
記住我


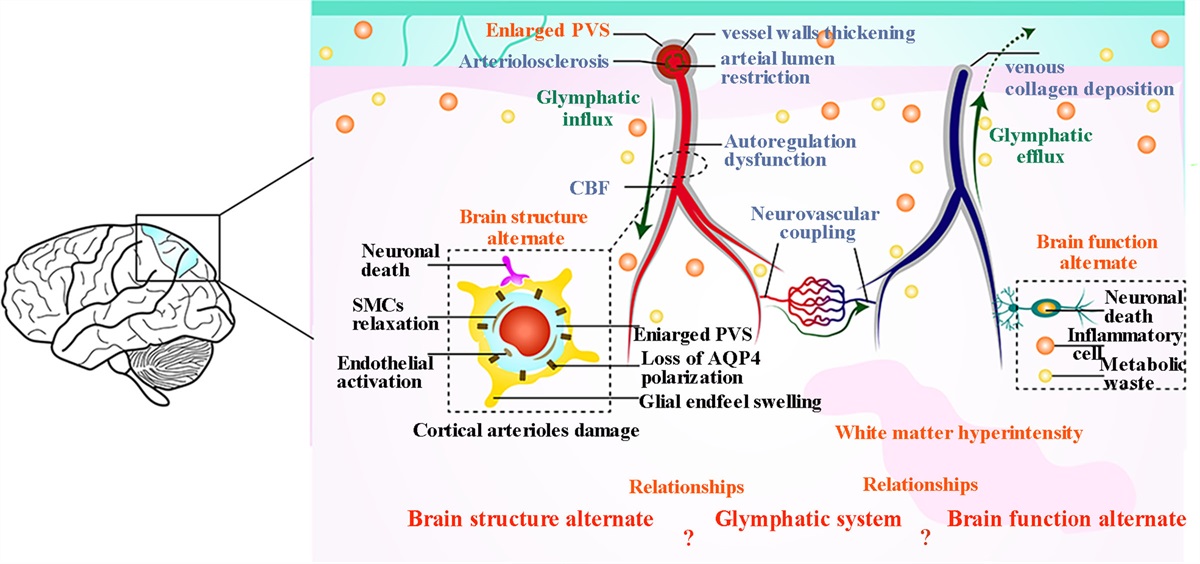
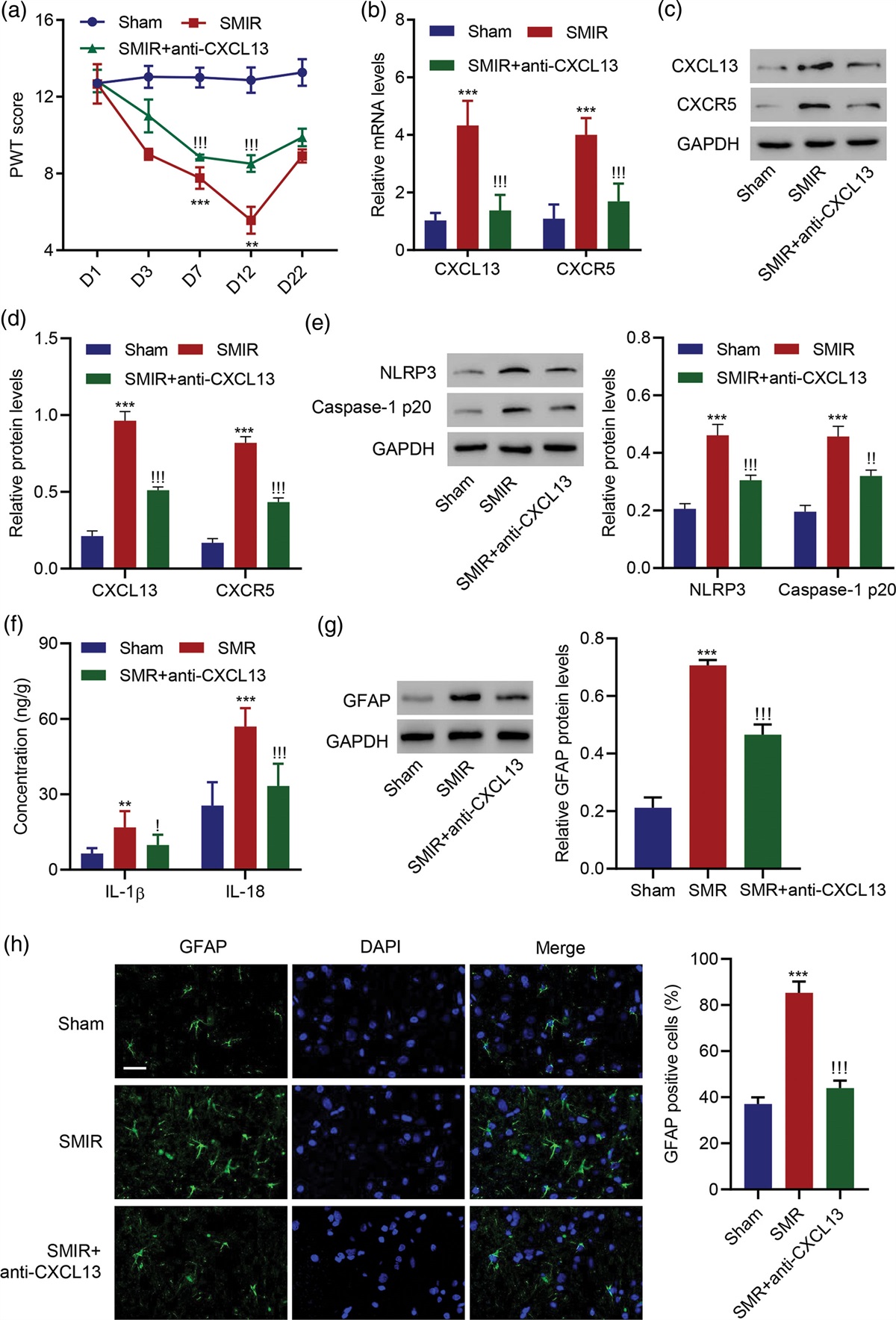
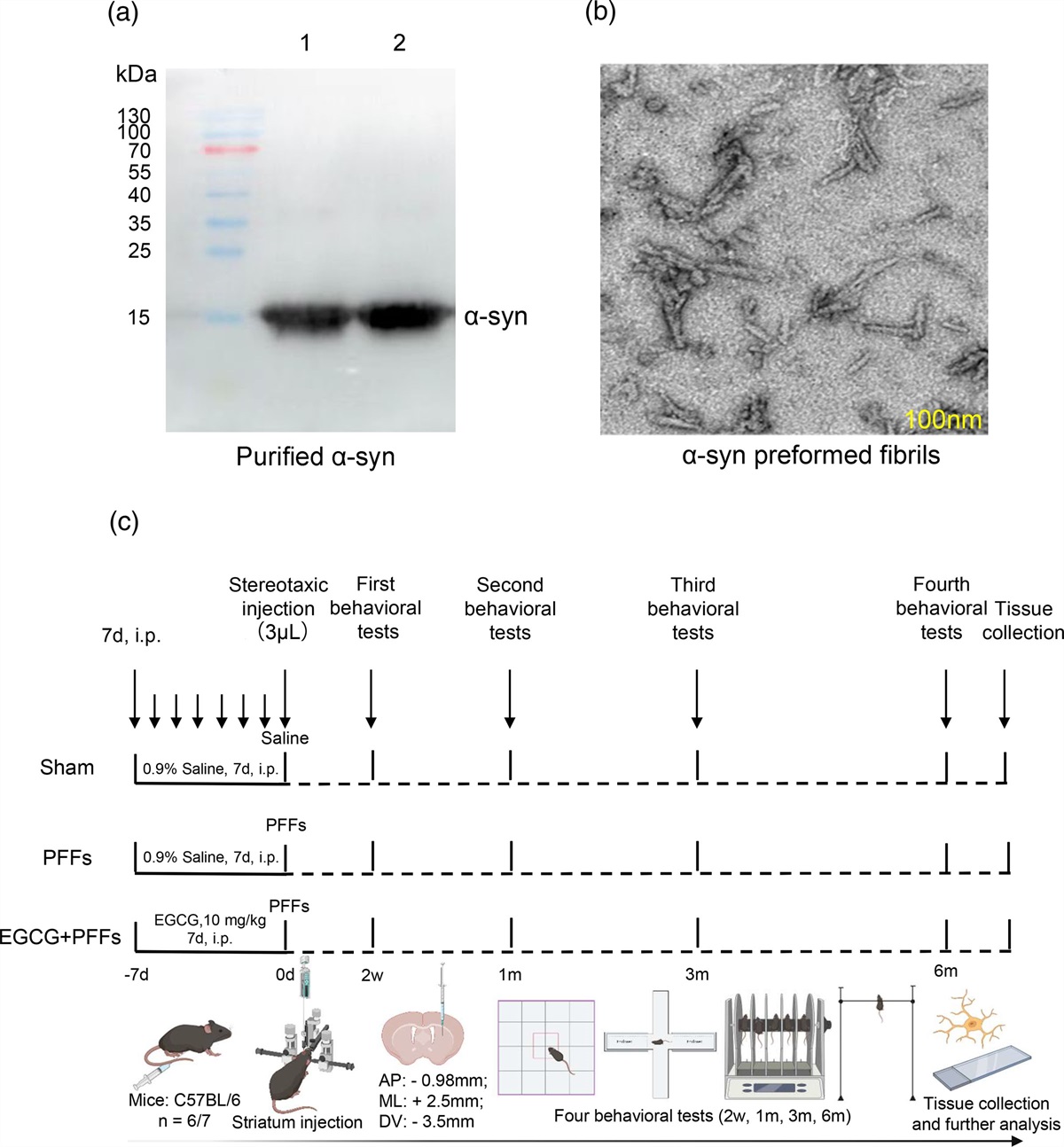
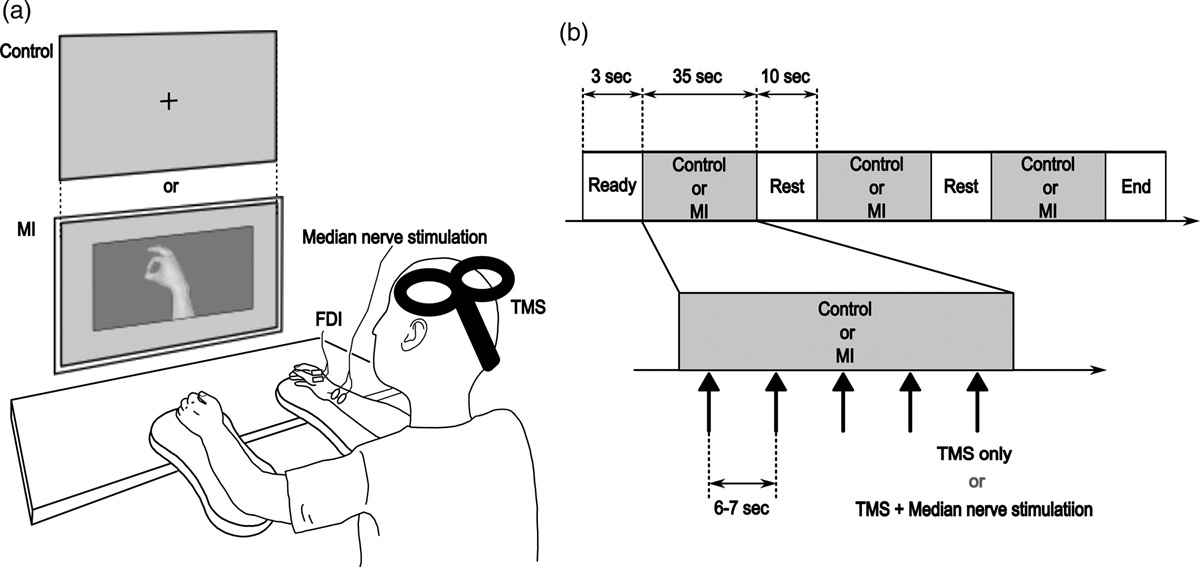
Stroke occurrences and mortality rates in the USA have declined in the past 30 years, but it remains a leading cause of long-term disability [1]. It is estimated that by 2030, around 3.4 million more people aged 18 years or older will experience a stroke [2]. Thrombolytic therapy, such as intravenous recombinant tissue plasminogen activator (tPA), has shown improved cognitive symptoms but must be administered within four and a half hours of the stroke [3]. Additionally, tPA is accompanied by an increased risk of intracerebral hemorrhage [4]. Ischemia-mediated cell death may be exacerbated by excitotoxicity, oxidative stress, free radical accumulation, mitochondrial dysfunction, impaired neurogenesis, angiogenesis, vasculogenesis and inflammation [5].
One potential form of therapy for stroke as a preemptive measure focuses on enhancing mitochondrial function. Preconditioning, exposing tissues to periods of ischemia for multiple, brief periods, can decrease the normal energy demand of the cell so that it may adapt to the state of oxygen and glucose deprivation (OGD) inherent in stroke. Cardiac studies have preconditioning that can drastically reduce the myocardial infarct size and irreversible injury caused by sustained or legitimate ischemia [6]. After preconditioning, rates of anaerobic glycolysis were decreased, ATP was preserved, and the cardiac insult remained focal rather than widespread [6]. In addition to the cardiac models of preconditioning, hypoxia has been shown to precondition neuronal progenitors [7]. By culturing human embryonic stem cell-derived neural progenitors in 0.1% oxygen, the cells were able to promote differentiation and survival to increase graft success [7]. Given that these neuronal stem cells can withstand hypoxic treatments, such preconditioned cells may be a good stem cell donor for transplantation in stroke.
