Despite being highly processive and inherently accurate, the genome duplication process in Escherichia coli or other prokaryotic cell is not intermittently completed without encountering impediments that can potentially pause the replisome unit.1, 2, 3 The progression of replication machinery is primarily hindered due to several considerable factors that include single or double-stranded breaks on the template, formation of unwanted secondary structures, or binding of undesirable proteins to the DNA near the replication site. However, these impairments are resolved through well-coordinated damage-specific repair mechanisms and that involves integrated action of various proteins especially helicases,[4] playing critical roles in rescue of the challenged replication fork.5, 6, 7, 8 Out of the various mechanisms, ‘recombinational repair pathway’ plays a noteworthy role in reassembly of the replication complex.[3] The recombination specific RecBCD complex generates single-stranded DNA through its nuclease/helicase activity and this is followed by strand invasion catalyzed by DNA strand exchange protein RecA.9, 10, 11 The nucleoprotein filament formed due to binding of RecA undergoes strand exchange with a homologous duplex giving rise to a Holliday junction, that in turn is resolved through RuvABC machinery.12, 13, 14, 15 Certain kinds of lesions bring replicating DNA unit carrying the infant strands in a mid-way halt, thus causing no net forward progression of the fork. Replication ‘fork reversal’ is a key protective mechanism that allows the forks to reverse their course when they encounter DNA lesions and resume DNA synthesis without chromosomal breakage.[16] The process of fork reversal could either be spontaneous[16] or catalyzed by a protein that finally leads towards the advancement of the fork in a direction opposite to replisome movement.17, 18, 19
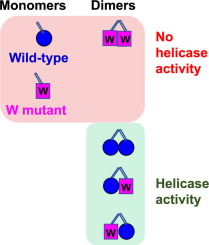
The RecG, in its monomeric form, is a key player in catalyzing the fork regression pathway and structurally modifies the stalled replication fork into Holliday junction through rearrangement of hydrogen bonds, thus making the halted fork a suitable target for recombination machinery.20, 21, 22 This works as a DNA translocase and unwinds a variety of branched DNA molecules in vitro, including Holliday junctions, D-loops, R-loops, and various models of replication forks.23, 24 It belongs to Superfamily 2 helicase (SFII helicase) and falls under the subfamily of ‘RecG-like helicases’[25], the family of proteins possessing several other properties in addition to its classical helicase activity of unwinding the double-stranded DNA. RecG comprises of three structural domains, viz. domain I, II, and III; the N-terminal domain I being the largest carrying more than half of the molecular weight of the protein.[26] The ‘wedge domain’, a highly conserved ‘greek key motif’ is present in domain I, is linked to domain II and III via an alpha-helical linker, and the other two domains are linked through a cleft. The protein binds to the three-way junction through domain I, with the wedge domain clamping RecG tightly onto the DNA to facilitate splitting of the junction and stabilizing the DNA-protein complex.[27] The domain II has an ATP binding site and it possesses helicase activity in conjugation with domain III. Acting as an atypical helicase, the protein translocates in a direction opposite to the movement of the replication machinery, which leads to the splitting of the newly replicated strands from the template DNA.[26] Being complementary to each other, the nascent leading and lagging strands anneal to form a four-way junction after getting separated from the respective parent strands. This four-way duplex intermediate structure is also famously known as ‘chicken foot intermediate’. Precise rearrangement of the chicken foot structure through branch migration leads to the development of Holliday junction by making it a suitable resolvase substrate.24, 28 It is not yet established and needs an appropriate justification that how RecG is guided towards a stalled replication fork with an abundance as low as 7 molecules per cell. However, it has been hypothesized that within the milieu of the genome, single-stranded binding protein SSB acts as a guiding unit to locate RecG at the site of the challenged replication fork for its rescue. 29, 30 Since, other than binding to single-stranded DNA and protecting them from degradation, SSB also interacts with a group of proteins for example PriA, RecQ, RecG, and Topoisomerase III, called ‘SSB-interactome’ facilitating their actions in genome modifications. Once loaded, the fork reversal activity of RecG has been hypothesized over the years through several bulk assays by determining the affinity and unwinding activity of RecG for various substrates resembling replication halted structures.21, 31 The reverse dynamics of the RecG through its course of rewinding the fork-DNA substrate has been studied employing magnetic and optical tweezers and it was verified that RecG catalyses the rewinding of the DNA substrate with high processivity.[32] It has also been found that RecG acts as a ‘stringent regulator’ for the rescue of stalled replication complexes and is capable of binding to a variety of fork structures with a difference in affinity and functional aspects. Acting as a helicase with 3’-5’ polarity, other than branched DNA substrates RecG has also been found to process negatively supercoiled DNA (scDNA), single-stranded DNA (ssDNA) or SSB coated M13 DNA. And, interestingly it has been verified that RecG has a stronger preference for negatively scDNA suggesting the phenomenon that substrate DNA should be converted from positive to negatively scDNA[33] for allowing RecG to function. Now, through deep functional analysis of every molecule via implementation of the high level of sensitivity and spatiotemporal resolution of the smFRET methods34, 35, we have closely monitored the action and quantified the dynamical heterogeneity of every possible arrested replication forked DNA upon being allowed to bind with RecG. We also documented the temporal kinetics in the unzipped and rezipped states of the substrate DNA while binding to RecG. Taking the same substrate fork and labelling it with donor and acceptor at different positions we could monitor the helicase action of RecG in both forward and reverse directions separately. By amalgamation of all the results we come up with a final model of ‘asymmetric fork regression’ where selectively lagging strand unwinding is catalyzed by RecG however processivity along the leading daughter strand is significantly less.






留言 (0)