
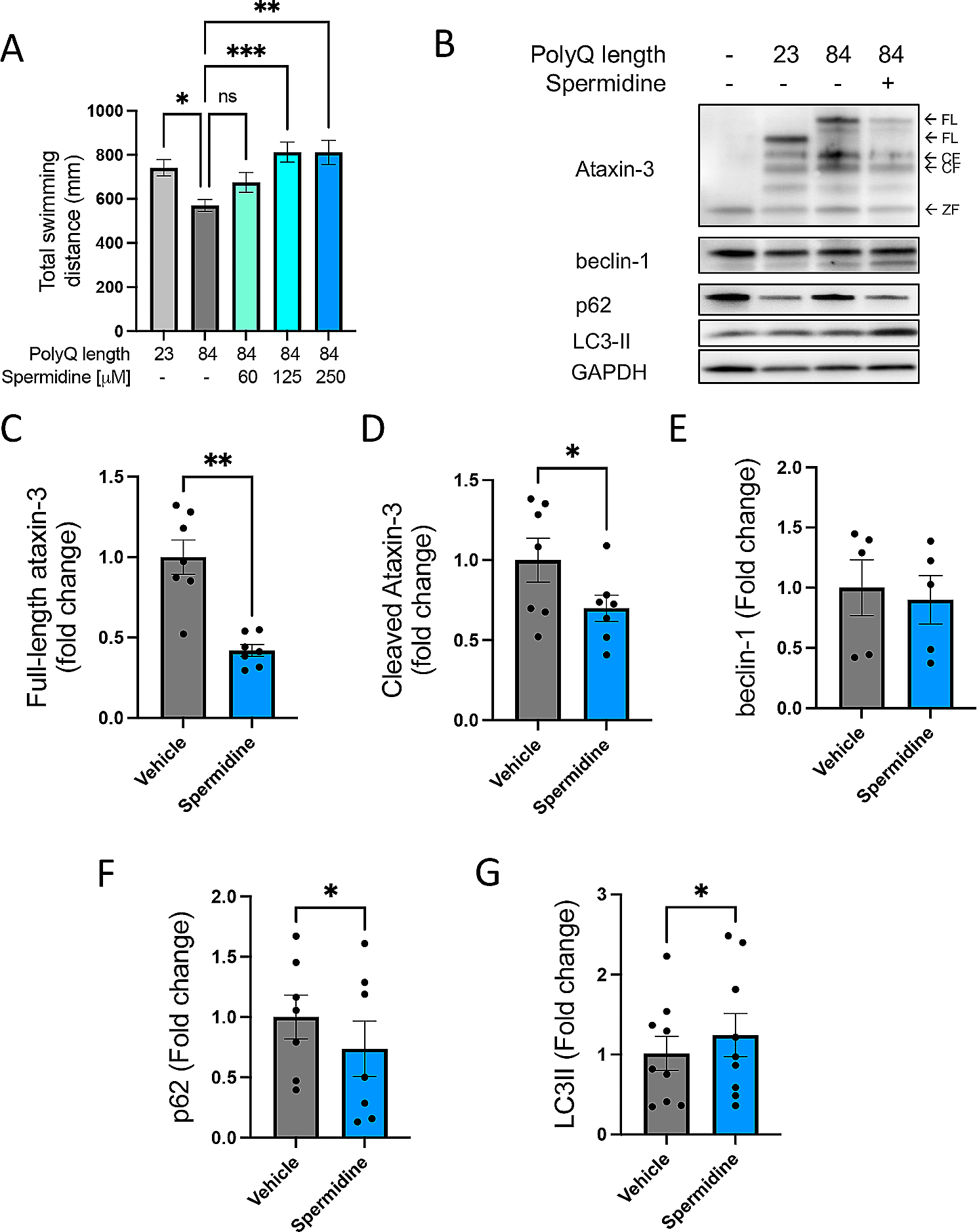
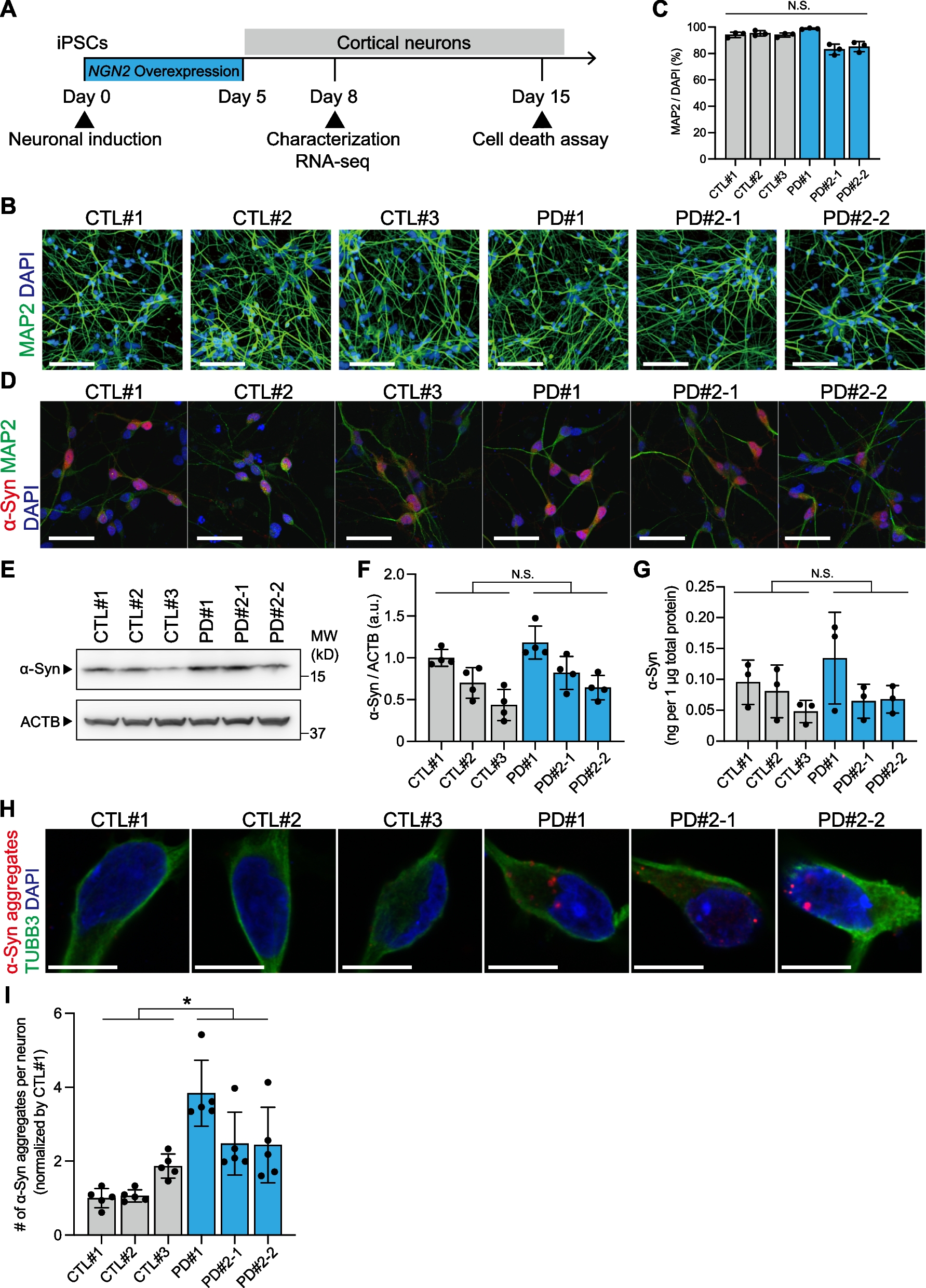
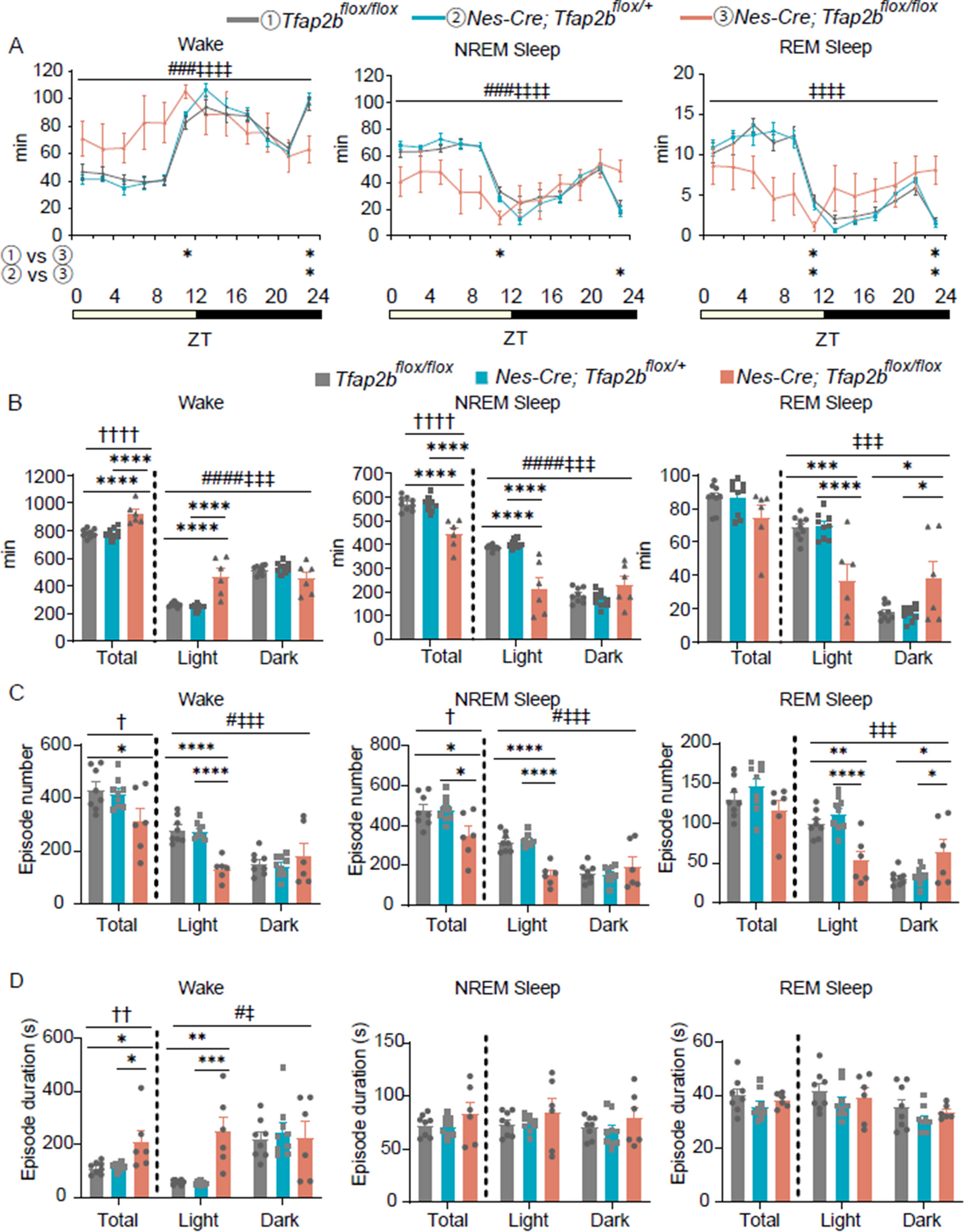
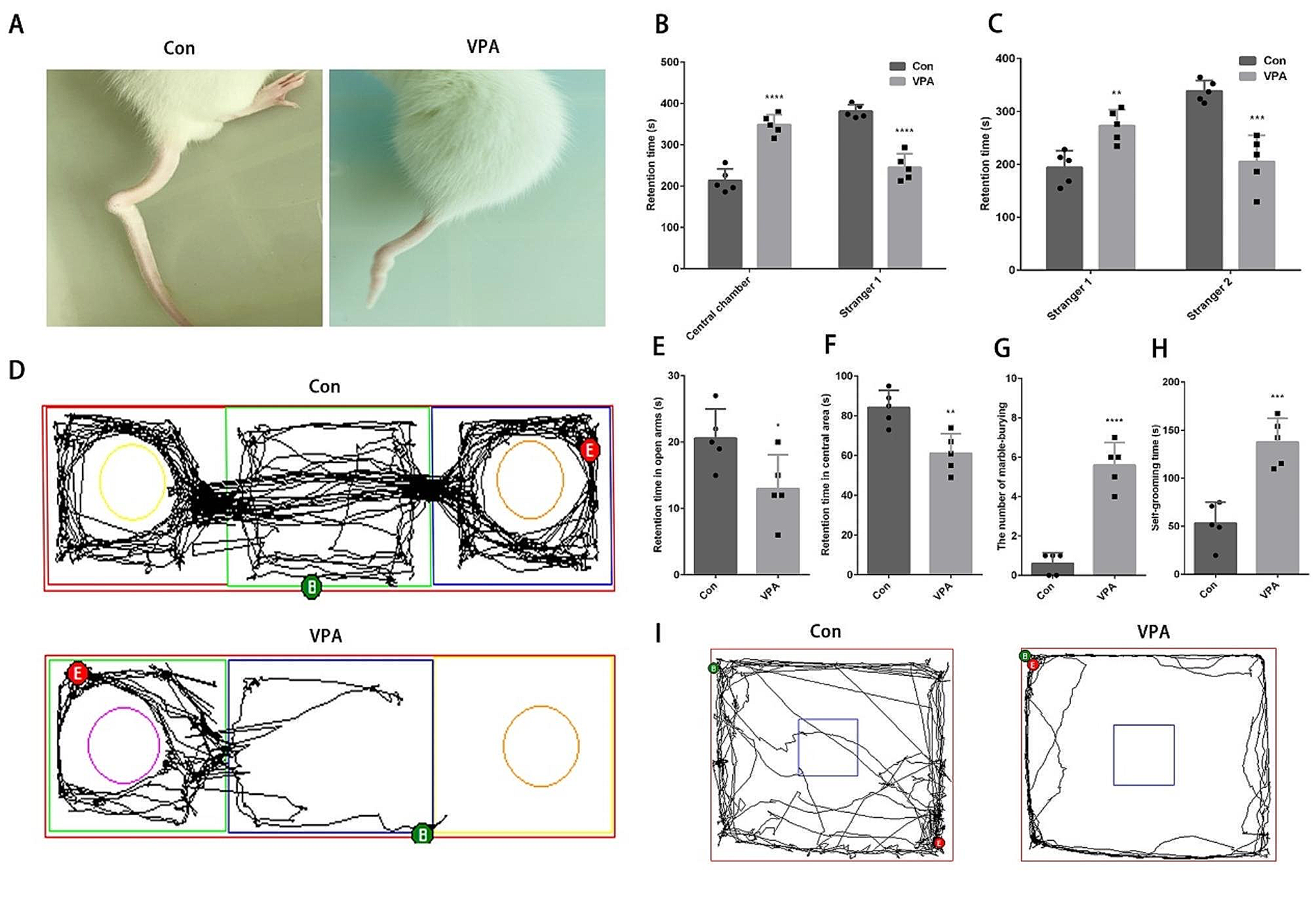
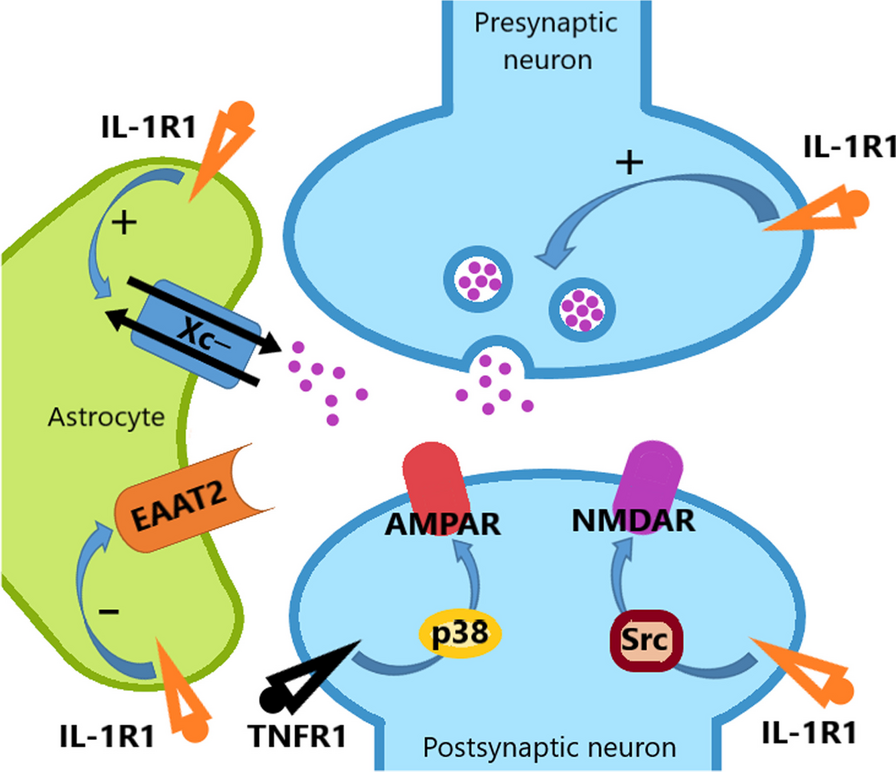
記住我




First, we established the specificity of puromycin labeling in the SUnSET assay [8, 13] of EYFP-expressing CA1 SOM-INs of acute hippocampal slices obtained from Sstires−Cre;Rosa26lsl−EYFP mice (SOM-EYFP-WT mice) [6] (Additional file 1—Materials and methods). Puromycin immunolabeling was present in approximately 25% of EYFP-expressing SOM-INs, but was