant’s head, and the target locations during the experiment.
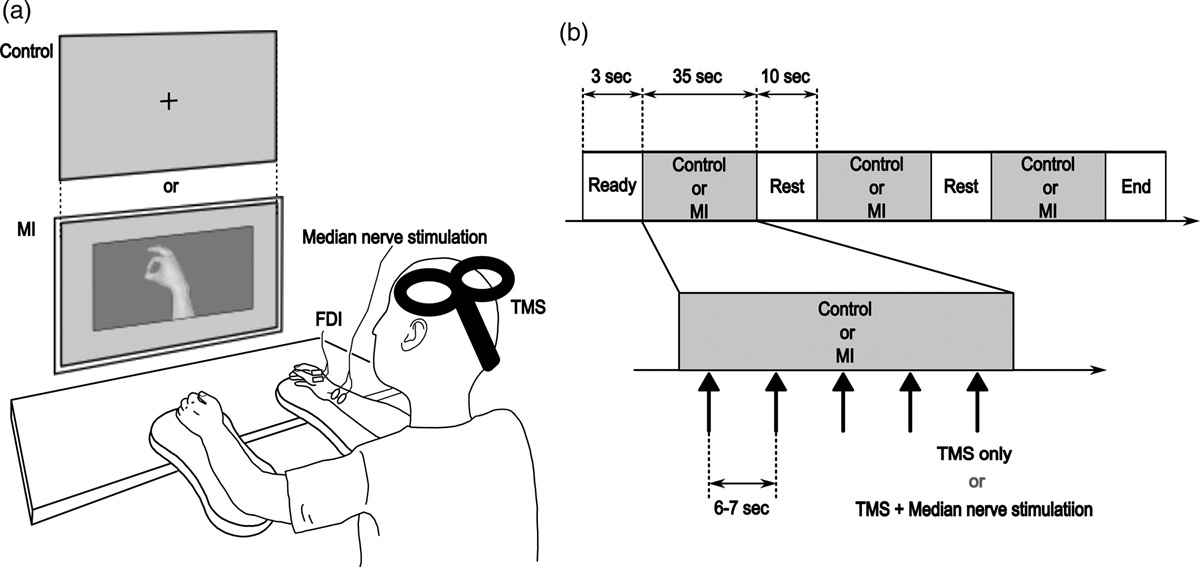
Experimental paradigm Transcranial magnetic stimulation procedureThe locations and the intensity of TMS were determined for each participant. All stimulations were delivered on the left side of the head. We used resting motor threshold, defined as the intensity inducing motor evoked potential (MEP) of more than 50 µV 50% of times [15] in the first dorsal interosseous in the right hand when stimulating a ‘hotspot’, defined as the location where MEP was induced with the lowest intensity. The measured motor thresholds in the present study were between 40 and 60% intensity and 49.8% on average. PMd and PMv were defined as 2 cm anterior and 1 cm medial and lateral to the hotspot, respectively (Fig. 1a) [16,17]. To avoid muscle twitch in the arm, supra-threshold intensity (90% of motor threshold) was used. Coil orientation was pointing 45° in the para-sagittal plane (i.e. coil top pointing anterior-dorsal direction) [18].