Unfortunately, there is a heightened clinical risk of harming the patient when exposed to hypoxic preconditioning. Finding a noninvasive metabolic switching strategy may present as a well-tolerated and innovative approach to achieve preconditioning. By changing the energy source, the cell can be trained to utilize alternate fuel pathways and essentially induce mitochondria to survive in conditions with lower energy availability. In glucose-prevalent conditions, cells rely more heavily on glycolysis [8]. In OGD-like conditions, the cells cannot afford to deplete carbohydrate sources in glycolysis because it does not produce a large quantity of ATP. Thus, metabolism will be redirected to oxidative phosphorylation (OxPhos) [9].
A gap in knowledge exists about the efficacy and mechanism of enhancing OxPhos as a viable therapy for stroke. Thus, in the present study, we assessed the in-vitro mitochondrial modifications in cultured mesenchymal stem cells (MSCs) subjected to metabolic switching by altering the carbohydrate source from glucose to galactose, because cells grown in galactose media become more dependent on OxPhos and less on glycolysis [10]. Within the mitochondria, the Krebs cycle serves as a key step in OxPhos by transferring energy from the molecules produced by glycolysis to electron carriers, which are subsequently utilized in the electron transport chain to generate ATP. Most organisms use glucose for glycolysis and OxPhos to make maximum amounts of ATP. In contrast, galactose metabolism requires additional ATP to enter the glycolytic pathway and produces no net ATP in glycolysis [11]. Thus, it is possible this energetic compromise forces galactose-grown cells to rely heavily on OxPhos for energy that triggers ischemic tolerance. In humans, the galactose-based diet mimics a ketogenic diet [12], as a ketogenic diet minimizes glucose available for metabolism and ultimately shifts cells into oxidative metabolism [13]. The use of a ketogenic diet to modulate mitochondrial function has been successful in intractable epilepsy, inherited metabolic diseases and cancer [13]. In this study, we probed the underlying mechanisms of metabolic switching to gain insights on the therapeutic potential of mitochondria-based treatments in ischemic stroke.
Using the Agilent Seahorse extracellular flux (XF) Mito Stress Kit [14], we measured the oxygen consumption rate/extracellular acidification rate (OCR/ECAR) ratio. The OCR measures OxPhos, whereas the ECAR measures glycolysis. Thus, a higher ratio signifies greater oxidative metabolism [15]. This ratio is particularly important, as fine-tuning the homeostatic balance between these two energy metabolic pathways can impact angiogenesis and other cellular processes [16]. In this study, we hypothesized that metabolic switching could enhance the rate of OxPhos in MSCs, ultimately increasing the OCR/ECAR ratio and enhancing these cells’ therapeutic protection against stroke.
MethodsFirst, we cultured MSCs either under normal ambient cell culture condition (normal MSCs) or under the metabolic switching paradigm (switched MSCs). We assayed these two groups of MSCs for OCR and ECAR to reveal potential differences in their mitochondrial respiration and glycolysis. All investigators were blinded to the treatment conditions. Assignment of MSCs to either ambient or metabolic switching was conducted randomly.
Mesenchymal stem cells culturesHuman umbilical cords (hUC; n = 03) were purchased from Zen-Bio and they were obtained after mothers’ informed consent, immediately after full-term births with normal vaginal delivery. The isolation of MSCs from the perivascular region of hUC was performed as previously described [13]. An enzymatic method was chosen for the isolation of MSCs to increase the cell harvest yield around the vessels. Briefly, the hUCs were washed in 1× PBS to remove bloodstains and then rinsed in warm Hanks’ Balanced Salt Solution (HBSS) (Gibco) supplemented by 2× antibiotics/antimycotics (Gibco). Next, the hUC was cut into pieces of about 5–6 cm in length and then carefully sectioned longitudinally to expose the blood vessels. For the isolation of MSCs, the blood vessels were isolated using forceps and scalpel and placed in 40 ml of HBSS (Gibco) supplemented with 100 U/ml Type I Collagenase (Sigma, St Louis, Missouri, USA) and 0.01 U/ml Hyaluronidase (Stemcell Technologies) in a 50 ml Falcon tube and left to digest in for 4 h at 37 °C. After the digestion was completed, all the vessels were removed from the suspension using forceps. The suspension, containing the cells, was centrifuged at 285 g for 10 min. Subsequently, the supernatant was discarded and the cellular pellet was treated with 50 ml of 0.8% ammonium chloride (Stemcell Technologies, Vancouver, Canada) and incubated at room temperature for 5 min to lyse the erythrocytes. Thereafter, the tube was centrifuged for 10 min at 285 g and the supernatant was discarded. The cells, obtained from the perivascular region of each hUC, were counted and plated in one noncoated T-75 tissue culture flask with a complete medium and put it into a 5% CO2 incubator at 37 °C. We achieved confluence of MSCs at around day 14 in vitro, then we either continued the ambient cell culture condition or initiated the metabolic switching paradigm. The latter cell culture condition involved switching glucose (10 mM; 3 days) with galactose (10 mM; 3 days) in the media, which were alternately performed twice before processing the cells for the Seahorse mitochondrial respiration and glycolytic assay.
