
記住我




Gadolinium-based contrast agents (GBCAs) are commonly used in contrast-enhanced magnetic resonance imaging (MRI) because of their ability to improve the visibility of internal body structures by increasing the relaxation rate of nearby water protons in vivo.1 The diagnostic application of GBCAs is currently considered largely safe in patients with normal renal function. However, after repeated administration of mainly linear GBCAs, retention of gadolinium (Gd) within different organs was reported to occur in patients without renal impairment.2,3 This resulted in restrictions on the use of linear GBCAs by the European Medicines Agency (EMA/625317/2017)4 and class warnings by the US Food and Drug Administration.5
Within central nervous system (CNS) structures, Kanda et al6 revealed for the first time a positive correlation between previous administrations of linear GBCAs and T1 hyperintensities in unenhanced brain MRI scans, in particular in the dentate nucleus (DN). These observations were confirmed by several other authors in humans7–12 and rodent models,13–19 principally after repeated application of linear GBCAs. Moreover, Gd retention in neuronal parenchyma, especially inside the DN, was quantitatively assessed by inductively coupled plasma mass spectrometry (ICP-MS) or laser ablation ICP-MS (LA-ICP-MS)17,20–26 after in vivo GBCA administration.
The risk of interstitial retention is especially high for the kinetically less stable linear GBCAs compared with macrocyclic agents.27,28 It is suggested that linear GBCAs partially dechelate in vivo, leading to transmetallation with endogenous cations such as calcium,29 iron,30 or zinc,31 resulting in long-term retention inside the brain. Contrary to that, macrocyclic GBCAs are expected to be washed out continuously in the form of the intact chelate,17,32 following specific clearance kinetics for the different macrocyclic compounds.33,34
In the context of chronic inflammation of the CNS such as in multiple sclerosis, GBCAs are primarily applied to determine the magnitude of blood-brain barrier (BBB) disruption and to estimate the number of active inflammatory lesions within the CNS.35 Hence, the concern for Gd retention is particularly relevant for patients with multiple sclerosis because GBCA-MRI is commonly used for initial diagnosis as well as for monitoring both clinical course and treatment response.36 Moreover, we previously reported using the experimental autoimmune encephalomyelitis (EAE) model that neuroinflammation may facilitate the retention of Gd in the mouse brains. We showed that 10 days after repeated gadopentetate injections, Gd levels were higher within inflamed brains compared with healthy controls (HCs) and demonstrated the formation of an elevated number of submicrometric Gd hotspots in the deep cerebellar nuclei (CN) of EAE mice.37
However, it has not yet been clarified for how long Gd may be retained in the inflamed CNS compared with healthy CNS, what factors may contribute to brain retention, how linear and macrocyclic GBCAs differ in terms of magnitude and kinetics of retention, and how Gd may affect neuronal tissue under inflammatory conditions. Therefore, we aimed to monitor and quantify Gd retention in brain tissue by MRI and LA-ICP-MS after repeated administration of either the linear gadopentetate or the macrocyclic gadobutrol in both healthy and EAE mice in vivo. Furthermore, using the well-established ex vivo model of organotypic hippocampal slice culture, we assessed Gd toxicity and how an inflammatory milieu may affect its toxic potential.
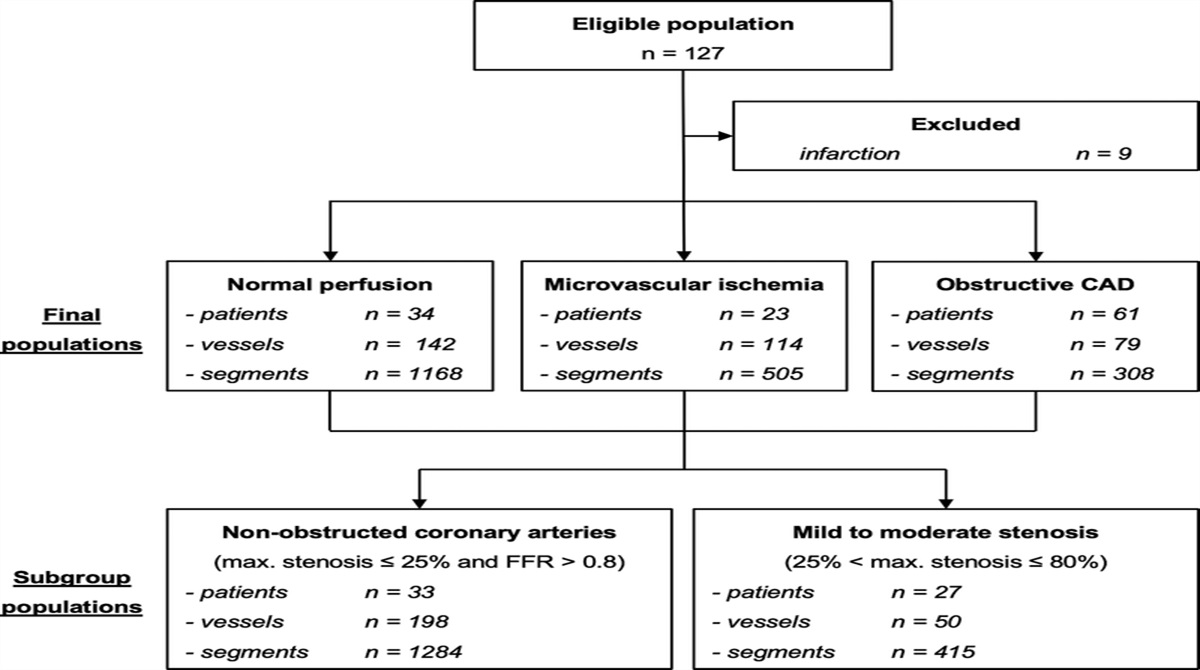
