
記住我
The chemicals were obtained from Sigma Chem Co., unless otherwise stated. As a vertebrate model, the early development of chick embryos has been chosen. Early stages do not present bioethical concerns, as up to stage 15 HH, the neural crest is not yet differentiated to form sensorial ganglia and sense nerves. Fertilized eggs were obtained by a commercial hatchery (Ladi hatchery, Carasco, GE, Italy), brought to the laboratory and immediately put in incubator at 38.5 °C.
Cholinomimetic substances and inhibitorsAtropine (AT). Competitive nonselective antagonist at central and peripheral muscarinic acetylcholine receptors. A0132 Sigma-Aldrich; purity ≥ 99% (TLC), powder, water solubility: 2 mg/mL. Molecular weight 289.37; EC Number 200–104-8. Mother solution is stable for several days at 4 °C.
Carbamylcholine (Carbachol, CCh). Non-selective cholinergic agonist (analog of acetylcholine) that is resistant to the action of cholinesterases. C4382 Sigma-Aldrich, purity grade ≥ 98% (titration), crystalline. Molecular Weight 182.65; EC Number 200–127-3. Water solubility 1 g/ml.
Experimental procedureFor each experiment, 3 groups of 60 eggs were employed. The experiments were repeated at different seasonal times. The eggs were incubated flat and unturning before the treatment.
Some previous experiments were made by injection of the drugs into the sub germinal chamber to establish the final drug concentrations suitable to be used in this work.
Three microliters of different concentrations of the drugs, in Tyrode solution (TS, Tyrode 1910), or bovine serum albumin (BSA), was injected in the sub germinal chamber, using a Hamilton syringe. The concentration able to cause effects on each developmental event (LOEC) was identified as 10−4 M for AT and 10−3 M for CCh.
Staging of embryosControl embryos were staged according to the incubation time and the morphology, as described by HH (Hamburger and Hamilton 1951) and by the number of somite pairs (Fig. 8 in Appendix).
The treated embryos were staged according to the incubation time and compared with the control embryos incubated for the same time.
Exposure to the cholinomimetic drugs and spatial–temporal distributionFertilized eggs of white leghorn chicken were incubated at 38.5 °C in a controlled humidity incubator with 12 h circadian rhythm, for 20 and 24 h to obtain embryos at stages 4 and 6, respectively. The eggs were taken from the incubator and opened under a stereomicroscope in sterile condition, under a laminar flux hood. A window of about 1 cm diameter was made in the eggshell, corresponding to the position of the blastodisc, to reach the embryo surface.