Mitochondrial respiration and glycolytic assayTo determine cellular OCR, the Seahorse extracellular flux analyzer XFe96 (102416; Agilent, Santa Clara, California, USA) was used in combination with sequential injection of various compounds. ECAR is measured simultaneously by quantifying lactate production as a measure of glycolysis [17]. OCR and ECAR measurements were performed following the manufacturer’s protocol. On the day of experiments, MSCs were detached from cell culture plates and seeded to a Seahorse 96-well plate (101085–004; Agilent) at 5.0 × 104 cells/well. Briefly, the Seahorse 96-well plate was centrifuged in a swing bucket rotator with slow acceleration (4 on a scale of 9) to a max speed of 450 rpm with 0 brake. Then, the plate orientation was reversed and centrifuged again to the max speed of 650 rpm with 0 brake. To determine OCR, the Seahorse extracellular flux analyzer XFe96 (102416; Agilent) was used in combination with sequential injection of various compounds [1 μmol/L oligomycin, 1 μmol/L carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP), 0.5 μmol/L Rotenone and Antimycin A] following the manufacturer’s protocol.
Extracellular flux analysis of cellular metabolic characteristics20 000 cells per well were plated in an XF24-well plate (Seahorse Bioscience, North Billerica, Massachusetts, USA). MSCs were then incubated for 45 min at 37 °C at ambient CO2 in HCO3-free Dulbecco’s Modified Eagle Medium (pH 7.4) containing 4 mM glutamine, 1 mM pyruvate and 10 mM glucose or 10 mM galactose. OCRs were determined in situ using a Seahorse Extracellular Flux Analyzer. Baseline oxygen consumption rate was measured 3 times for 4 min each separated by a 2 min wait and a 2 min mix. Following the measurement of basal respiration, oligomycin (600 ng/ml) (Sigma–Aldrich, Oakville, Ontario, Canada) was injected into each well, followed by three cycles of 2 min mix, 2 min wait and 4 min measurement to measure state 4 (nonphosphorylating) respiration. Then, FCCP (1 mM) (Sigma–Aldrich) was injected into each well, followed by three cycles of 2 min mix, 2 min wait and 2.5 min measurement to measure maximal respiration. Finally, antimycin A (4 mM) (Sigma–Aldrich) was injected into each well, followed by three cycles of 2 min mix, 2 min wait and 3 min measurement to measure extramitochondrial O2 consumption. The cells were then collected for the determination of protein content (Bradford assay). The data are presented as mitochondrial OCR (basal, oligomycin or FCCP OCR are reported following the subtraction of OCR in the presence of antimycin) and expressed per mg of total cellular protein. Finally, for the normalization, Hoechst 33342 solution was added in port D. Data were obtained using Wave software.
Statistical analysisThe data were evaluated using analysis of variance followed by post hoc Bonferroni tests. Statistical significance was preset at P < 0.05. Data are presented as mean ± SD.
ResultsTo measure the basal amount of OxPhos compared to glycolysis in switched MSCs and normal MSCs, the OCR/ECAR ratios at the first three timepoints of the mitochondrial respiration assay were analyzed (Figs. 1, 2) [14,18]. These three timepoints are of utmost clinical relevance, given that intrinsic MSCs preconditioned with metabolic switching would be in a basal energetic state. switched MSCs exhibited a significantly higher OCR/ECAR (P < 0.0001) at all three of these points compared to normal MSCs. Thus, metabolic switching effectively shifted mitochondrial metabolism from glycolysis towards OxPhos and greater energy production.
 Fig. 1:
Fig. 1: OCR/ECAR Ratio. Line chart shows the oxygen consumption rate (OCR) to extracellular acidification rate (ECAR) ratio changes in rate mode comparing MSCs grown under ambient cell culture condition (red) and MSCs exposed to metabolic switching condition (blue). MSCs, extracellular acidification rate.
