




記住我

Toxoplasmosis, an obligate intracellular parasitic zoonotic disease caused by Toxoplasma gondii (T. gondii), is widespread around the world and infects almost all warm-blooded animals, including humans (Nasiru Wana et al., 2020b; Matta et al., 2021). It is known to cause reproductive failure, resulting in huge economic losses owing to abortion and weak offspring in food animals (Stelzer et al., 2019; Nayeri et al., 2021). Studies have shown that most animals are infected by the ingestion of food and water with oocysts from cat feces, and humans are infected due to consumption of raw and undercooked meat containing bradyzoites or oocysts (Uddin et al., 2021). Approximately one-third of the human population is exposed to T. gondii infection through oral, blood and congenital transmission (Abbas et al., 2020; Nasiru Wana et al., 2020a). Felids, the only final host, are infected by tachyzoites, oocysts, or bradyzoites, while other animals, such as livestock, birds, and fish, can all act as intermediate hosts (Nasiru Wana et al., 2020a; S. Al-Malki, 2021). T. gondii-infected cats are involved in the transmission of T. gondii, but clinical symptoms of T. gondii infection usually go unnoticed and cannot be used for early diagnosis (Boothroyd and Grigg, 2002). Moreover, pet owners are at risk of T. gondii infection due to the unknown relationship between animal-to-human transmission and pet feeding. At the same time, stray cats and dogs are adopted as companion pets, and unclear infection status may be responsible for the transmission of T. gondii to humans. Therefore, rapid, sensitive, and on-site detection technology of T. gondii is vital for early infection warning, which helps prevent animal-to-human transmission and effectively maintain public and veterinary health (Uddin et al., 2021).
The diagnosis of T. gondii infection frequently depends on serological testing under laboratory conditions and is limited by shortcomings such as being time-consuming, laborious, a health hazard and requiring skilled personnel (Kotresha and Noordin, 2010; Liu et al., 2015). At present, various molecular biological methods with increased sensitivity and specificity have attracted researchers’ attention and have got a significant achievement in the detection of T. gondii (Switaj et al., 2005; Ybanez et al., 2020), such as conventional PCR, quantitative PCR (qPCR), and loop-mediated isothermal amplification (LAMP). PCR has been widely used for T. gondii detection since T. gondii detection was achieved targeting the B1 gene in 1989 (Burg et al., 1989), and qPCR is capable of identifying low copies of target genes and evaluating disease progression (Teixeira et al., 2013). However, skilled personnel and expensive equipment are indispensable for PCR and qPCR, limiting their application in resource-poor regions and precluding their use at the point of care. LAMP is a relatively novel, less time-consuming and simple diagnostic technology under isothermal conditions (Hegazy et al., 2020), and aerosol contamination and reaction mixture instability are the main barriers for accurate detection. Moreover, the use of chromogenic agents, such as hydroxy naphthol blue (HNB), in the LAMP system allows results determination with the naked eye following a visual colour change from violet to blue, thereby reducing the dependence on equipment (Goto et al., 2009; Sheikh et al., 2020). The sensitivity and specificity of molecular biological methods are also affected by DNA extraction technology (Edvinsson et al., 2004). Extraction of T. gondii DNA is time-consuming, tedious and ill-suited under field conditions, also limiting the application of nucleic acid-based molecular biological methods (Liu et al., 2015). In addition, it is not clear that if the DNA in T. gondii spores was extracted effectively during the sample treatment, which may lead to faulty detection.
Herein, on the basis of an established membrane-based nucleic acid extraction method, we aimed to develop rapid, sensitive and specific qLAMP and visual LAMP assays for T. gondii detection by optimizing the reaction system and determining the procedure, while protective reagents were also used to improve the stability of the LAMP system, achieving point-of-care testing, equipment independence and real-time detection and proposing a novel strategy for T. gondii investigation, prevention and control.
Materials and methodsSample and animalsThe DNA of T. gondii, Giardia lamblia, Trichomonas foetus, Cryptosporidium muris and Neospora caninum were stored in Shandong Key Lab of Preventive Veterinary Medicine, College of Veterinary Medicine, Qingdao Agriculture University (QAU). T. gondii clinical cat faeces samples were collected from pet hospitals in Qingdao, Shandong province.
Design of T. gondii LAMP primersThe conserved region sequences of T. gondii in the SAG1 gene were retrieved from GenBank (GenBank accession no. X14080), and the T. gondii LAMP primers were designed by Primer Explorer V5 to amplify a fragment about 202 bp. The oligonucleotide sequences of the primers are shown in Table 1. The optimal primers were analysed and screened by Oligo 6.0 and synthesized by Shanghai Sangon Biotech (China).
TABLE 1
Table 1 T. gondii-specific primer sets used in this study.
Construction and identification of recombinant plasmidThe SAG1 gene sequences of T. gondii were retrieved from GenBank (GenBank accession no. X14080) and synthesized by Shanghai Sangon Biotech, and the recombinant plasmid was constructed by linking it with the pMD18-T vector. The mass concentration of the recombinant plasmid was measured and stored at -20°C after the sequence was identified.
