
記住我
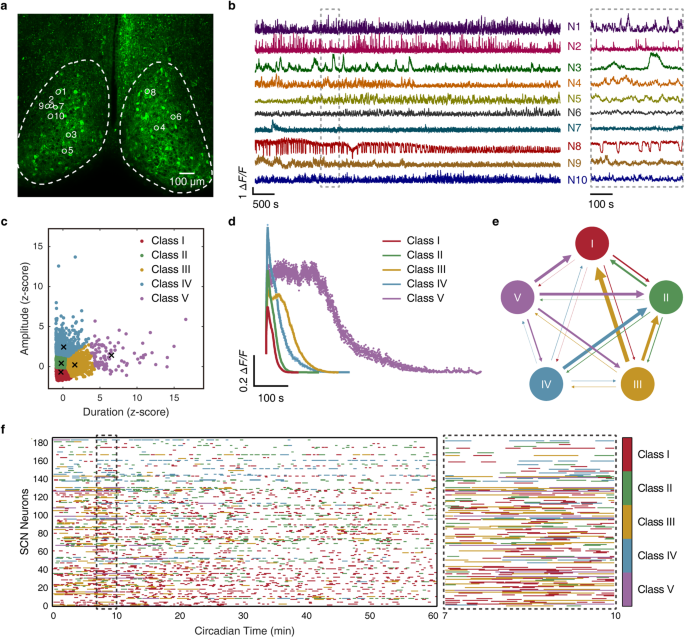

Three groups recently determined the cryo-EM structures of full-length transmembrane B cell receptor (BCR) in its resting state, representing a key advance toward our understanding of BCR signaling. These findings explain the molecular principles underlying BCR assembly and provide opportunities for structure-guided rational engineering of BCRs for the treatment of hematological and autoimmune diseases.
B cells play a central role in the adaptive immune response and provide durable protection against pathogens by differentiating into plasma cells that secrete pathogen-specific antibodies. B cell activation is triggered by antigen recognition by the B cell receptor (BCR), resulting in B cell proliferation and affinity maturation within germinal centers.1 Notably, B cell activation is critical for vaccine development and a molecular understanding of the role of B cells in autoimmune diseases.
The BCR is composed of homodimeric membrane-bound immunoglobulin (mIg) that confers antigen specificity (IgD and IgM expressed by mature naïve B cells; IgG, IgA and IgE expressed by memory B cells and plasma cells). mIg assembles with a disulfide-linked heterodimer formed by Igα and Igβ subunits that serves as a signal transduction platform.2 Each Igα and Igβ chain contains an Ig-like extracellular domain (ECD), a transmembrane (TM) domain and an intracellular immunoreceptor tyrosine-based activation motif (ITAM). Upon antigen binding, BCRs cluster in the plasma membrane, resulting in phosphorylation of ITAMs. The resulting signaling cascade drives B cell–T cell collaboration and differentiation of B cells into memory cells or antibody-producing plasma cells.
A key challenge in the field remains how antigen binding triggers BCR activation and signal transduction. Even though several models have been proposed on multivalent antigen binding inducing BCR oligomerization and activation,3 the underlying mechanism remains elusive due to lack of supportive structural information of the entire assembled BCR.4,5
The cryo-EM structures of human IgM-BCR and IgG-BCR reported by the Zhiwei Huang lab,6 together with that of human IgM-BCR by the Yigong Shi lab7 and murine IgM-BCR by the Hao Wu lab8 define the organization principles of BCR assembly. Notably, the Yigong Shi lab used IgM-BCR bound to Polatuzumab vedotin Fab fragment, an Igβ-targeting monoclonal antibody conjugated to an antimitotic agent that has been approved for the treatment of B-cell lymphomas.9
The three well-aligned IgM-BCR structures6,7,8 highlight the common architectural features required for assembly of the BCR complex characterized by a 1:1 stoichiometry of two mIg chains associated with Igα/Igβ subunits. The membrane-spanning pair of mIg chains adopt a Y-shaped topology, in which the Fab fragments are flexible whereas the Fc domains tightly pack with the ECDs of the Igα/Igβ heterodimer. The Ig-folds of Igα and Igβ are aligned in parallel with ~2-fold symmetry, with notable asymmetric alignment of Igα solely with the Fc domain of one mIg chain. Within the juxtamembrane region, the membrane-proximal connecting peptide (CP) of one mIg chain adopts a defined interdigitated topology with the Igα/Igβ heterodimer, resulting in a braided network of interactions that stabilize complex formation. Within the membrane-spanning region, the TM segments of the pair of mIg chains and those of the Igα/Igβ heterodimer form a compact four-helix bundle stabilized by hydrogen bonds involving conserved residues between the TM helices. By contrast, the cytoplasmic ITAM tails of Igα/Igβ are not traceable, suggesting that they are highly flexible. The observed interdigitated structural features strongly imply co-folding of mIg and Igα/Igβ in the assembly of the BCR complex.