A piece of agar, soaked with the test substances, was posed over the vitelline membrane, which is freely permeable to several substances dissolved in water, (Pons et al. 1985; Garcia et al. 1983; Rymen and Stockx 1974).
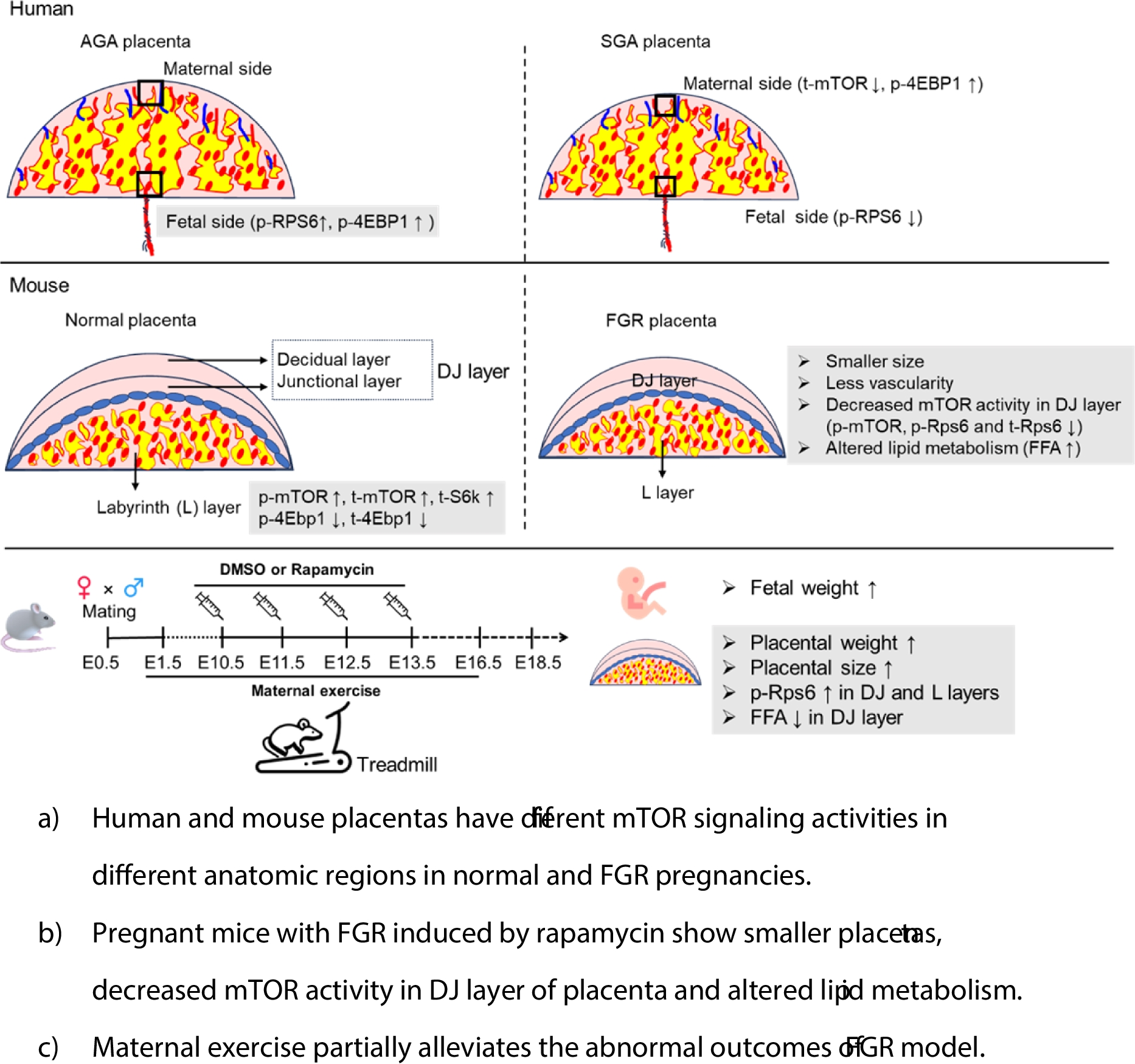
The agar was prepared by dissolving 20 g of Bacto Agar in 1 L TS, without stirring, in a thermostatic bath at 80 °C. After melting, the liquid was poured in a petri dish up to 1 mm high. After cooling and solidification, a parallelepiped of 2 × 2 × 3 mm was cut by a microscalpel (Moria, Paris) and soaked in 3 μL of 1 mM AT/TS or 2 μL of 0.1 M carbachol/BSA. The piece of agar was placed on the vitelline membrane at different distances from the opaque area. The position was cephalic or lateral or caudal to the embryo (see Fig. 1a). For controls, agar was soaked with sterile TS. With a pipette, the albumen was drawn to remove the liquid and free the surface of the membrane, so that the piece of agar could stick stably, then the shell was again filled with 1/9 albumen/TS, supplemented with 1% glucose. To maintain synchronic development, the eggs were taken simultaneously from the incubator, maintained outside along all the procedure (lasting some hours for 30 eggs), and put again into the incubator at the same time.
Fig. 1
Estimation of the position of embryo structures. a Scheme of a stage 4 HH, corresponding to time 0 (T0) of the experiment, the rectangles show the positions of the agar: (A) cephalic, (B) caudal; (C, D) lateral cephalic; (E, F) lateral caudal. b Theoretical growth of the control embryos during the experimental procedure: EA, exposure since 20 h incubation, EB, exposure since 24 h. The dotted line suggests the position of heart. This graph was used to extrapolate retrospectively the position of the different morphogenetic fields along time, to be compared with drugs distribution in space and time
After treatment, the egg window was sealed with the removed shell fragment, held on a piece of UV sterile cello-tape. The eggs were posed in the incubator and maintained unturned up to 33 and 50 h total incubation, respectively.