Preparation of T. gondii DNAWhatman™ FTA™ Elute Cards (GE Healthcare Life Sciences, MA, USA) were used for room temperature collection, preservation, and purification of nucleic acids from samples for detection. Briefly, the faeces samples were treated with 50 μL SEMP (containing 1 M Tris-HCl, 700 mM EDTA, 10% SDS, β-mercaptoethanol and equilibrium phenol) (Solarbio, China) in a 1.5 mL nuclease-free tube (Cao et al., 2020). After full homogenization, 0.2 mg/mL chitinase was added at room temperature for 30 min to remove the impact of spores for DNA extraction. Nucleic acid was absorbed onto FTA elute cards, which were cut into 2 mm discs using a sterile 2 mm Harris punch (WB100039) and a cutting mat, and 2 mm discs were punched out and placed into the homogenate, stirring the nucleic acid adsorption material with tweezers to fully wet it. The FTA discs were washed twice, and nucleic acids were eluted with 20 μL sterile water to obtain T. gondii DNA. The concentration of nucleic acids was measured using a NanoDrop2000 and stored at -80°C. Finally, 1 μL of eluant was added to the qLAMP and visual LAMP systems for detection.
Preembedding and freeze-drying of detection reagentsTube cap preembedding: Neutral red (Solarbio, China) was added to the tube cap of PCR tubes and dried at room temperature. Then, paraffin at 52°C to 54°C was added to cover with neutral red, finishing the tube cap preembedding process.
Reagents preembedding: Mixed detection reagents, specific primers and protectants were added to the PCR tubes and placed in a freeze dryer. Lyophilization was started with prefreezing for 1 h below −40°C and then vacuumed to 0.08–0.10 mbar, followed by heating at −5°C for 1.5 h (Cao et al., 2020).
qLAMPFirst, the reaction system was determined by optimizing the concentrations of Mg2+ (2 mM, 4 mM, 6 mM, 8 mM) (NEB, USA) and dNTPs (1.0 mM, 1.2 mM, 1.4 mM, 1.6 mM) (NEB, USA). The best reaction system of qLAMP was determined by analysing the reproducibility and the Ct value. Each run contained diluted T. gondii positive standard plasmid (9.2×106 copies/μL) and a negative control without plasmid. The amplification was carried out in the C1000 thermal cycler (BioRad, United States) with a reaction condition of 65°C for 1 min, totally 60 cycles.Three different rooms were used for template preparation, qLAMP master mix preparation, and amplification by qLAMP to circumvent any carry-over contamination with amplified products. The optimal reaction system and conditions were determined as shown in Table 2.
TABLE 2
Table 2 Details of preembedded reagents for detection.
Visual LAMP reactionDifferent colors were exhibited by the chromogenic agents. The T. gondii positive standard plasmid was detected with Neutral Red (Solarbio, China) and concentrations that were the same as those used in the established qLAMP reaction. The reaction mixture was incubated with a reaction condition of 65°C for 1 h. The positive results were indicated by a color change from orange to pink.
Sensitivity assayThe limit of detection (LOD) was determined for qLAMP and visual LAMP assays. Serial dilutions of T. gondii positive standard plasmid were prepared from 9.2×107 copies/μL ~ 9.2×100 copies/μL. 9.2×107 copies/μL ~ 9.2×100 copies/μL series of concentrations were measured for qLAMP and visual LAMP. At least three independent experiments of each concentration were assayed, and ddH2O served as the negative control.
Specificity assayThe specificity for T. gondii of qLAMP and visual LAMP assays was evaluated using T. gondii positive standard plasmid and other parasites DNA (Giardia lamblia, Trichomonas foetus, Cryptosporidium muris and Neospora caninum). Three independent experiments for each sample were tested, and ddH2O served as the negative control.
Reproducibility assay of qLAMPThe T. gondii positive standard plasmid was diluted from 9.2×106 to 9.2×104 copies/µL. Each concentration was repeated 3 times, and a one-time qLAMP assay was conducted to perform the intrabatch repeatability test by comparing the standard deviation (SD) and the coefficient of variation (CV). Three qLAMP assays using diluted T. gondii positive standard plasmid were conducted to perform the interbatch repeatability test by comparing the standard deviation and the CV.
Comparison of qLAMP and visual LAMP with TaqMan qPCR using clinical samplesThe diagnostic applicability of visual LAMP and qLAMP assays was evaluated by testing 200 suspected T. gondii clinical cat faeces samples collected from pet hospitals in Qingdao, Shandong province. The cat faeces samples were detected by visual LAMP, qLAMP and TaqMan qPCR (Liu et al., 2015). The feasibility of visual LAMP and qPCR was evaluated by measuring the diagnostic specificity (DSp), diagnostic sensitivity (DSe) and degree of agreement compared with TaqMan qPCR.
Statistical analysisThe calculation of the diagnostic sensitivity (DSe) and diagnostic specificity (DSp) between the two methods was based on the following formula. DSe = TP/(TP + FN) and DSp = TN/(TN + FP), where TP means true-positive cases, FN means false-negative cases, TN means true-negative cases, and FP means false-positive cases. Kappa = (Po – Pc)/(1 – Pc), where Po is the proportion of observed agreements and Pc is the proportion of agreements expected by chance. The degree of agreement was quantified by Cohen’s kappa coefficient (κ), with the following definitions: no agreement (κ ≤ 0), poor agreement (κ = 0.01 – 0.20), fair agreement (κ = 0.21 – 0.40), moderate agreement (κ = 0.41 – 0.60), substantial agreement (κ = 0.61 – 0.80) and perfect agreement (κ = 0.81 – 1.00) (Sim and Wright, 2005). Precision was evaluated by obtaining mean time-to-detection values and standard deviations (SD) of each set of replicates at a given concentration and calculating coefficients of variation (CV = SD/Mean).