 Fig. 1: Task design and schedule. (a) Locations of stimulation and task environments. The locations of the dorsal/ventral premotor cortex (PMd/PMv) are defined relative to the hotspot of the first dorsal interosseous (FDI). Sham is shot in the air near the left ear. Horizontal 2D task space is located in front of the participants, where they made rapid arm-reaching movement from the start position to the target. (b) Motor adaptation paradigm. Visual rotation is applied to the online cursor feedback about hand movement in the Rotation trial, inducing adaptation. The target shifts to the cursor direction so that the cursor always hits the target, clamping the task performance feedback [20]. (c) Task schedule. One block consists of three sets of 10 Null trials with veridical cursor for washout and 10 Rotation trials for adaptation. There is a short break at the end, and the participants repeat 12 blocks. One TMS location is selected per set and once per block. (d) Time flow of one trial, designed based on the previous study [10].Motor task and schedule
Fig. 1: Task design and schedule. (a) Locations of stimulation and task environments. The locations of the dorsal/ventral premotor cortex (PMd/PMv) are defined relative to the hotspot of the first dorsal interosseous (FDI). Sham is shot in the air near the left ear. Horizontal 2D task space is located in front of the participants, where they made rapid arm-reaching movement from the start position to the target. (b) Motor adaptation paradigm. Visual rotation is applied to the online cursor feedback about hand movement in the Rotation trial, inducing adaptation. The target shifts to the cursor direction so that the cursor always hits the target, clamping the task performance feedback [20]. (c) Task schedule. One block consists of three sets of 10 Null trials with veridical cursor for washout and 10 Rotation trials for adaptation. There is a short break at the end, and the participants repeat 12 blocks. One TMS location is selected per set and once per block. (d) Time flow of one trial, designed based on the previous study [10].Motor task and schedule
The ability of motor adaptation was assessed in a visuomotor rotation task. The participants were asked to make a rapid reaching movement to cross the visual target. The online cursor feedback was provided during movement to show the position of the occluded hand (Fig. 1a). In the Rotation trials (Fig. 1b), the cursor was rotated by +5° with respect to the direction of the visual target. The counter-clockwise (CCW) direction was defined as the positive direction. The participants were expected to adapt to the rotation by recalibration of visuomotor mapping, changing their movement in the opposite direction to the rotation over trials [19]. Since performance feedback (difference between the cursor and the target) was known to influence adaptation [20], the target disappeared before the participants initiated movement. The target reappeared before crossing, but its position was shifted to align with the direction of the cursor movement so that the cursor always hit the target, clamping the performance feedback across trials. TMS was applied before they initiated the movement. Since TMS might affect movement itself [21] in addition to the adaptation ability, adaptation behavior was assessed in the next trial, in which reaching was performed without TMS or the cursor feedback.
The task consisted of 12 blocks, each of which consisted of three sets of washout and adaptation (Fig. 1c). One set consisted of 10 ‘Null’ trials with veridical cursor feedback serving as washout of the previous adaptation, which was followed by 10 Rotation trials (five pairs of a trial with TMS and the cursor, followed by a trial without them). There was a 30-s break at the end of each block. TMS was delivered in Rotation trials with the cursor, and, to avoid TMS from always coinciding with rotation, TMS was also delivered in every odd Null trial. There were three TMS locations for the three sets (one location per set): PMd, PMv, and Sham (TMS pulse delivered in the air, Fig. 1a). The order of the TMS locations was randomized across the blocks such that each of the six possible orders (PMd → PMv → Sham, PMd → Sham → PMv, and so on) was randomly selected twice in the 12 blocks. Also, to avoid use-dependent learning [22] due to repeatedly adapting to the same direction, the rotation of the cursor was flipped to the negative [clockwise (CW)] direction in the even blocks.
Trial flowFigure 1d shows the time flow of a single trial that was based on the original study [10]. In each trial, the manipulandum brought the hand to the 9-mm circular start area in front of the participants, and the 5-mm circle was shown as the cursor feedback. After 500 ms, a 5-mm circular target appeared at 10 cm from the start. The target was randomly selected from 0° (right in front), ±6°, and ±12° to avoid use-dependent learning [22]. After an additional wait, the target flashed as a go-signal and disappeared, at which TMS was delivered (only in the trials described above). A rapid shooting movement was made to the target. A target reappeared when the hand reached at 8 cm, and a trial ended once the hand exceeded 10 cm. A warning message was shown if the movement duration was not within 150–250 ms. A trial was aborted and retried if the movement was not initiated within 100–450 ms after go-signal or when the movement duration exceeded 500 ms.
