Ingesting alcohol by animals date back tens of millions of years, and by humans hundreds of thousands of years (Lamas-Paz et al., 2018; Seitz and Neuman, 2021; WHO, 2018). Nowadays, excessive alcohol (ethanol) consumption is a global public health problem (Andreuccetti et al., 2012; Garcia et al., 2015; Silveira et al., 2014). Ingested alcohol is absorbed through the stomach and intestines and finally transported to the liver via the portal vein (Cederbaum, 2012). The liver has a myriad of functions, such as immunity from disease and support of the digestion and metabolism of xenobiotic compounds, including ethanol (EtOH) (Hyun et al., 2021). Ethanol is oxidized to acetaldehyde by the cytosolic enzyme alcohol dehydrogenase (ADH), which is further oxidized to acetate by aldehyde dehydrogenase (ALDH) (Ceni et al., 2014; Ghosh Dastidar et al., 2018). Long-term alcohol intake also induces the microsomal ethanol-oxidizing system (MEOS), a process which often generates hepatocyte-toxic free radicals (Lu and Cederbaum, 2018), in turn leading to liver damage by alcoholic liver disease (ALD) (Hosseini et al., 2019), also known as alcohol-related liver disease (Szabo et al., 2019). ALD encompasses a broad spectrum of liver disorders, ranging from simple steatosis (fatty liver), alcoholic hepatitis (AH), alcoholic liver fibrosis and alcoholic cirrhosis (Nagata et al., 2007; Tang et al., 2014).
A growing body of literature shows that people subject to health-care inequality are more vulnerable to both chronic ethanol consumption (WHO, 2018) and neglected tropical diseases, with profound impact on human health (Houweling et al., 2016). Intestinal schistosomiasis mansoni is a neglected parasitic disease caused by the blood-dwelling flatworm Schistosoma mansoni, which poses a serious threat to health, social and economic development of people living under poor water, sanitation and hygiene (WASH) conditions in rural areas in sub-Saharan Africa, Brazil and the Caribbean islands (WHO, 2016).
The complex life cycle of this parasite includes a broad range of definitive mammalian hosts (humans, non-human primates and rodents) and intermediate freshwater snail hosts (Biomphalaria spp.). Mammalian hosts become infected when they come into contact with water containing cercariae (free-swimming larval stages) expelled by infected snails into the environment. Upon skin penetration of a mammalian host, the cercariae transform into schistosomula, invade a blood vessel and migrate through the heart and lungs to the liver where they attain sexual maturity (Costain et al., 2018). After around four weeks, paired worms migrate to the mesenteric veins to lay eggs on the endothelial lining of capillary walls (Andrade, 2009). During the pre-patent phase of S. mansoni infection, Th1 cells play a pro-inflammatory role. After egg laying (week 5 onwards), soluble egg antigen (SEA) shifts the Th1 profile to a Th2-mediated immune response (Colley and Secor, 2014).
Eggs either migrate through the intestinal tissue toward the gut lumen, to be expelled in the feces, or are carried by the blood flow into the small liver sinusoids (Costain et al., 2018). A large number of schistosome eggs trapped in these sites induces a granulomatous inflammatory process and tissue pathology (Andrade, 2009; Cheever et al., 2002; Colley and Secor, 2014). Many cells, such as lymphocytes, macrophages, eosinophils, basophils, monocytes and fibroblasts, cause the granulomatous inflammation (Andrade, 2009; Pearce, 2005; Schwartz and Fallon, 2018). However, granulomas play a protective role by limiting SEA to the intestine and liver (Amiri et al., 1992; Hams et al., 2013). Schistosomiasis mansoni can be categorized into acute (8–10 weeks after infection) and chronic phases (12 weeks onward). During acute infection in mice, hepatic granulomas are classified into three categories (exudative, exudative-productive and productive), according to cellular composition and evolution stage (Lenzi et al., 1998).
Previous studies have indicated that schistosomiasis mansoni and chronic alcohol consumption independently cause chronic liver disease (CLD), but through different mechanisms (Andrade, 2009; Boye et al., 2016; Lambertucci, 2014). Although both factors independently coexist among Brazilian patients, a correlation between alcohol abuse and the intensity of periportal thickening has not been found (Cota et al., 2006). In animal models of ethanol intake, hepatic granulomas were reduced in diameter after 7% (Castro et al., 1993) or 35% exposure (Orrego et al., 1981) in mice. Our preliminary experiments have for the first time shown that chronic ethanol intake causes morphological changes of the tegument of male S. mansoni and the reproductive system of both male and female worms (Brandão-Bezerra et al., 2019). To our knowledge, the effect of comorbidity on liver pathology remains unclear. Using the mouse model of S. mansoni acute infection and long-term ethanol intake, we attempted to address this gap, using both histopathological and stereological evaluations.

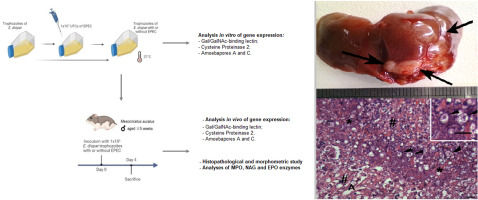
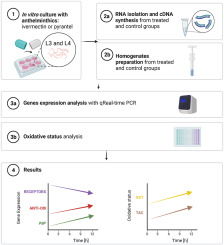


留言 (0)