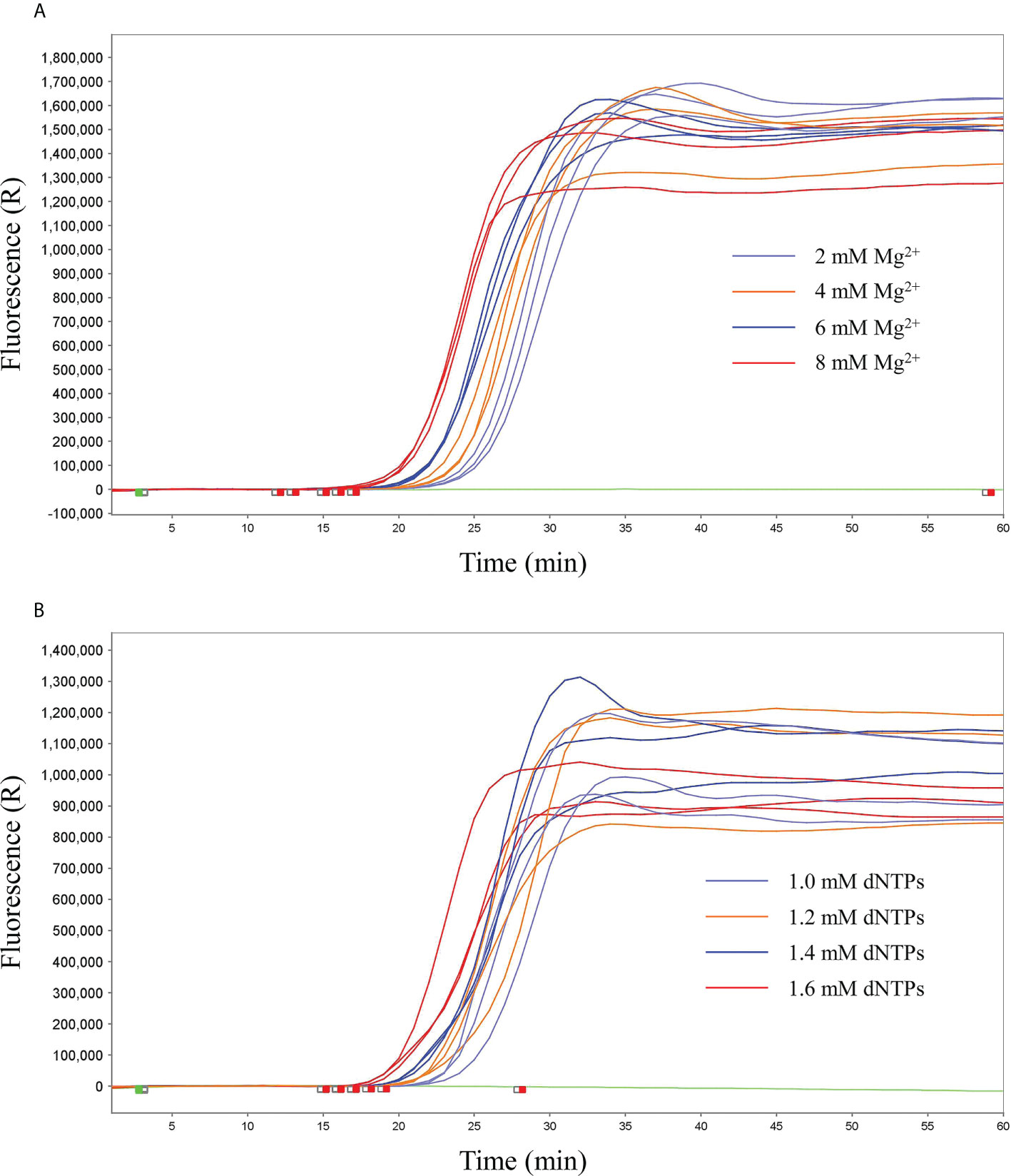
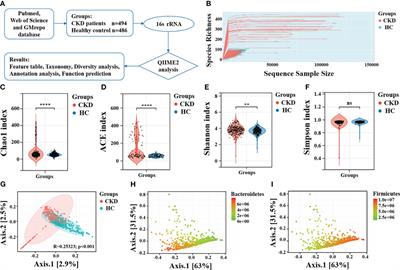
ResultsIdentification of the qLAMP reactionPreliminary experiments were performed only with primers in the absence of any templates to evaluate the negative control precision and revealed no significant primer dimer or self-amplifying reaction. According to the reproducibility and Ct value, the better reproducibility was found with the 8 mM Mg2+ (Ct value, 18 ± 0.25) and 1.4 mM dNTPs (Ct value, 19 ± 0.24) in reaction mixture. Therefore, 8 mM Mg2+ and 1.4 mM dNTPs were determined as the optimal qLAMP reaction concentrations (Figure 1).
FIGURE 1
Figure 1 Screening of optimal concentrations of Mg2+ and dNTPs. (A) Screening of optimal concentrations of Mg2+, 2 mM, 4 mM, 6 mM, and 8 mM. (B) Screening of the optimal concentrations of dNTPs, 1.0 mM, 1.2 mM, 1.4 mM, and 1.6 mM.