Session scheduleThe participants received task instruction and briefly practiced each trial type to familiarize themselves with the task. Then, the TMS procedure was performed. After that, they performed the motor task. A break between the sixth block and the seventh block was extended from 30 s to 3 min.
Data analysisAdaptation behavior is measured as changes in the reach direction over trials, and how stimulation of PMd modulates the behavior is examined by comparing it to the control conditions (PMv and Sham). To reduce the trial-by-trial variability (noise), individual means were first taken across blocks for each condition. The reach direction data for negative rotation (in the even blocks) were multiplied by −1 to match the direction of adaptation before the means were computed. For the hand trajectories (Fig. 2a), means were taken for Null and Rotation trials without TMS. For the profiles of changes in the hand direction (Fig. 2b), means were taken for the last Null trial and each Rotation trial. For the average hand directions across trials (Fig. 2c), five Rotation trials without the cursor/TMS were separated into halves (the third trial was not included, Fig. 2b), and the mean direction (μh) was taken for each half. Group means and SEM were calculated from these individual means.
 Fig.2:
Fig.2: Modulation of learning behavior by TMS. (a) The hand trajectories of the individuals (gray) and the conditions (colored) across Null and Rotation trials. To assess adaptation behavior without potential influence of TMS on movement itself, only those trials without TMS are included. (b) The profiles of changes in the hand direction over trials. (c) The mean hand directions in Rotation trials, separated by first and second halves. Dots/lines and error bars represent means and SEM.
We assessed the modulatory effects of TMS on PMd while considering effects from other factors, including the progress of adaptation by time, location-independent effects of TMS (TMSLI), and individual differences. To do so, μh was analyzed by a linear mixed effect model [23] with TMS [(PMv and Sham) vs. PMd], Half (first vs. second), and TMSLI [Sham vs. (PMd and PMv)] as the fixed effects and Participant as the random intercept effect, as follows:
μh = TMS + Half + TMSLI + Participant + ε
where ε represented a residual. TMS represented the main factor of interest: whether or not adaptation measured as μh was modulated by TMS on PMd. Half represented the progress of adaptation, where μh in the second half was expected to be larger than the first. TMSLI represented a potential artifact of delivering TMS regardless of location. Finally, Participant represents interparticipant differences in the baseline level of adaptation. Dummy coding was used to represent each fixed factor as following (TMS: Sham/PMv = 0, PMd = 1; Half: 1st = 0, 2nd = 1; TMSLI: Sham = 0, PMd/v = 1). All tests were performed as two-sided tests, and results with P < 0.05 were considered statistically significant. The 95% confidence interval (CI) of each fixed effect was reported. All the data analyses were performed on R version 4.0.2 with lme4 [23] and lmerTest [24] packages.
ResultsFigure 2a shows the average hand trajectories in Null and Rotation trials without TMS. Each TMS condition showed similar patterns in which the hand moved straight to the target in the Null trials but showed slight deviation in the CW direction in the Rotation trials (i.e. adaptation compensating the CCW rotation). Figure 2b shows the profiles of changes in the hand direction over trials, exhibiting increasing hand directions like a typical learning curve. The hand direction in PMd appeared slightly larger than the other conditions, which became clearer in the average hand direction in the Rotation trials (μh), especially for the second half (Fig. 2c).
The linear mixed effect model analysis was performed on μh to statistically examine the effects of PMd stimulation on the adaptation behavior as well as other factors (see Methods section for detail). The analysis demonstrated that μh was larger in the second half than the first half (µ = 1.27; 95% CI, 1.02–1.51; t = 10.02; P < 10−16, Cohen’s d = 1.79), confirming more adaptation over time. It also demonstrated that μh was larger when PMd was stimulated compared with PMv and Sham (µ = 0.39; 95% CI, 0.08–0.69; t = 2.49; P = 0.01; Cohen’s d = 0.45), also confirming the effect of the main interest in the present study. In contrast, we did not find evidence that the modulation was a result of TMS stimulation per se because a comparison of PMd and PMv to Sham was not found significant (µ = −0.05; 95% CI, −0.36-0.25; t = −0.34; P = 0.73; Cohen’s d = −0.06). Note that one participant showed a very large reach direction (around 8.5°) compare to the others in the second half of the PMd condition (Fig. 2c). Since the participant also showed the largest reach directions in the second half of PMv and Sham, it was likely to be the participant’s characteristic, which the random effect in the model would take into account. Indeed, the same statistical significance was preserved when the participant was removed from the data sets, confirming that the results were not biased by the single participant who exhibited large adaptation.