 Fig. 2:
Fig. 2: Bioenergetic profiles. (a) Bar chart demonstrates the OCR rates (darker shade) and ECAR rates (lighter shade) at time point 0, basal metabolism, for 6 cultures of normal MSCs (left) and switched MSCs (right). (b) Scatter plot shows average OCR on the y-axis vs. ECAR on the x-axis of 6 cultures of normal MSCs (right) and switched MSCs (left). Points closer to the x-axis and further from the y-axis favor glycolysis, while points further from the x-axis and closer to the y-axis favor oxidative phosphorylation. ECAR, extracellular acidification rate; MSCs, extracellular acidification rate; OCR, oxygen consumption rat.
Once oligomycin was added, the ATP synthase protein was inhibited [14,18]. A cell line with a greater reliance on OxPhos would be impacted more by the addition of this inhibitor. This was reflected in our findings, as the OCR/ECAR ratio of the switched MSCs was significantly more decreased after oligomycin addition (time points 4–6) (time point 4 P < 0.001; time points 5 and 6 P < 0.0001) than that in the normal MSCs (Fig. 1). The addition of oligomycin further strengthened the argument that switched MSCs favored OxPhos more than normal MSCs.
In a similar fashion, FCCP uncoupled the proton gradient from ATP synthesis. When measuring OCR, uncoupling allowed for the measurement of spare respiratory capacity (SRC). SRC refers to the cell’s ability to respond to stressful situations and upregulate energy production [14,18]. The time points 7–9 showed that normal MSCs displayed a significantly greater OCR/ECAR than switched MSCs (P < 0.0001) after FCCP addition (Fig. 1) suggesting a greater SRC in normal MSCs than switched MSCs. These observations support the notion that the switched MSCs already harbored superior energy production at the basal period, as shown by time points 1–3.
Lastly, rotenone and antimycin A were injected into the assay to determine nonmitochondrial respiration. Rotenone inhibited complex I and antimycin A inhibited complex III, ultimately halting OxPhos [14,18]. Normal MSCs showed significantly greater OCR/ECAR after this third injection, measured at time points 10–12, compared to switched MSCs (P < 0.0001) (Fig. 1). These findings further lend support to switched MSCs’ greater reliance on OxPhos compared to normal MSCs, thus switched MSCs had less of a need for extraneous energy sources. Normal MSCs performed more glycolysis, a nonmitochondrial energy production process, also explaining the greater OCR/ECAR ratio.
Ultimately, the OCR/ECAR ratios observed in our mitochondrial stress assay suggest that metabolic switching of switched MSCs generated ‘super’ mitochondria capable of greater OCR compared to normal MSCs.
DiscussionThis study exemplifies the modulation of MSCs’ metabolism from a glycolytic to an oxidative state. While OCR and ECAR can be analyzed individually to measure OxPhos and glycolysis levels, the ratio of these two factors is important to understand the cells’ overall metabolic balance and functionality [16,19]. Using the Agilent Seahorse XF Mito Stress Kit [14], we demonstrated an increase in the OCR/ECAR ratio in switched MSCs compared to normal MSCs at baseline. By inhibiting ATP synthase with oligomycin, we showed that switched MSCs relied heavily on OxPhos for energy production than normal MSCs. Uncoupling the mitochondrial proton gradient from energy production, we illustrated that normal MSCs mounted a greater SRC under stressful conditions or increased energy demand. While we expected switched MSCs to be more efficient and more adaptable to stress via increased energy production, it is likely that these cells already harbored an elevated basal OCR/ECAR thereby lessening the need for SRC. Lastly, we observed that normal MSCs exhibited a higher OCR/ECAR after shutting down mitochondrial respiration, supporting that normal MSCs performed more glycolysis than switched MSCs. These findings support the use of metabolic switching to precondition MSCs as a prelude to cell transplantation in ischemic stroke. Furthermore, it advances the concept of enhancing OxPhos as a mechanism for mitigating ischemic stroke damage. The use of MSCs represents a model system for testing metabolic switching as a therapeutic approach to enhance the survival of stem cells, as well as intrinsic cells of the neurovascular unit within the stroke penumbra. Enhancing ATP production and reducing reactive oxidative species (ROS) may be induced pharmacologically via mitochondrial inhibitors [5] and antioxidant drugs [6] or physiologically through hypothermia [8] and exercise [20]. Stand-alone or combination treatments that replicate metabolic switching outcomes of efficient mitochondrial ATP production and ROS reduction pose as potent stroke therapeutics.