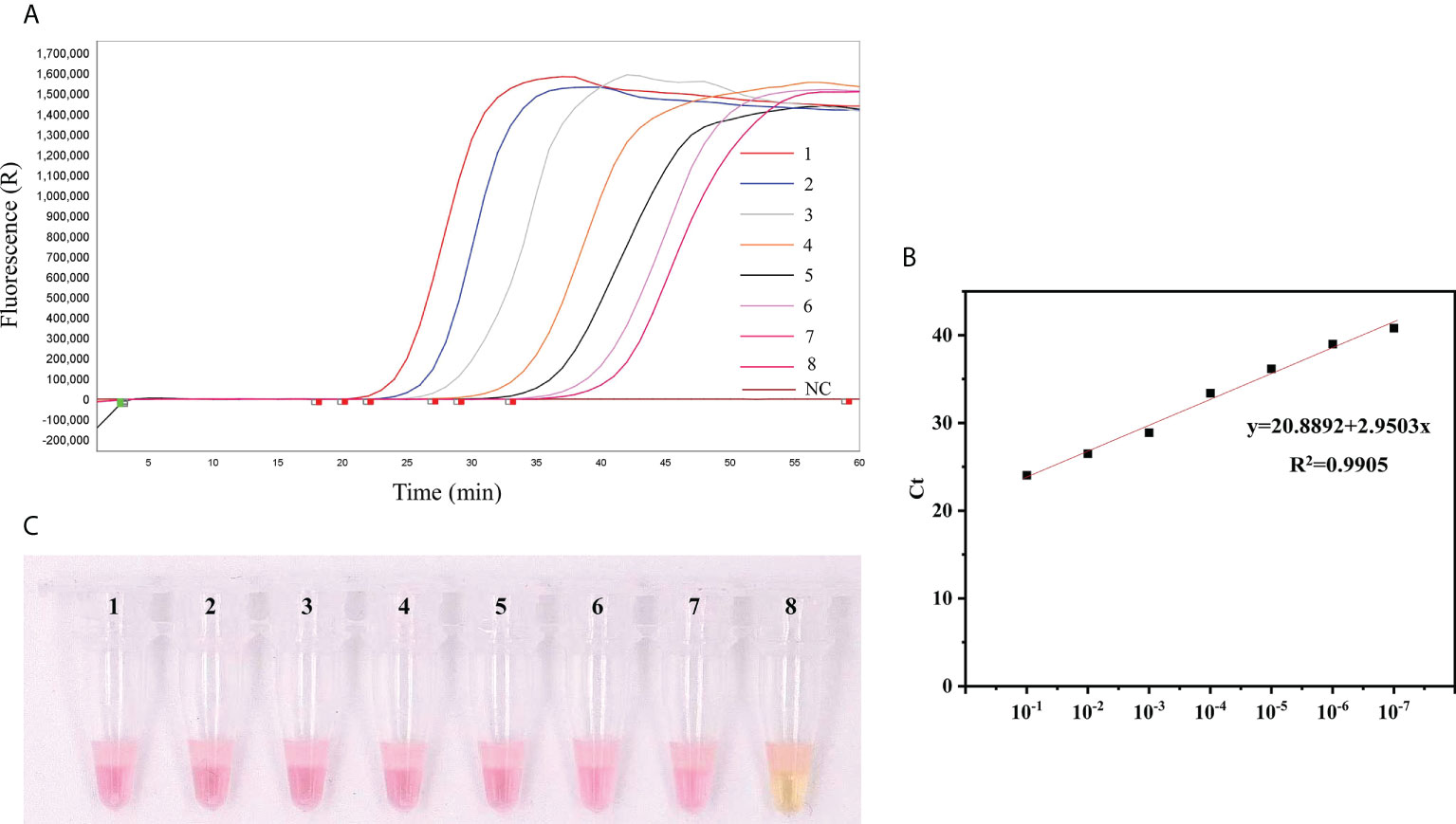
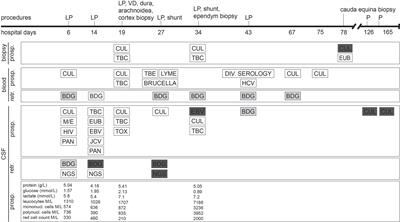
Sensitivity analysisThe sensitivities of qLAMP and visual LAMP (9.2 × 107 ~ 9.2 × 100 copies/µL) assays were assessed using serial dilutions of T. gondii positive standard plasmid DNA. All assays efficiently detected lower copies of T. gondii DNA (Figure 2), and the LODs of the two assays were 92 copies/µL and 92 copies/µL, respectively.
FIGURE 2
Figure 2 Comparison of the sensitivity of qLAMP and visual LAMP. (A) Results of qLAMP analysis. Lanes 1-8: Reaction results from a 10-fold serial dilution of T. gondii positive standard plasmids from 9.2 × 107 to 9.2 × 100 copies/µL per reaction; Lane 9: NC (negative control, nuclease-free water), (B) Standard curve of qLAMP, (C) Results of visual LAMP analysis. Lanes 1-8: 9.2 × 107 to 9.2 × 100 copies/µL.
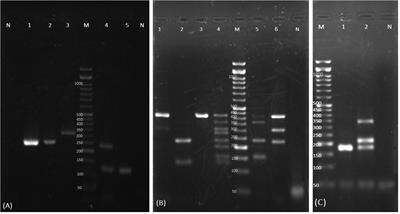
Specificity analysisWe tested the specificity for T. gondii positive standard plasmid DNA of qLAMP and visual LAMP assays. T. gondii-positive standard plasmid DNA, along with Giardia lamblia, Trichomonas foetus, Cryptosporidium muris and Neospora caninum, was used in the specificity test (Figure 3). All three assays only amplified T. gondii, with no cross-reaction with any of the other parasites.
FIGURE 3
Figure 3 Detection specificity of T. gondii positive standard plasmids analysed using qLAMP and visual LAMP. (A) Specificity results for qLAMP, (B) Specificity results for visual LAMP in detecting Giardia lamblia, Trichomonas foetus, Cryptosporidium muris and Neospora caninum. NC, Negative control, nuclease-free water.