After the incubation, control and treated embryos were collected and rinsed in TS and the solution was drawn, till the embryos remained flat at the bottom of the Petri dish. Then, pre-cooled 3% paraformaldehyde (PFA) in TS was added drop by drop to cover the embryos. Fixation lasted 45 min at 4 °C, then the embryos were rinsed in different buffers, according to the subsequent procedures: in 0.1 M maleate buffer pH 6.0 for AChE activity localization; in PBS pH 7.4 for immunohistochemical staining. Staging of the embryos was made according to HH, the stage of exposed embryos was referred to the stage of the respective unexposed control embryos. The embryos stained for the revelation of AChE activity, were then dehydrated, mounted in resin, and observed under a Leitz microscope, equipped with a micrometric objective, to measure the embryos and control the distance from the agar.
Morphological and biochemical analysesHistochemical localization of AChE activityEmbryos, fixed in 2% PFA/TS for 15 min at 4 °C, were rinsed in 100 mM maleate buffer pH 6.0 and incubated in the medium suggested by Karnovsky and Roots (1964), containing 100 mM Na citrate, 30 mM Cu++ sulfate and 100 mM K+ ferricyanide in 100 mM maleate buffer, pH 6.0, 10 mg acetylthiocholine iodide (AcTChI, specifically hydrolyzed by AChE, and, with minor efficiency, by butyrylcholinesterase (BChE). The incubation was carried out in the dark at 4 °C overnight. Controls for the specificity of the reaction were performed by incubation without substrate or by pre-incubating for 30 min in a medium containing 1 μM eserine (Physostigmine), non-selective inhibitor of cholinesterases, of BW284c51 (anti-AChE, Burrough-Wellcome, USA).
Biochemical measurementAChE activity was measured by the quantitative method of Ellman et al. (1961), which was modified ad hoc for the Jenway spectrophotometer (6405 Jenway, Gransmore Green, UK).
Ten microliters of 0.5% Triton X-100 extracts was incubated in the presence of 50 μL of the substrates acetyl-β-metyl thiocholine iodide (AcMTChI) or butyrylthiocholine iodide (BuTChI) in phosphate buffer, pH 8.0, and stained by 50 μL of dithiobis-nitrobenzoic acid (DTNB). The reaction was allowed to develop for 10 min at room temperature and the absorption was measured at 412 nm and compared to a blank obtained by omitting the substrate. The enzyme activity was expressed in units = μmol of ACh hydrolyzed min−1 mgprotein−1. Each measurement was performed in triplicate and related to total protein content (Bradford protein assay, measured at 595 nm wavelength on the same spectrophotometer).
Immunohistochemical localization of α-tubulinsα-Tubulin labeling was used to detect the neurofilaments (Breuss et al. 2017) emerging from neural tube cells.
After fixation, the embryos were dehydrated with alcohol, clarified in xylol, and embedded in paraffin. Slices 5 μm thick with a Reichert-Jung microtome were made. Dewaxed slices were rinsed in PBS containing 0.5 M glycine, 1% serum albumin (BSA), and 5% goat serum albumin (GSA) and incubated for 2 h at room temperature in the primary antibody anti-αtubulin (Sigma), diluted 1:500 in PBS, 1%BSA, and 0.1%GSA. After rinsing in PBS, the secondary ab (FITC-conjugated anti-mouse immunoglobulin, Cappel, I) diluted 1: in PBS/BS/GSA was used for 2 h incubation in the dark. Specific controls were performed by omitting the primary antibody.
Mathematical modeling of the exposureThe transport by diffusion of AT and CCh in the aqueous solution was modeled, to compute at each instant the exposure of the embryonic segments and to establish the related interference in the inductive events taking place in the single embryo structures at each developmental stage.