Although the OCR/ECAR ratio is commonly used to diagnose the Warburg effect in cancer cells [19,21], this study uniquely proposes adjusting the cells’ OCR/ECAR ratio as a treatment for stroke. Ultimately, this study may be relevant to caloric restriction to rewire neurons and precondition them to be more resilient to ischemia. This diet-modulated mechanism of preconditioning could be incredibly beneficial for individuals at a high risk of stroke, for example, a patient with pre- or postsurgical complications, fever, leukocytosis, hypertension, hypercoagulability or cardiac comorbidities [22]. Cognizant that stroke is an age-related disorder and mitochondrial functions deteriorate during aging, the incorporation of aging as a co-morbidity factor of a stroke may further define the role of metabolic switching in mitochondrial function [23,24]. Further research must be conducted to determine the optimal shifting paradigm to precondition MSCs. Additionally, when applied clinically, ideal diet conditions must be established to properly nourish the patient while also adjusting their cellular metabolism.
The present results highlight metabolic switching as a potential stroke treatment. In parallel, the use of MSCs in general offers yet another potential treatment, in particular, MSCs stand as potent donor stem cells for transplantation therapy in stroke [5,25]. However, generating more efficient mitochondria in MSCs before transplantation remains an unexplored adjunct therapy. Mitochondrial damage has been deemed a principal cause of ischemic cell death [26], with mitochondrial transfer from astrocytes to ischemic cells ameliorating stroke deficits [27]. MSCs possess the capacity to differentiate into multiple mesodermal lineages, which may exert therapeutic effects as immunoregulatory or trophic regulators following stroke. Because MSCs are not neurons, we acknowledge the limitation that MSCs may not fully recapitulate neuronal metabolism. Moreover, recognizing the complex response of multiple cell types within the neurovascular unit to stroke and the development of ischemic tolerance, further research is needed to translate our findings of metabolic switching in MSCs to the central nervous system setting. Nevertheless, the present study reveals that metabolic switching represents a robust approach in harnessing healthy mitochondria in MSCs. The amplified OCR/ECAR ratio in switched MSCs elevated the cells’ oxidative metabolism, which may represent a promising therapeutic strategy for stroke.
ConclusionMetabolic switching creates MSCs that harbor healthier and more efficient mitochondria amplifying their functional properties as transplantable stem cells in stroke. The increased OxPhos, as evidenced by the increased OCR/ECAR ratio, supports the use of metabolic switching for potent induction of ischemic tolerance.
AcknowledgementsM.M., J.-Y.L. and C.V.B. wrote the main manuscript text. A.G., H.N., V.C., J.-Y.L., and C.V.B. conducted experiments, performed data collection, and performed data analysis. M.M and J-Y.L. created the figures. All authors reviewed and approved the final manuscript.
All data is available as requested from the corresponding author C.V.B.
Conflicts of interestC.V.B. was funded by the National Institutes of Health (NIH) R01NS090962, NIH R01NS102395 and NIH R21NS109575. Additionally, C.V.B. was funded and received royalties and stock options from Astellas, Asterias, Sanbio, Athersys, KMPHC, and International Stem Cell Corporation and has also received consultant compensation from Chiesi Farmaceutici. C.V.B. also declares patents and patent applications related to stem cell therapy. For the remaining authors, there are no conflicts of interest.