DiscussionThe present study shows that TMS on PMd at go-signal can facilitate visuomotor adaptation in humans. The effect appears region-specific because adaptation is found significantly larger when PMd is compared with PMv and Sham but not when PMd and PMv are compared with Sham. This is in line with previous reports suggesting different roles of PMd and PMv [12,13]. The effect is different from previously reported TMS effects on attenuated online error-correction [21] because adaptation is measured in absence of the cursor feedback and because the stimulation is applied in a preceding trial in the present study. Thus, to our knowledge, this is the first evidence for facilitation of human visuomotor adaptation by noninvasive brain stimulation on PMd.
Interestingly, the direction of modulation in the present study is opposite to the original study, which finds attenuates adaptation by stimulation [10]. One possibility is that TMS in the present study may be facilitating while stimulation is disruptive in the original study. It has been argued that low-intensity TMS tends to facilitate neural activities [25]. Thus, while we use supra-motor threshold intensity primarily to avoid muscle twitch, the intensity may be low enough to facilitate neural activities in PMd instead of disrupting them. Since excitatory changes between PMd and the cerebellum are observed during visuomotor adaptation [7], TMS may facilitate the modulatory processes between the regions, which is manifested as enhanced adaptation. It is worthwhile to examine this possibility in conjunction with how modulation may differ among types of motor learning because a previous study shows attenuated visuomotor association learning using TMS above threshold [5]. Pinpointing the cause of these differences in modulation may provide insights into the general role of PMd in motor learning. Since noninvasive neuromodulation is a promising tool for discovering neural mechanisms of motor learning and for facilitating motor rehabilitation [11], the present study demonstrates that PMd is a feasible target region for pursuing these research and practical goals.
AcknowledgementsThis work was supported by grants from Japan Society for the Promotion of Science KAKENHI (Grant numbers: 19H04977, 19H05729, and 22H00498). T.S. was supported by a JSPS Research Fellowship for Young Scientists (JSPS KAKENHI: 19J20366).
Conceptualization: T.S., K.N., and J.I. Methodology: T.S., K.N., and J.I. Data curation: T.S. and K.N. Funding acquisition and project administration: J.I. Writing – original draft: T.S., K.N., and J.I. Writing – review & editing: T.S. and J.I.
Conflicts of interestThere are no conflicts of interest.