Reproducibility analysisThe intrabatch repeatability test and the interbatch repeatability test were measured using the T. gondii positive standard plasmid with concentrations of 9.2×106 ~ 9.2×104 copies/μL by qLAMP. The results showed that the reproducibility performance was excellent, and the intrabatch coefficient of variation (CV, 1.34% ~ 1.68%) and interbatch coefficient of variation (CV, 1.25% ~ 1.72%) were less than 2% (Table 3).
TABLE 3
Table 3 Reproducibility analysis for T. gondii positive standard plasmid DNA by qLAMP.
Performance of qLAMP and visual LAMP for clinical samples compared with TaqMan qPCR testingTo evaluate the practical application of qLAMP and visual LAMP to detect T. gondii, 200 cat faeces samples suspected to be positive for T. gondii were assessed, and the results were compared with those obtained using TaqMan qPCR (Table 4).
TABLE 4
Table 4 Comparison of the tests for T. gondii positive standard plasmid DNA by qLAMP, visual LAMP and TaqMan qPCR.
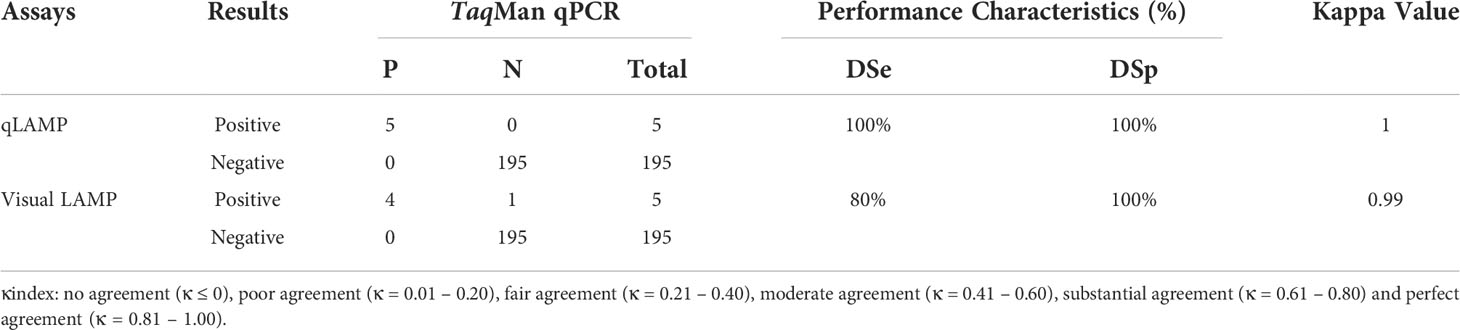
Overall, using TaqMan qPCR, 5 samples were confirmed to be positive for T. gondii DNA, and 195 samples were confirmed to be negative, with undetermined Ct values. qLAMP detected 5 samples as T. gondii-DNA-positive and 195 as negative. Visual LAMP indicated 4 positive and 196 negative samples. Agreement analysis according to clinical sample detection demonstrated that the kappa values between qLAMP and visual LAMP with TaqMan qPCR were 1.0 and 0.99, respectively (Table 4). Additionally, in comparison to TaqMan qPCR, the sensitivity of qLAMP and visual LAMP to identify T. gondii was 100% and 80%, respectively, while the specificity of qLAMP and visual LAMP to identify T. gondii was 100% and 100%, respectively. Thus, qLAMP and visual LAMP showed perfect diagnostic agreement with TaqMan qPCR to detect T. gondii in clinical samples.
DiscussionToxoplasma gondii is considered as one of the most successful eukaryotic pathogens that not only causes huge economic losses to animal husbandry but also harms human health (Liu et al., 2012; Liu et al., 2015). At present, numerous molecular techniques, such as LAMP and qPCR, have been applied for the detection of T. gondii, achieving satisfying performance in sensitivity and specificity. However, pet cats are gradually regarded as popular domestic pet in young population, and the owners are willing to spend money and time on cats. Therefore, we aimed to develop a rapid, accurate and at-home detection technology for T. gondii, meeting the demand of pet owners.
