
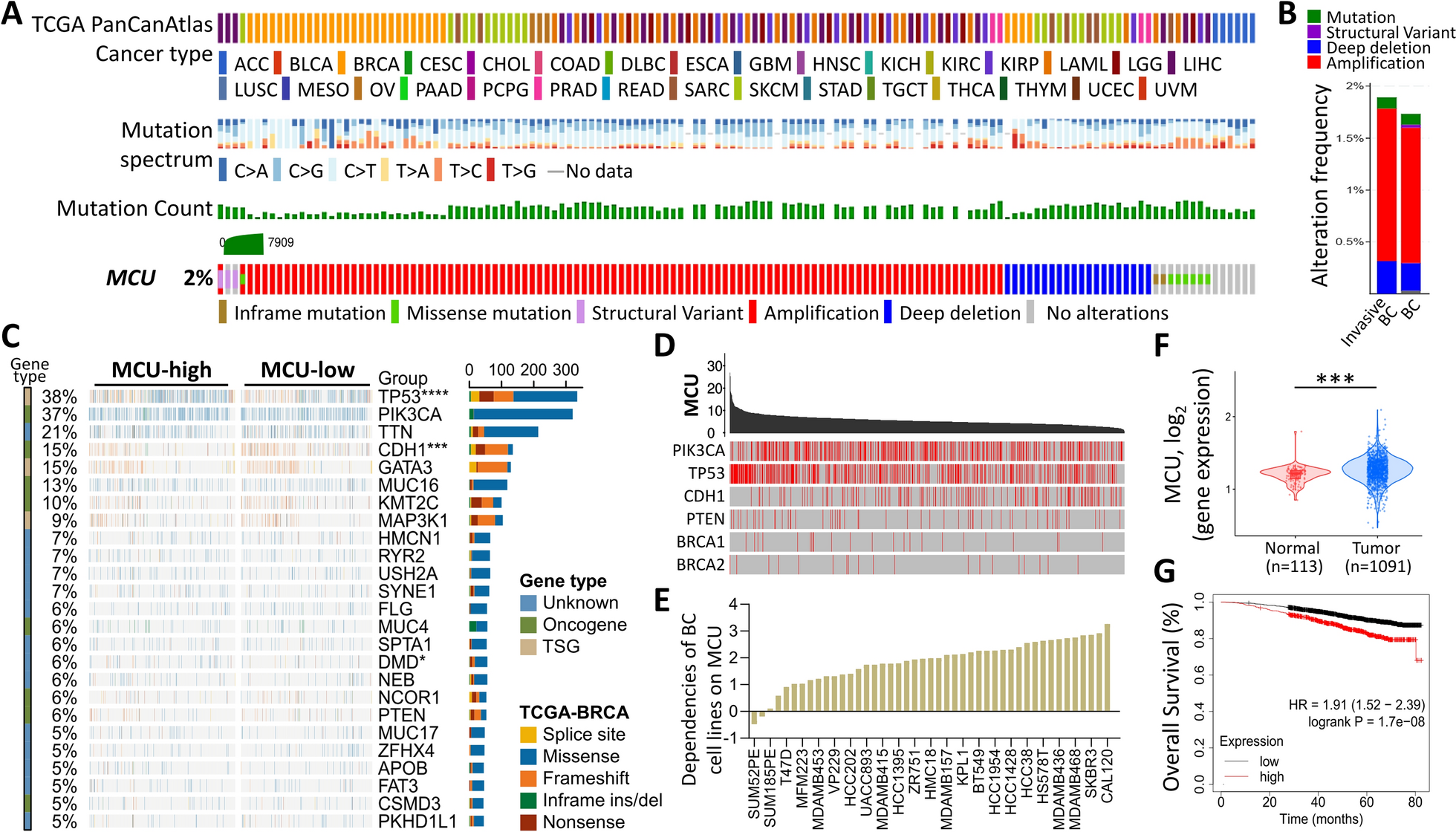
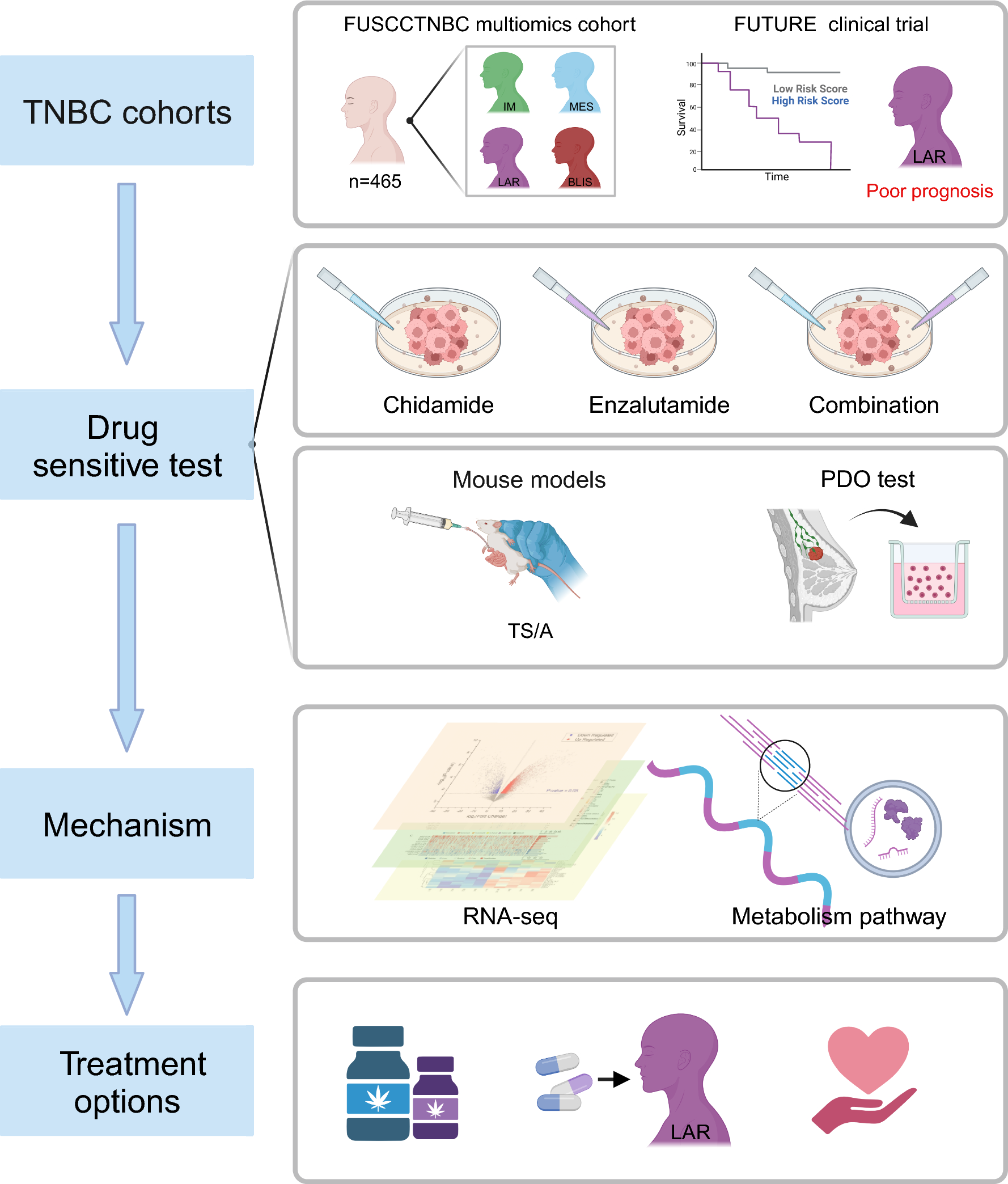
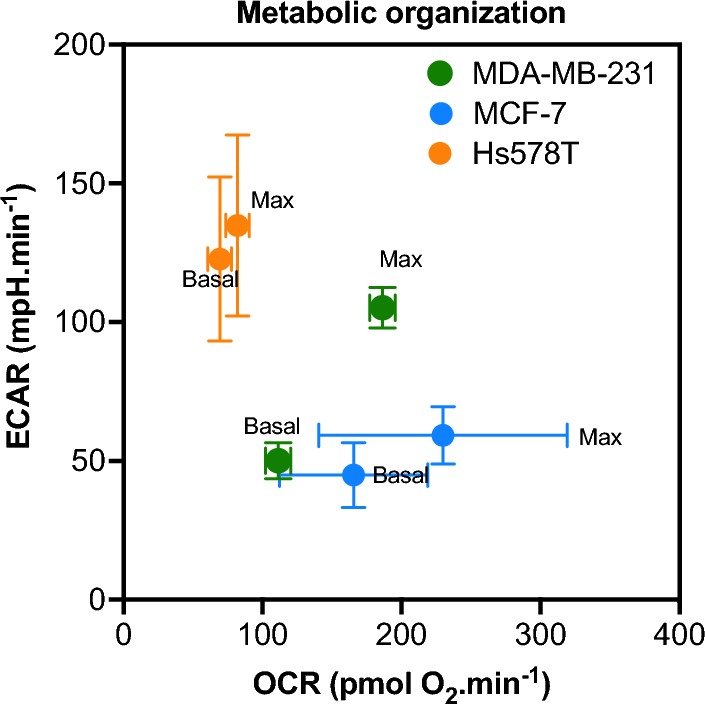
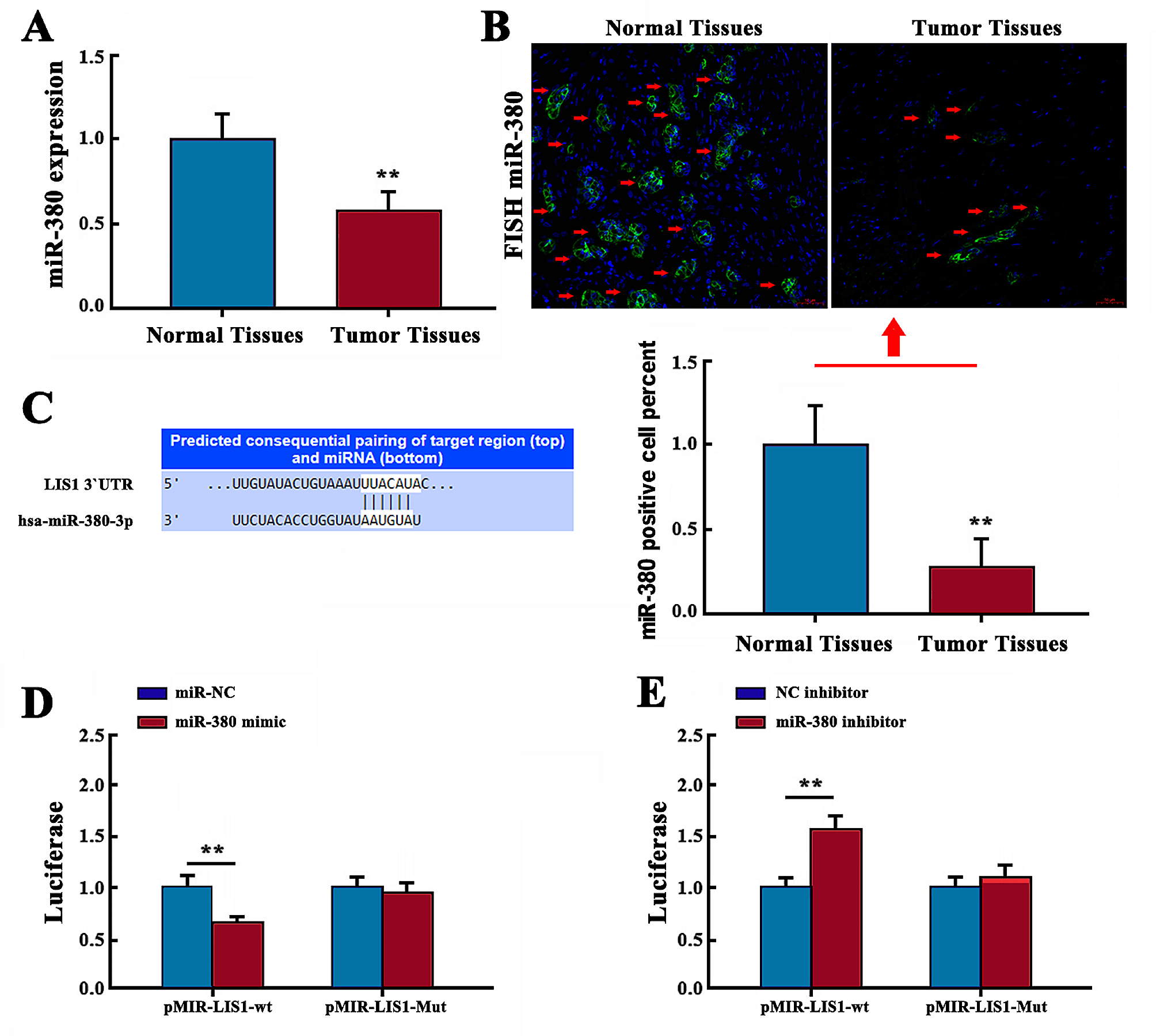
記住我
To systematically dissect the potential similarities and differences between UC-NKs and P-NKs, we took advantage of the “3IL”-based strategy (hIL-2, hIL-15 and hIL-18) for in vitro amplification and activation of NK cells from mononuclear cells (MNCs) as we recently reported [2]. Generally, both the seeded initial UC-MNCs and P-MNCs (day 0) showed a unicellular state, followed by a clustered intermediate morphology (day 7, day 10), and finally the dense and incompact state (day 14) (Fig. 1A). As shown by the FCM diagrams and statistical analyses, a higher percentage of total CD3−CD56+ NK cells as well as the representative activated CD3−CD56+CD16+ NK