The general governing equation for the three-dimensional diffusion equation of a chemical substance, subject to a first order reaction in homogeneous phase and linear reversible and instantaneous sorption, can be written as:
$$R\frac=_\frac^c}^}+_\frac^c}^}+_\frac^c}^}-\lambda c$$
(1)
where c is the solute concentration [M cm−3] in the considered control volume, x is the longitudinal coordinate, y and z are the horizontal transverse and the vertical coordinates, respectively, Dx is the longitudinal diffusion coefficient [cm2s−1], Dy and Dz are the horizontal transverse and the vertical transverse diffusion coefficients [cm2s−1], respectively, t is time [s], λ is the first order decay constant [s−1], and R is the retardation factor [-] related to sorption.
The retardation factor can be eliminated from the term on the left by replacing Dx, Dy, Dz with Dx/R, Dy/R, Dz/R, and λ with λ/R.
The initial condition can be expresses as follows:
$$c\left(x,y,z,0\right)=0$$
(2)
If we consider the source as a box of finite volume inserted into the domain, the following boundary conditions hold:
$$\beginc(\pm \infty ,y,z,t)=0& \mathrm& \frac_,t)}=0\end$$
(3)
$$\beginc(x,\pm \infty ,z,t)=0& \mathrm& \frac_,t)}=0\end$$
(4)
$$\beginc(x,y,\pm \infty ,t)=0& \mathrm& \frac_,t)}=0\end$$
(5)
where Lx, Ly, Lz [cm] are the borders of domain, supposed as finite or infinite, and the generation source term is expressed as:
$$r=\left\_f\left(t\right) ; 0<x<_; _<y<_ ; _<z<_\\ 0\end\right.$$
The general solution of Eq. (1) in infinite domain and (3), (4), and (5) expressed as first type boundary conditions on concentration can be formulated as follows:
$$\beginc(x,y,z,t)=\frac_}_^f(t-\tau )exp\left[-\lambda (\tau )\right]\left[erfc\frac_}_\tau }}-erfc\frac_\tau }}\right]\\ \left[erfc\frac_}_\tau }}-erfc\frac_}_\tau }}\right]\left[erfc\frac_}_\tau }}-erfc\frac_}_\tau }}\right] \, }\tau \end$$
(6)
This solution is valid for constant diffusion coefficients (Paladino et al. 2018).
To derive possible closed forms of this analytical solution, the boundary condition describing the source can be formulated as a Dirichlet (first type) or as a Newmann (second type) condition if a semi-finite x domain is considered. A Robin (third type) boundary condition cannot represent our experimental conditions since the velocity of the flux in input is negligible and anyway difficult to be evaluated (Massabo’ et al. 2011). With this approach, Eq. (3) can be reformulated as:
$$c(0,y,z,t)=\left\_f(t)\begin;& \end-D\frac=g(t)\\\ 0\end\right.\begin\begin-_<y<_;& -_<z<_\end\\ otherwise\end$$
(7)
$$\beginc(+\infty ,y,z,t)=0& \mathrm& \frac=0\end$$
(8)
where c0 [M cm−3] is the initial source concentration, f(t) is a dimensionless time function, and g(t) is a time function [M cm−3 s−1].
The main problem in modeling complex experiments involving drug release is the choice of reliable boundary conditions for the borders, i.e., well representing both the shape of the domain under study, and a good description of the source. To represent the experimental domain, the releasing source (the agar block) of dimension \(2_* 2_* 2_\) is set with the releasing face at z = 0, so to have a plane source generating the drug that diffuses in a quasi-2D domain to the exposed cells of the embryo (Fig. 9 in Appendix). A semi-finite x-domain is considered, and the releasing agar reduces to a linear source. Regarding the lateral boundary conditions in y-domain, since the embryo is smaller than the diffusion domain (germinal chamber) and it is generally in the middle of it, the hypothesis of infinite boundaries holds completely true.