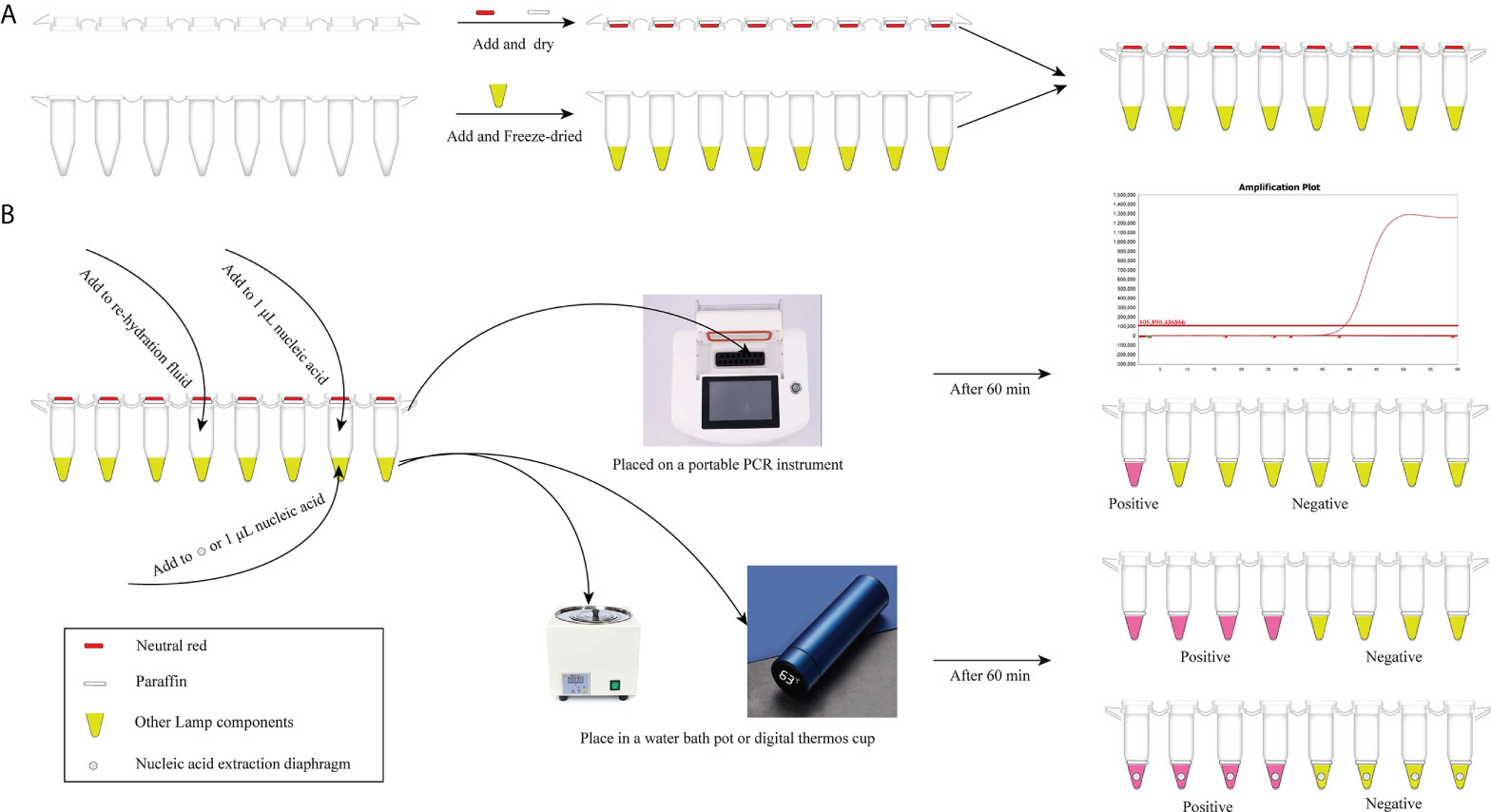
In our study, considering that the aerosol pollution is troublesome for qLAMP and visual LAMP detection, detailed anti-pollution protocols is one of the highlights. We took full advantage of the PCR tubes and lyophilization, weakening dependency on other packing container (Scheme 1). To remove aerosol pollution and improve the accuracy of the detection results, three detailed protocols are designed. Conserved regions (SAG1) were chosen as the target genes for primer design by sequence blasting and sensitivity analysis, improving the specificity of amplification. Then, PCR tube caps and paraffin were utilized to conduct detection reagent preembedding, reducing the nonspecific amplification and preventing aerosol pollution without opening the tube caps. Visual LAMP detection can be completed only with a simple heat block and minimal ease of operation, and qLAMP can be finished with only portable PCR equipment, significantly reducing the time required and achieving a point-of-care test (Scheme 1). Above all, a detailed anti-contamination protocol is used for T. gondii detection, which is convenient for non-professionals to obtain T. gondii detection with no limitations by time and place. Especially, with the gradual increase of dependency for companion pets, convenient, easy-operation and at-home test can attract more attention of pet owners and achieve rapid detection within 60 minutes.
SCHEME 1
Scheme 1 The T. gondii detection procedure with qLAMP and visual LAMP. (A) Preembedding of neutral red and paraffin and freeze-drying of detection reagents. (B) Detection procedure of qLAMP and visual LAMP. qLAMP detection of T. gondii is performed in field conditions with a portable PCR instrument and takes within 60 min. Visual LAMP detection of T. gondii is performed in field conditions with a heating block or vacuum cup within 60 min.
In addition, commercial kits were used to perform DNA extraction, which is time-consuming, tedious, and ill-suited at the POC (Raele et al., 2015). So the membrane technology of nucleic acid extraction is the lightspot to reduce time cost of T. gondii DNA extraction, achieving the goal of professional independence and no requirement for other laboratory precision equipment. Given the impact of T. gondii spores for DNA extraction, chitinase was used to predigested spores to improve the DNA extraction efficiency.
LAMP assay is more widely used, and reagents are easier to obtain, thereby reducing the diagnosis cost. Moreover, modest equipment and ease of operation are needed, making it convenient for use in field conditions (Mori and Notomi, 2009). However, the stability of the LAMP reaction system is an urgent issue, as colour changes of detection reagents can be performed automatically with temperature changing when chromogenic agents exist, resulting in false-positives and limiting the application of LAMP. Studies have shown that trehalose can conduct nonspecific protective actions for biomolecules by replacing hydrones and weakening molecular movement (Sampedro et al., 2001). In our study, 10% trehalose was used in detection reactions, exhibiting satisfactory reproducibility. Therefore, the protective reagent trehalose was added to the detection reagents, and prelyophilization was conducted to improve the stability of the detection reagents, lengthening the storage time of the detection reagents and achieving long-distance transport. Additionally, the use of trehalose can protect the detection reagents from reacting automatically, circumventing false-positive results.