cell subset was observed in P-MNCs (day 0) and the derivations including P-NKs (day 14) over that in the corresponding UC-MNCs and UC-NKs, respectively (Fig. 1B, C). Simultaneously, total cell number and total NK cells revealed gradual and constant increase during the process of in vitro culture, and in particular, those in the P-NKs group (Fig. 1D, E). Interestingly, the fold change of total P-NK cells was comparable to that in the UC-NKs after the 14-days’ induction (Fig. 1E). Additionally, besides NK cells, a small and higher proportion of CD3+ lymphocytes and CD3+CD4+ Th cells together with CD3+CD56+ NKT-like cells were observed in the UC-NKs group as well (Fig. 1F, G).
Fig. 1
Comparison of the cytomorphology and components of UC-NKs and P-NKs. A Phase contrast images of UC-NKs and P-NKs at the indicated time points (day 0, 7, 10, 14) during the 14-days’ ex vivo induction. Scale bar = 50 μm. B, C FCM diagrams B and statistical analyses C of total (CD3−CD56+) and activated (CD3−CD56+CD16+) UC-NKs and P-NKs. D, E Statistical analyses of the numbers of total (CD3−CD56+) UC-NKs and P-NKs D and the activated (CD3−CD56+CD16+) UC-NKs and P-NKs E. F, G FCM diagrams F and statistical analyses G of total CD3+CD56− T lymphocytes, CD4+ T cells, CD8+ T cells and CD3+CD56+ NKT cells in UC-NKs and P-NKs. All data were shown as mean ± SEM (N = 3 independent experiments). *, P < 0.05; **, P < 0.01; ***, P < 0.001; NS, not significant
UC-NKs and P-NKs revealed diversity in subpopulations and cellular vitalityIn addition to generating comparable amount of NK cells from UC-MNCs and P-MNCs, we were curious about whether there’re any similarities and differences between the cytotoxic phenotype and function of the derived UC-NKs and P-NKs. For the purpose, we assessed the NK cell phenotype by monitoring the expression of various activating surface biomarkers, which were involved in regulating the dynamic equilibrium of NK cell activation and inhibition as we and other investigators reported [1, 2, 8]. As to the resting NK cells in the UC-MNCs and P-MNCs, both of them showed minimal expression of NKG2D, NKp44 and NKp46 but revealed higher level of NKG2A expression instead, which were collectively distinguish from the UC-NKs and P-NKs after ex vivo expansion (Fig. 2A, B). Interestingly, the proportion of NK cells with NKp44 or NKp46 expression in the UC-NKs group was moderately higher than that in the P-NKs group at day 14 of ex vivo culture (Fig. 2A, B).