References 1. Weinrich M, Wise SP. The premotor cortex of the monkey. J Neurosci. 1982; 2:1329–1345. 2. Wessberg J, Stambaugh CR, Kralik JD, Beck PD, Laubach M, Chapin JK, et al. Real-time prediction of hand trajectory by ensembles of cortical neurons in primates. Nature. 2000; 408:361–365. 3. Schluter ND, Rushworth MF, Mills KR, Passingham RE. Signal-, set-, and movement-related activity in the human premotor cortex. Neuropsychologia. 1999; 37:233–243. 4. Mushiake H, Inase M, Tanji J. Neuronal activity in the primate premotor, supplementary, and precentral motor cortex during visually guided and internally determined sequential movements. J Neurophysiol. 1991; 66:705–718. 5. Praeg E, Herwig U, Lutz K, Jancke L. The role of the right dorsal premotor cortex in visuomotor learning: a transcranial magnetic stimulation study. Neuroreport. 2005; 16:1715–1718. 6. Praeg E, Esslen M, Lutz K, Jancke L. Neuronal modifications during visuomotor association learning assessed by electric brain tomography. Brain Topogr. 2006; 19:61–75. 7. Tzvi E, Koeth F, Karabanov AN, Siebner HR, Krämer UM. Cerebellar - Premotor cortex interactions underlying visuomotor adaptation. Neuroimage. 2020; 220:117142. 8. Tseng YW, Diedrichsen J, Krakauer JW, Shadmehr R, Bastian AJ. Sensory prediction errors drive cerebellum-dependent adaptation of reaching. J Neurophysiol. 2007; 98:54–62. 9. Izawa J, Criscimagna-Hemminger SE, Shadmehr R. Cerebellar contributions to reach adaptation and learning sensory consequences of action. J Neurosci. 2012; 32:4230–4239. 10. Vyas S, O’Shea DJ, Ryu SI, Shenoy KV. Causal role of motor preparation during error-driven learning. Neuron. 2020; 106:329–339.e4. 11. Reis J, Robertson E, Krakauer JW, Rothwell J, Marshall L, Gerloff C, et al. Consensus: “Can tDCS and TMS enhance motor learning and memory formation?”. Brain Stimul. 2008; 1:363–369. 12. Kurata K, Hoffman DS. Differential effects of muscimol microinjection into dorsal and ventral aspects of the premotor cortex of monkeys. J Neurophysiol. 1994; 71:1151–1164. 13. Hoshi E, Tanji J. Contrasting neuronal activity in the dorsal and ventral premotor areas during preparation to reach. J Neurophysiol. 2002; 87:1123–1128. 14. Hoshi E, Tanji J. Distinctions between dorsal and ventral premotor areas: anatomical connectivity and functional properties. Curr Opin Neurobiol. 2007; 17:234–242. 15. Rothwell JC, Hallett M, Berardelli A, Eisen A, Rossini P, Paulus W. Magnetic stimulation: motor evoked potentials. The international federation of clinical neurophysiology. Electroencephalogr Clin Neurophysiol Suppl. 1999; 52:97–103. 16. Fink GR, Frackowiak RS, Pietrzyk U, Passingham RE. Multiple nonprimary motor areas in the human cortex. J Neurophysiol. 1997; 77:2164–2174. 17. Schluter ND, Rushworth MF, Passingham RE, Mills KR. Temporary interference in human lateral premotor cortex suggests dominance for the selection of movements. A study using transcranial magnetic stimulation. Brain. 1998; 121(Pt 5):785–799. 18. Mills KR, Boniface SJ, Schubert M. Magnetic brain stimulation with a double coil: the importance of coil orientation. Electroencephalogr Clin Neurophysiol. 1992; 85:17–21. 19. Hinder MR, Tresilian JR, Riek S, Carson RG. The contribution of visual feedback to visuomotor adaptation: how much and when? Brain Res. 2008; 1197:123–134. 20. Leow LA, Marinovic W, de Rugy A, Carroll TJ. Task errors drive memories that improve sensorimotor adaptation. J Neurosci. 2020; 40:3075–3088. 21. Lee JH, van Donkelaar P. The human dorsal premotor cortex generates on-line error corrections during sensorimotor adaptation. J Neurosci. 2006; 26:3330–3334. 22. Verstynen T, Sabes PN. How each movement changes the next: an experimental and theoretical study of fast adaptive priors in reaching. J Neurosci. 2011; 31:10050–10059. 23. Bates D, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. Journal of Statistical Software. 2015; 67:1–48. 24. Kuznetsova A, Brockhoff PB, Christensen RH. lmerTest package: tests in linear mixed effects models. J Stat Softw. 2017; 82:1–26. 25. Silvanto J, Cattaneo Z. Common framework for “virtual lesion” and state-dependent TMS: The facilitatory/suppressive range model of online TMS effects on behavior. Brain Cogn. 2017; 119:32–38.
留言 (0)