References 1. Koton S, Schneider AL, Rosamond WD, Shahar E, Sang Y, Gottesman RF, Coresh J. Stroke incidence and mortality trends in US communities, 1987 to 2011. JAMA. 2014; 312:259–268. 2. Ovbiagele B, Goldstein LB, Higashida RT, Howard VJ, Johnston SC, Khavjou OA, et al.; American Heart Association Advocacy Coordinating Committee and Stroke Council. Forecasting the future of stroke in the United States: a policy statement from the American Heart Association and American Stroke Association. Stroke. 2013; 44:2361–2375. 3. Hacke W, Kaste M, Bluhmki E, Brozman M, Dávalos A, Guidetti D, et al.; ECASS Investigators. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. N Engl J Med. 2008; 359:1317–1329. 4. National Institute of Neurological, D. and P.A.S.S.G. Stroke rt. Tissue plasminogen activator for acute ischemic stroke. N Engl J Med. 1995; 333:1581–1587. 5. Anthony S, Cabantan D, Monsour M, Borlongan CV. Neuroinflammation, stem cells, and stroke. Stroke. 2022; 53:1460–1472. 6. Murry CE, Richard VJ, Reimer KA, Jennings RB. Ischemic preconditioning slows energy metabolism and delays ultrastructural damage during a sustained ischemic episode. Circ Res. 1990; 66:913–931. 7. Francis KR, Wei L. Human embryonic stem cell neural differentiation and enhanced cell survival promoted by hypoxic preconditioning. Cell Death Dis. 2010; 1:e22. 8. Zuurveld JG, Oosterhof A, Veerkamp JH, van Moerkerk HT. Oxidative metabolism of cultured human skeletal muscle cells in comparison with biopsy material. Biochim Biophys Acta. 1985; 844:1–8. 9. Redman LM, Ravussin E. Caloric restriction in humans: impact on physiological, psychological, and behavioral outcomes. Antioxid Redox Signal. 2011; 14:275–287. 10. Kase ET, Nikolić N, Bakke SS, Bogen KK, Aas V, Thoresen GH, Rustan AC. Remodeling of oxidative energy metabolism by galactose improves glucose handling and metabolic switching in human skeletal muscle cells. PLoS One. 2013; 8:e59972. 11. Conte F, van Buuringen N, Voermans NC, Lefeber DJ. Galactose in human metabolism, glycosylation and congenital metabolic diseases: time for a closer look. Biochim Biophys Acta Gen Subj. 2021; 1865:129898. 12. Qu C, Keijer J, Adjobo-Hermans MJW, van de Wal M, Schirris T, van Karnebeek C, et al. The ketogenic diet as a therapeutic intervention strategy in mitochondrial disease. Int J Biochem Cell Biol. 2021; 138:106050. 13. Vidali S, Aminzadeh S, Lambert B, Rutherford T, Sperl W, Kofler B, Feichtinger RG. Mitochondria: the ketogenic diet–a metabolism-based therapy. Int J Biochem Cell Biol. 2015; 63:55–59. 14. Agilent Technologies. Agilent Seahorse XF Cell Mito Stress Test Kit User Guide. Agilent Technologies, Inc. 2019. 15. Rogers GW, Throckmorton H, Burroughs SE. Gaining Insights into Disease Biology for Target Identification and Validation using Seahorse XF Technology. Agilent Technologies. 2019:1–9. 16. Yetkin-Arik B, Vogels IMC, Nowak-Sliwinska P, Weiss A, Houtkooper RH, Van Noorden CJF, et al. The role of glycolysis and mitochondrial respiration in the formation and functioning of endothelial tip cells during angiogenesis. Sci Rep. 2019; 9:12608. 17. Rose S, Frye RE, Slattery J, Wynne R, Tippett M, Pavliv O, et al. Oxidative stress induces mitochondrial dysfunction in a subset of autism lymphoblastoid cell lines in a well-matched case control cohort. PLoS One. 2014; 9:e85436. 18. Divakaruni AS, Paradyse A, Ferrick DA, Murphy AN, Jastroch M. Analysis and interpretation of microplate-based oxygen consumption and pH data. Methods Enzymol. 2014; 547:309–354. 19. Zhang J, Nuebel E, Wisidagama DR, Setoguchi K, Hong JS, Van Horn CM, et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nat Protoc. 2012; 7:1068–1085. 20. Fiorenza M, Lemminger AK, Marker M, Eibye K, Iaia FM, Bangsbo J, Hostrup M. High-intensity exercise training enhances mitochondrial oxidative phosphorylation efficiency in a temperature-dependent manner in human skeletal muscle: implications for exercise performance. FASEB J. 2019; 33:8976–8989. 21. Dar S, Chhina J, Mert I, Chitale D, Buekers T, Kaur H, et al. Bioenergetic adaptations in chemoresistant ovarian cancer cells. Sci Rep. 2017; 7:8760. 22. Cumbler E. In-hospital ischemic stroke. Neurohospitalist. 2015; 5:173–181. 23. Srivastava S. The mitochondrial basis of aging and age-related disorders. Genes (Basel). 2017; 8:E398. 24. Sun N, Youle RJ, Finkel T. The mitochondrial basis of aging. Mol Cell. 2016; 61:654–666. 25. Nguyen H, Lee JY, Sanberg PR, Napoli E, Borlongan CV. Eye opener in stroke. Stroke. 2019; 50:2197–2206. 26. Wang C, Youle RJ. The role of mitochondria in apoptosis*. Annu Rev Genet. 2009; 43:95–118. 27. Hayakawa K, Esposito E, Wang X, Terasaki Y, Liu Y, Xing C, et al. Transfer of mitochondria from astrocytes to neurons after stroke. Nature. 2016; 535:551–555.
留言 (0)