Fig. 2
Comparison of the subpopulations and cellular vitality of UC-NKs and P-NKs A, B FCM diagrams A and statistical analyses B of the subpopulations of NK cells (NKG2D+, NKG2A+, NKp44+, NKp46+) in UC-NKs and P-NKs at day 0 and day 14 of ex vivo expansion and activation. C Fold change of total cells and NK cells at the indicated time points (day 0, 7, 10, 14) during the 14-days’ ex vivo induction. D, E FCM diagrams D and statistical analyses E of the distribution of sub-stages in cell cycle. F, G FCM diagrams F and statistical analyses G of the apoptotic cells in UC-NKs and P-NKs. All data were shown as mean ± SEM (N = 3 independent experiments). *, P < 0.05; **, P < 0.01; NS, not significant
Cellular vitality has been recognized as the fundamental for biological efficacy in cytotherapy [21, 22]. According to the curve of cumulative population, the fold change of total cells rather than the NK cells in the P-NK group was moderately higher than that in the UC-NK group (Fig. 2C). Simultaneously, the distributions of sub-stages of cell cycle in the UC-NK and P-NK groups revealed statistical differences whereas most of the cells consistently stayed in the G0/G1 stage with minimal difference (Fig. 2D, E). Furthermore, as shown by the FCM diagram based on the Annexin V and 7-AAD staining, the proportion of apoptotic NK cells in the UC-NK group after 14-days’ ex vivo expansion was a little higher than that in the P-NK group (Fig. 2F, G). Taken together, both the resting and expanded NKs (day 0, day 10) in the P-NKs group manifested diversity in cytotoxic phenotype and cellular vitality compared to the UC-NK group.
UC-NKs and P-NKs exhibited distinguishable landscape of gene expression profileHaving disclosed the biological properties between UC-NKs and P-NKs, we next try to estimate the potentially genetic characteristics with the aid of RNA-SEQ. Generally, UC-MNCs (UC_day 0) and P-MNCs (P_day 0) as well as UC-NKs (UC_day 14) and P-NKs (P_day 14) showed more similarities in gene expression distributions, which was confirmed by the HeatMap diagram of Pearson values (Fig. 3A, B). Simultaneously, principal component analysis (PCA) of the transcriptome data including PC1 (66.34%) and PC2 (23.44%) revealed a clear clustering between UC-NKs and P-NKs (Fig. 3C).
Fig. 3
The landscape of gene expression profile of UC-NKs and P-NKs A The distribution of genes with various ranges of FPKM among UC-MNCs (UC_d0), UC-NKs (UC_d14), P-MNCs (P_d0), P-NKs (P_d14). B The correlation of the aforementioned groups. C Principal component analysis (PCA) of the aforementioned groups. D Volcano plot assay of up-regulated DEGs (Up), downregulated (Down) DEGs and non DEGs (no-DEG) between UC-NKs (UC_d14) and P-NKs (P_d14). E, F Venn map diagram E and Hierarchical cluster analysis F of DEGs among the indicated groups. G, H Gene Ontology Biological Process (GOBP) analysis of the DEGs G and differentially expressed transcripts H between UC-NKs (UC_d14) and P-NKs (P_d14)
Furthermore, volcano plot analysis intuitively exhibited the expression pattern of upregulated (Up, in red), downregulated (Down, in green) and not differentially expressed genes (no-DEGs, in gray), respectively (Fig. 3D). As shown by the Venn map diagram, a total number of 7,427 DEGs were clustered into categories seven categories (Fig. 3E). In consistence, unsupervised hierarchical clustering analysis on the basis of FPKM values of the DEGs also disclosed that UC-NKs (UC_day 14) and P-NKs (P_day 14) closer evolutionary relationship than UC-MNCs (UC_day 0) or P-MNCs (P_day 0) (Fig. 3F). To obtain further insight into the similarities and differences between UC-NKs (UC_day 14) and P-NKs (P_day 14), we conducted gene ontology (GO) analysis and found that the DEGs were principally associated with immunity (e.g., immune response, immune system process, regulation of immune process), signal regulation (e.g., cytokine-mediated signaling pathway, signal transduction, cell surface receptor signaling pathway, IFN-γ mediated signaling pathway) and antigen presentation, while the differentially expressed transcripts were mainly involved in biological process (e.g., regulation of transcription, methylation, protein phosphorylation, metabolic process) and immune system process instead (Fig. 3H).
Potential variations of UC-NKs and P-NKs in multifaceted biological processesTo further clarify the potential differences between UC-NKs (UC_day 14) and P-NKs (P_day 14), we took advantage of the KEGG pathway analysis, and found that cell adhesion molecule (CAM), cytokine-cytokine receptor interaction and T lymphocyte differentiation (e.g., Th1, Th2 and Th 17 cells) relevant signaling pathways were representatively enriched based on DEGs (Fig. 4A). Instead, a series of signaling pathways such as NF-kB, AMPK, TNF and neurotrophin signaling pathway were solely enriched based on differentially expressed transcripts between UC-NKs (UC_day 14) and P-NKs (P_day 14) (Fig. 4B).