Different target genes, including SAG1, SAG2, B1 and 529-bp REP, can be identified by LAMP, in which SAG1, SAG2 and B1 target genes are used for early diagnosis and to identify T. gondii in human blood samples (Lau et al., 2010; Hu et al., 2012; Qu et al., 2013). The 529-bp REP gene is suitable for reverse transcription LAMP (RT-LAMP) detection in mice and meat samples (Qu et al., 2013). In our previous study, three groups of primers were designed for qLAMP detection targeting SAG1, SAG2 and 529-bp REP genes. We found that lower copies of T. gondii can be identified by SAG1 primers, and considering that early diagnosis of in the blood of T. gondii infected pigs based on SAG1 LAMP (Wang et al., 2013), therefore, the SAG1 gene was ultimately chosen as the target gene to use in qLAMP for the detection of T. gondii.
Herein, qLAMP and visual LAMP assays were developed to quickly diagnose T. gondii in clinical cat faeces samples. Our data showed that the specificities of qLAMP and visual LAMP were all 100% (Figure 3), and their sensitivities were 92 copies/µL and 92 copies/µL, respectively (Figure 2), indicating that these assays based on nucleic acid extraction membrane techniques had good applicability in clinical practice, similar to that of TaqMan qPCR. In addition, our data showed that based on the detection sensitivity of a T. gondii-positive plasmid, the qLAMP assay could detect 92 copies/µL served as suitable tests of clinical samples with low titres (Liu et al., 2015). Moreover, our validation results showed that these two methods were in perfect agreement with those of TaqMan qPCR (with kappa values of 1 and 0.99, respectively) (Table 4). In particular, visual LAMP detection can be completed within 60 minutes, by which the speed and simplicity of the methods make visual LAMP ideally suited for molecular applications both within and outside the laboratory, including limited-resource settings (Scheme 1).
In conclusion, this was the first application of no-equipment DNA extraction for LAMP technology for T. gondii detection. Additionally, trehalose was used to improve the stability of the LAMP reaction system, paraffin was added at preembedding reagents to reduce aerosol pollution, and neutral red was used to achieve visual and point-of-care test detection, minimizing the time cost and providing convenience for pet owners. Our present qLAMP and visual LAMP methods not only have a short turnaround time but also avoid cross-contamination problems and dependence on expensive equipment, which are desirable characteristics amenable to POCT in resource-limited and domestic settings.
Data availability statementThe original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author ContributionsData curation: ZC, KZ, DY and QZ. Formal analysis: JW and YY. Methodology: ZC, KZ, DY, QZ and YY. Project administration: ZC and JW. Supervision: ZC and HN. Writing–original draft: ZC, KZ and DY. Writing–review & editing: ZC and HN. All authors contributed to the article and approved the submitted version.
FundingThe study was supported by the Shandong Modern Agricultural Technology & Industry System (SDAIT-21-13) and the Research Foundation for Distinguished Scholars of Qingdao Agricultural University (663-1120018 and 665-1120046).
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
ReferencesBoothroyd, J. C., Grigg, M. E. (2002). Population biology of Toxoplasma gondii and its relevance to human infection: do different strains cause different disease? Curr. Opin. Microbiol. 5, 438–442. doi: 10.1016/s1369-5274(02)00349-1
PubMed Abstract | CrossRef Full Text | Google Scholar
Burg, J. L., Grover, C. M., Pouletty, P., Boothroyd, J. C. (1989). Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol. 27, 1787–1792. doi: 10.1128/jcm.27.8.1787-1792.1989
PubMed Abstract | CrossRef Full Text | Google Scholar
Cao, Z., Dong, X., Zou, P.-Z., Wan, X.-Y., Li, C., Huang, J. (2020). Development of a cost-efficient micro-detection slide system for the detection of multiple shrimp pathogens. Analytic. Biochem. 599, 113735. doi: 10.1016/j.ab.2020.113735
CrossRef Full Text | Google Scholar
Edvinsson, B., Jalal, S., Nord, C. E., Pedersen, B. S., EvengÅrd, B. (2004). DNA Extraction and PCR assays for detection of Toxoplasma gondii. APMIS 112, 342–348. doi: 10.1111/j.1600-0463.2004.apm1120604.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Goto, M., Honda, E., Ogura, A., Nomoto, A., Hanaki, K.-I. (2009). Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. BioTechniques 46, 167–172. doi: 10.2144/000113072
PubMed Abstract | CrossRef Full Text | Google Scholar
Hegazy, M. K., Awad, S. I., Saleh, N. E., Hegazy, M. M. (2020). Loop mediated isothermal amplification (LAMP) of Toxoplasma DNA from dried blood spots. Exp. Parasitol. 211, 107869. doi: 10.1016/j.exppara.2020.107869
PubMed Abstract | CrossRef Full Text | Google Scholar
Hu, X., Pan, C. W., Li, Y. F., Wang, H., Tan, F. (2012). Urine sample used for detection of toxoplasma gondii infection by loop-mediated isothermal amplification (LAMP). Folia Parasitol. (Praha). 59, 21–26. doi: 10.14411/fp.2012.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Lau, Y. L., Meganathan, P., Sonaimuthu, P., Thiruvengadam, G., Nissapatorn, V., Chen, Y. (2010). Specific, sensitive, and rapid diagnosis of active toxoplasmosis by a loop-mediated isothermal amplification method using blood samples from patients. J. Clin. Microbiol. 48, 3698–3702. doi: 10.1128/jcm.00462-10