absent in slices not treated with puromycin, or slices processed for SUnSET assay but without puromycin primary antibody (Additional file 2: Fig. S1). These results confirm the specificity of puromycin immunolabeling and indicate a detectable basal level of protein synthesis in SOM-INs of hippocampal slices.
Synaptic mechanisms that induce chemical late LTP in CA1 pyramidal cells (NMDAR, cAMP) [14] are different from the induction mechanisms implicated in chemical persistent LTP in CA1 SOM interneurons (mGluR1a) [6, 12]. So next, we examined if repeated mGluR1 chemical stimulation, an effective protocol for induction of mTORC1-mediated persistent LTP at PC-SOM synapses [6, 12], increases protein synthesis in SOM-INs of control SOM-EYFP-WT mice. After repeated application of the mGluR1/5 agonist (S)-3,5-dihydroxyphenylglycine (DHPG) in presence of the mGluR5 antagonist 2-methyl-6-(phenylethynyl)-pyridine (MPEP), puromycin immunolabeling was increased in SOM-INs relative to sham-treated slices (Fig. 1a), indicating that chemical induction of persistent LTP stimulates protein synthesis in SOM-INs. We tested the role of mTORC1 in persistent LTP-induced protein synthesis using mice with a conditional knockout of Raptor, an essential component of mTORC1, in SOM-INs (Sstires−Cre;Rosa26lsl−EYFP;Rptorfl/fl knock-out mice; SOM-EYFP-Raptor-KO mice) [6]. Repeated mGluR1 stimulation of slices from SOM-EYFP-Raptor-KO mice failed to increase puromycin immunolabeling in SOM-INs (Fig. 1a), indicating that protein synthesis elicited by induction of persistent LTP in SOM-INs is mediated by mTORC1 signaling. Basal level of puromycin immunofluorescence in SOM-INs of sham-treated slices was similar in SOM-EYFP-WT and SOM-EYFP-Raptor-KO mice (Additional file 2: Fig. S2a), suggesting undetectable mTORC1 control of basal protein synthesis in SOM-INs. We also examined puromycin immunofluorescence in CA1 stratum pyramidale after repeated mGluR1 stimulation. DHPG treatment did not increase stratum pyramidale puromycin immunofluorescence in slices of SOM-EYFP-WT or SOM-EYFP-Raptor-KO mice (Additional file 2: Fig. S2b-c), suggesting that induction protocol for persistent LTP at PC-SOM synapses stimulates protein synthesis in SOM-INs, but may not in PCs.