In addition, the Zhiwei Huang group identified unexpected differences in ECD component alignments between IgM- and IgG-BCR complexes.6 The interaction between Fc and Ig-like domains adopt distinct “side-by-side” and “head-by-side” alignments for IgM-BCR and IgG-BCR, respectively. These differences could impact the threshold for signaling, highlighting the need for further structural investigation of other BCR subtypes.
The domain organization (Fig. 1a) and structure (Fig. 1b) of the BCR complex can be compared to its T cell counterpart, the T cell receptor (TCR)–CD3 complex (Fig. 1c, d) solved earlier by the Zhiwei Huang lab.10 In both receptor families, a dimeric receptor is responsible for ligand binding (peptide-MHC for TCR, antigen for BCR) while dimeric signaling modules with cytoplasmic ITAM motifs trigger cellular activation. While the BCR asymmetrically assembles with a single signaling dimer (Igα/Igβ), the TCR associates with three signaling dimers (CD3γε, CD3δε, ζ-ζ).11 As a consequence, the BCR has two ITAM motifs, while 10 ITAM motifs are present in the TCR–CD3 complex contributing to the high sensitivity of TCR signaling.
Fig. 1: Structure of the BCR and comparison with the structure of the TCR.
a Schematic drawings of domain architectures of the human BCR. Disulfide bonds are shown as yellow lines. b Two views of the overall structure of the BCR complex shown in a ribbon representation (PDB: 7XQ8). The Igα/Igβ subunits are highlighted in a surface representation. c Schematic drawings of domain architectures of the human TCR. d The structure of the TCR–CD3 complex shown in a ribbon representation (PDB: 6JXR). The TCR subunits are also highlighted in a surface representation.
The BCR forms a four-helix transmembrane bundle of mIg with Igα/Igβ while the TCR–CD3 complex forms an eight-helix bundle in which the TM helices of the three signaling dimers closely associate with the two TCR TM helices. A particularly interesting aspect of this comparison of BCR and TCR–CD3 is the importance of membrane-proximal CPs and the TM helices in ensuring specificity and stability of assembly. The importance of the TM interactions is supported by the greater sequence conservation of TM domains compared to ECDs. In the case of the BCR, conserved polar interactions are critical for assembly of mIg with Igα/Igβ. In the TCR–CD3 complex, each of the three positively charged amino acids of the TCR TM domains interacts with a pair of acidic TM residues of the interacting CD3γε, CD3δε and ζ-ζ dimers.12 Specificity of assembly is essential for both BCRs and TCR–CD3 because activation of these receptors is essential for all aspects of adaptive immunity conferred by B cells and T cells and initiates major transitions in cellular function. Further, the conserved nature of these intra-membrane and membrane-proximal interactions facilitates assembly of Igα/Igβ with five different classes of mIg molecules (IgD, IgM, IgG, IgA and IgE) as well as assembly of the three conserved CD3 dimers with TCRαβ or TCRγδ.
The reported structures of the IgM-BCR and IgG-BCR provide a structural platform for B cell signaling and set the stage for addressing major challenges in the field. Thus, which conformational changes in BCR result in ITAM signaling following antigen binding by the Fab domains? Notably, is there a role for BCR dimerization and/or oligomerization to trigger BCR signaling, and if so, which interfacial contacts mediate such clustering? Does the asymmetric BCR complex form specific interactions with co-stimulatory receptors during B cell signaling? Finally, how can this new structural information guide the rational design of small-molecule inhibitors or antibodies for the treatment of B cell-mediated hematological malignancies or autoimmune diseases?
留言 (0)