The analytical solution becomes:
$$c(x,y,z,t)=\frac_x}_}}_^f(t-\tau )\mathrm\left[-\lambda \tau -\frac^}_\tau }\right]\left[erfc\frac_}_\tau }}-erfc\frac_}_\tau }}\right]\frac}\tau$$
(9)
Finally, as regards the description of the source release represented by \(f\left(t\right)\), the agar is a semi-solid (colloidal) basal medium, and the mechanisms of drug release from it are complex, including the diffusion of the chemical into the colloid, its dissolution, the dissolution of the medium, swelling and erosion processes by solvents (Rivadeneira et al. 2018). In swellable releasing devices, the chemical is dispersed into a glassy hydrophilic polymer and then compressed to form a solid. A gel-like phase is formed due to water swelling, and the bioactive agent is released, usually with a first-order release kinetics (Brazel and Peppas 2000). In swelling-controlled release systems, the chemical is dispersed into the gel-phase as films, disks, or spheres, and it is released at the interface, corresponding to the water penetration front. In this case, relaxation of the hydrogel influences the diffusion mechanism of the water-soluble chemical that remains immobile and begins its diffusion as the polymer swells with water. So, at initial times there is a moderated release, then the continued swelling of the matrix causes the drug to diffuse increasingly easily, prolonging and linearizing the release curve. The release mechanism could be described by different kinetics, depending on the diffusion regime inside the agar.
In this simulation, we neglected the initial transient of drug release, whose characteristic time is much smaller than the diffusion (exposure) time. Moreover, since at the end of the experiments some residual drug was still present inside the agar, we put \(f\left(t\right)\)=1 in Eq. (7), so concentration at the boundary in x = 0 is constant, and equal to the average volumetric concentration of the chemical inside the agar, taken as c0, and computed by knowing both the volume of the agar and the quantity of chemical injected into it during the experiments.
Moreover, we considered an isotropous medium inside the germination chamber, so the diffusion coefficients in x and y direction are both equal to the drug diffusion coefficient in water. The considered values of \(D\) are \(6\pm 3*^\) cm2 s−1 (Di Cagno et al. 2018). We must notice that the albumin surrounding the chick embryo is about 90% water and 10% proteins that are known for their strong drug binding activity. Somaratne et al. (2020) observed that egg white proteins can form a compact and microstructurally homogeneous gel at certain pH conditions, capable to reduce the diffusion coefficients of many chemicals. Conversely, the development of albumin-based drug delivery systems (Elsadek and Kratz 2012) suggests an opposite behavior for some particular drugs. The uncertainty here considered for the chosen diffusion coefficient is high, and could take into account these effects, even if they could be better described with a properly experimentally determined value of D in egg white. Another possible approach to consider the effect of albumin using the proposed model is to describe the bind as an adsorption mechanism and embed it into the retardation factor of Eq. (9), as proposed in (Peng et al. 2009), where an equilibrium reaction between the drug and the proteins is investigated.
Our proposed model is a deterministic physically based diffusion model, based on mass conservation principle. This means that the parameters inside the model have physical–chemical meaning, and they represent properties of the materials used in the experimental procedure. For this reason, it is not necessary to perform the model identification and the model validation steps to adopt it (Paladino et al. 2019). Anyway, even if parameters (i.e., the diffusion coefficient of the drugs inside the germination chamber) can be estimated in proper devoted diffusion experiments, in this case, the experimental punctual measurement of the concentration in time inside a germination chamber is not feasible (so synthetic egg white should be used). Therefore, model uncertainty exists; it is not due to the model shape, but it only derives from the errors in the chosen values of the diffusion coefficients, in this case estimated in water, and in the hypotheses embedded in the boundary conditions and describing the release mechanism.
Microscope images and spatial dimensioningThe embryos stained in toto for AChE activity revelation (60 exposed to AT, 60 exposed to CCh and 40 exposed to physiological solution) were fixed, mounted in resin, and images were taken on the light microscope (Leitz, DE). The final position of each embryo with respect to the agar was observed and classified as in Fig. 1a. The embryo images were georeferenced into the domain space. To do this, both the distance from the source and the embryo length at the final time of exposure were experimentally measured.
Statistical elaboration of the measures was performed to establish if the variation of the dimensions was due to the exposure or to individual variability.
留言 (0)