Fig. 1
Stimulation of protein synthesis in SOM-INs by chemical and optogenetic induction of persistent and transient LTP, respectively. a Protocol of chemical persistent LTP induction (top left), representative images (middle), cumulative distribution plots and summary bar graphs (bottom; each group 3 independent slice experiments from 3 animals, 3–6 sections analyzed per experiment) of puromycin immunofluorescence in SOM-INs of control SOM-EYFP-WT mice (left) and conditional SOM-EYFP-Raptor-KO mice (right), showing increase in fluorescence after DHPG treatment relative to Sham-treatment in control mice, but not in conditional knockout mice. Summary bar graph (mean ± SEM; SOM-EYFP-WT mice, Sham = 395 cells and DHPG = 396 cells; SOM-EYFP-Raptor KO mice, Sham = 457 cells and DHPG = 487 cells). b Protocol of optogenetic (TBSopto) transient LTP induction (top), representative images (middle), cumulative distribution plot and summary bar graph (bottom; each group 4 independent slice experiments from 2 animals, 4–6 sections analyzed per experiment) of puromycin immunofluorescence in SOM-INs of SOM-EYFP-WT mice, showing an increase in fluorescence after TBSopto relative to control (unstimulated) slices. Summary bar graph (mean ± SEM; Control 570 cells and TBSopto 761 cells). c Protocol of consecutive induction of TBSopto (in absence of puromycin) and repeated mGluR1 (in presence of puromycin) LTP (top), representative images (middle), cumulative distribution plot and summary bar graph (bottom; each group 6–8 independent slice experiments from 3–4 animals, 3–6 sections analyzed per experiment) of puromycin immunofluorescence in SOM-INs of SOM-EYFP-WT mice, showing that the increase in fluorescence after DHPG treatment is impaired by prior application of TBSopto. Summary bar graph (mean ± SEM; Ctl, Sham 781 cells and DHPG 805 cells; TBSopto, Sham 925 cells and DHPG: 869 cells). In all panels: arrows indicate cells with colocalization of EYFP with puromycin immunofluorescence; scale bars, 50 µm; Kolmogorov–Smirnov tests (cumulative distribution tests) or Student’s t-tests (group mean tests), * p < 0.05, ** p < 0.01, **** p < 0.0001 and ns not significant
Next, we determined if optogenetic theta-burst stimulation (TBSopto), an effective protocol to induce transient LTP at PC-SOM synapses [6, 9], increases protein synthesis in SOM-INs. hChR2 was expressed in PCs by CA1 injections of AAV2/9-CaMKIIa-hChR2(E123T/T159C)-mCherry in SOM-EYFP-WT mice, as previously [9] (Additional file 1—Materials and methods). Optogenetic theta burst stimulation (TBSopto) was then given as previously [9] in slices prepared for SUnSET assay (Fig. 1b). Puromycin immunolabeling was increased in SOM-INs after TBSopto relative to unstimulated slices from hChR2-expressing mice (Fig. 1b), indicating that optogenetic induction of transient LTP stimulates protein synthesis in SOM-INs.
Next, we examined if protein synthesis elicited in SOM-INs by TBSopto and repeated mGluR1 stimulation interact with each other. TBSopto was given to slices in absence of puromycin and repeated mGluR1 stimulation was applied 30 min later in presence of puromycin (Fig. 1c). In control slices (without prior TBSopto), puromycin immunofluorescence was increased in SOM-INs after DHPG treatment (Fig. 1c). But in slices with prior TBSopto, the increase in puromycin immunofluorescence after DHPG treatment was impaired (Fig. 1c). Thus, induction of transient LTP prevents further stimulation of protein synthesis by persistent LTP induction, suggesting both forms of LTP share signaling mechanisms.
留言 (0)