Fig. 4
Potential variations of UC-NKs and P-NKs in multifaceted biological processes A, B KEGG analysis of the DEGs A and differentially expressed transcripts B between UC-NKs (UC_d14) and P-NKs (P_d14). C, E Potential variations of UC-NKs and P-NKs in representative signaling pathways C and biological processes D, E. F PPI analysis of the 412 DEGs (log2FC > 1.0) between UC-NKs (UC_d14) and P-NKs (P_d14). G KDA analysis of the 204 DEGs (log2FC > 2.0) between UC-NKs (UC_d14) and P-NKs (P_d14)
In details, by utilizing the gene set enrichment analysis (GSEA), we further noticed that the datasets were uniquely enriched in immunoregulation-associated signaling pathways including IL6-JAK/STAT3 signaling (FDR = 0.00493) and KRAS signaling (FDR = 0.00020) rather than P53 pathway (FDR = 0.97013) and TNF-α signaling (FDR = 0.12045) (Fig. 4C). In consistent with the KEGG analysis, a series of tumor immunosurveillance-associated bioprocesses were representatively enriched such as coagulation, complement, coagulation and complement cascades, inflammatory response and epithelial-mesenchymal transition (EMT) (Fig. 4D, E). Furthermore, protein–protein interaction (PPI) analysis based on the STRING and DIAMOND database indicated the 412 DEGs (Log2|P-NKs/UC-NKs|> 1, Q value < 0.05, N = 412) between UC-NKs (UC_day 14) and P-NKs (P_day 14) (Fig. 4F). Additionally, with the aid of KDA analysis, the spatial distribution and interaction of the representative 204 DEGs (Log2|P-NKs/UC-NKs|> 2, Q value < 0.05, N = 204), and in particular, the pivotal immunoregulation-related genes (e.g., CTLA4, VCAM-1, CD28, ITGAX, CD68 and CD86) are intuitively displayed (Fig. 4G). Collectively, our results implicated the multifaceted variations of UC-NKs and P-NKs at transcriptome level.
P-NKs manifested superiority in cytotoxicity over UC-NKs in vitroHaving illuminated the multidimensional diversities in cellular phenotypes and molecular features, we turn to explore the in vitro cytotoxic activity of UC-NKs and P-NKs. We took advantage of tumor-killing model by coculturing the indicated NK cells with the K562 human chronic myeloid leukemia (CML) cell line as we recently reported [2]. On the one hand, the proportions of P-NK cells with CD107a expression at most of the effector-to-target ratios (E: T = 1:1, 1:3, 1:5) was higher than those in the UC-NKs groups (Fig. 5A, B). On the other hand, P-NK cells revealed preferable killing capacity (E: T = 1:3, 1:5) over UC-NK cells according to the analysis of residual living K562 cells (Fig. 5C, D).
Fig. 5
Comparison of the in vitro cytotoxicity upon tumor cell lines A, B FCM diagrams A and statistical analyses B of CD107a+ NK cells in coculturing with UC-NKs and P-NKs at various effector-to-target ratios (E: T = 1:1, 1:3, 1:5). C, D The comparison of the cytotoxicity of UC-NKs and P-NKs against K562 cells at the indicated E:T ratios. E, F FCM diagrams E and statistical analyses F of CD107a+ NK cells in coculturing with UC-NKs and P-NKs at the indicated effector-to-target ratio (E: T = 1:5). G, H The comparison of the cytotoxicity of UC-NKs and P-NKs against HT29 cells at the indicated E:T ratio (E: T = 1:5). All data were shown as mean ± SEM (N = 3 independent experiments). *, P < 0.05; **, P < 0.01; NS, not significant
To further verify the cytotoxicity of the aforementioned NK cells, we conducted P-NK cells or UC-NK cell coculture with HT29 human colorectal adenocarcinoma cell line as we reported before [17]. Similarly, the percentage of CD107a+ P-NK cells at the indicated effector-to-target ratios (E: T = 1:5) was higher than that in the UC-NK cell group (Fig. 5E, F). Simultaneously, P-NK cells revealed preferable killing capacity (E: T = 1:5) over UC-NK cells according to the analysis of residual living HT29 cells (Fig. 5G, H). Collectively, our data indicated the moderate superiority of P-NK cells in cytotoxicity over UC-NKs in vitro.
留言 (0)