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, Q., Singla, L. D., Zhou, H. (2012). Vaccines against Toxoplasma gondii: status, challenges and future directions. Hum. Vaccin. Immunother. 8, 1305–1308. doi: 10.4161/hv.21006
PubMed Abstract | CrossRef Full Text | Google Scholar
Matta, S. K., Rinkenberger, N., Dunay, I. R., Sibley, L. D. (2021). Toxoplasma gondii infection and its implications within the central nervous system. Nat. Rev. Microbiol. 19, 467–480. doi: 10.1038/s41579-021-00518-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Mori, Y., Notomi, T. (2009). Loop-mediated isothermal amplification (LAMP): a rapid, accurate, and cost-effective diagnostic method for infectious diseases. J. Infect. Chemother. 15, 62–69. doi: 10.1007/s10156-009-0669-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Nasiru Wana, M., Mohd Moklas, M. A., Watanabe, M., Nordin, N., Zasmy Unyah, N., Alhassan Abdullahi, S., et al. (2020a). A review on the prevalence of Toxoplasma gondii in humans and animals reported in Malaysia from 2008-2018. Int. J. Environ. Res. Public Health 17 (13), 4809. doi: 10.3390/ijerph17134809
CrossRef Full Text | Google Scholar
Nasiru Wana, M., Mohd Moklas, M. A., Watanabe, M., Zasmy Unyah, N., Alhassan Abdullahi, S., Ahmad Issa Alapid, A., et al. (2020b). Molecular detection and genetic diversity of toxoplasma gondii oocysts in cat faeces from klang valley, Malaysia, using B1 and REP genes in 2018. Pathogens 9 (7), 576. doi: 10.3390/pathogens9070576
CrossRef Full Text | Google Scholar
Nayeri, T., Sarvi, S., Moosazadeh, M., Daryani, A. (2021). Global prevalence of Toxoplasma gondii infection in the aborted fetuses and ruminants that had an abortion: A systematic review and meta-analysis. Vet. Parasitol. 290, 109370. doi: 10.1016/j.vetpar.2021.109370
PubMed Abstract | CrossRef Full Text | Google Scholar
Qu, D., Zhou, H., Han, J., Tao, S., Zheng, B., Chi, N., et al. (2013). Development of reverse transcription loop-mediated isothermal amplification (RT-LAMP) as a diagnostic tool of Toxoplasma gondii in pork. Vet. Parasitol. 192, 98–103. doi: 10.1016/j.vetpar.2012.10.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Raele, D. A., Garofolo, G., Galante, D., Cafiero, M. A. (2015). Molecular detection of coxiella burnetii using an alternative loop-mediated isothermal amplification assay (LAMP). Vet. Ital. 51, 73–78. doi: 10.12834/VetIt.304.1168.4
PubMed Abstract | CrossRef Full Text | Google Scholar
S. Al-Malki, E. (2021). Toxoplasmosis: stages of the protozoan life cycle and risk assessment in humans and animals for an enhanced awareness and an improved socio-economic status. Saudi. J. Biol. Sci. 28, 962–969. doi: 10.1016/j.sjbs.2020.11.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Sampedro, J. G., Cortes, P., Munoz-Clares, R. A., Fernandez, A., Uribe, S. (2001). Thermal inactivation of the plasma membrane h+-ATPase from kluyveromyces lactis. protection by trehalose. Biochim. Biophys. Acta 1544, 64–73. doi: 10.1016/s0167-4838(00)00205-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Sheikh, N., Kumar, S., Sharma, H. K., Bhagyawant, S. S., Thavaselvam, D. (2020). Development of a rapid and sensitive colorimetric loop-mediated isothermal amplification assay: A novel technology for the detection of coxiella burnetii from minimally processed clinical samples. Front. Cell Infect. Microbiol. 10. doi: 10.3389/fcimb.2020.00127
PubMed Abstract | CrossRef Full Text | Google Scholar
Stelzer, S., Basso, W., Benavides Silvan, J., Ortega-Mora, L. M., Maksimov, P., Gethmann, J., et al. (2019). Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne. Parasitol. 15, e00037. doi: 10.1016/j.fawpar.2019.e00037
PubMed Abstract | CrossRef Full Text | Google Scholar
Switaj, K., Master, A., Skrzypczak, M., Zaborowski, P. (2005). Recent trends in molecular diagnostics for Toxoplasma gondii infections. Clin. Microbiol. Infect. 11, 170–176. doi: 10.1111/j.1469-0691.2004.01073.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Teixeira, L. E., Kanunfre, K. A., Shimokawa, P. T., Targa, L. S., Rodrigues, J. C., Domingues, W., et al. (2013). The performance of four molecular methods for the laboratory diagnosis of congenital toxoplasmosis in amniotic fluid samples. Rev. da. Sociedade. Bras. Medicina. Trop. 46, 584–588. doi: 10.1590/0037-8682-0095-2013
CrossRef Full Text | Google Scholar
Uddin, A., Hossain, D., Ahsan, M. I., Atikuzzaman, M., Karim, M. R. (2021). Review on diagnosis and molecular characterization of Toxoplasma gondii in humans and animals. Trop. BioMed. 38, 511–539. doi: 10.47665/tb.38.4.091
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, Y., Wang, G., Zhang, D., Yin, H., Wang, M. (2013). Detection of acute toxoplasmosis in pigs using loop-mediated isothermal amplification and quantitative PCR. Korean. J. Parasitol. 51, 573–577. doi: 10.3347/kjp.2013.51.5.573
PubMed Abstract | CrossRef Full Text | Google Scholar
Ybanez, R. H. D., Ybanez, A. P., Nishikawa, Y. (2020). Review on the current trends of toxoplasmosis serodiagnosis in humans. Front. Cell Infect. Microbiol. 10. doi: 10.3389/fcimb.2020.00204
留言 (0)