MATERIALS AND METHODS Mouse Model of EAE and Study DesignAnimal experiments were conducted in accordance with national and institutional guidelines for the care and use of laboratory animals and with directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 and were approved by the Berlin State Office for Health and Social Affairs (LAGeSo, registration number G106/19). Experimental autoimmune encephalomyelitis was actively induced in 9- to 12-week-old SJL/J female mice (Janvier Labs, France; n = 29) by subcutaneous immunization with 250 μg proteolipid protein peptide (PLP139–151; purity 95%; Pepceuticals, Leicester, United Kingdom) and 800 μg Mycobacterium tuberculosis H37Ra (Difco, Franklin Lakes, NJ) diluted in 100 μL complete Freund's adjuvant. In addition, 250 ng pertussis toxin (List, Biological Laboratories, Campbell, CA) were injected intraperitoneally on day 0 (day of immunization) and day 2 of the study. The EAE mice were monitored daily for clinical deficiency as follows: 0 = no disease; 1 = complete tail paralysis; 2 = hindlimb paresis; 3 = hindlimb plegia; 4 = paraplegia and forelimb weakness; and 5 = moribund or death. Experimental autoimmune encephalomyelitis (n = 29) and HC (n = 24) mice were exposed to 8 intravenous injections via the tail vein of either a linear GBCA (gadopentetate dimeglumine; Magnevist, Bayer, Germany, n = 27) or a macrocyclic GBCA (gadobutrol; Gadovist, Bayer, n = 26) at 2.5 mmol/kg body weight (BW), which corresponds to a cumulative dose of 20 mmol/kg BW. As displayed in Figure 1A, 4 consecutive daily injections of either gadopentetate or gadobutrol were followed by a 2-day break to reduce stress for the animals and 4 additional injection days as described previously.37 The GBCA applications started on day 12 to 13 postimmunization when EAE mice showed a peak of disability. The EAE mice presented first clinical signs on days 8 to 10, reached maximal clinical disability on day 12 to 13, and further showed a relapsing-remitting course until day 64 (Fig. 1B). No significant differences in EAE score between the time points of LA-ICP-MS, comparing the gadopentetate and gadobutrol groups, were detected (Kruskal-Wallis test; all EAE animals killed at respective time points [n = 4–5], P = 0.7509; EAE animals examined using LA-ICP-MS [n = 2], P = 0.3824).
 FIGURE 1:
FIGURE 1: Animal study design. A, Schematic illustration of the in vivo setup. A total of 24 HC and 29 EAE mice received 8 intravenous injections of either gadopentetate or gadobutrol at 2.5 mmol/kg BW (cumulative dose: 20 mmol/kg BW, 2-day pause after 4 consecutive applications). Mice underwent a baseline MRI before the first injection and further scans at day 1, 10, and 40 pfi of GBCAs. In EAE mice, baseline MRI started on day 12 to 13 postimmunization (peak of disease). After the MRI scans, 4 to 5 animals per group were sacrificed; brains were processed for LA-ICP-MS, and blood was collected for ICP-MS (n = 2/group). B, Clinical course of EAE animals based on the EAE score (mean ± SD). Red: gadopentetate-treated animals (n = 15), green: gadobutrol-treated animals (n = 14).
MRI and Image Data AnalysisWhole-brain coronal MRI scans were performed in vivo in EAE and HC mice on a 7 T small-animal scanner (Bruker PharmaScan, Ettlingen, Germany), running ParaVision 6.1 software at baseline (immediately before the first GBCA injection) and 1, 10, and 40 days post final injection (pfi) of either a linear or a macrocyclic GBCA. Measurements were conducted using a 72-mm linear volume coil for excitation (RAPID Biomedical GmbH, Rimpar, Germany) and a mouse head surface coil (Bruker, Ettlingen, Germany). T1 map image acquisition was conducted using an axial 2-dimensional rapid acquisition with relaxation enhancement at variable repetition times (RARE-VTR) sequence (echo time, 9.83 milliseconds; 8 repetition times from 255 to 7000 milliseconds; rare factor = 2; field of view, 19.2 mm2; matrix, 128 × 128; number of slices = 10; slice thickness, 1 mm; scan time, 17 minutes 13 seconds). During imaging, mice were anesthetized with 1.0% to 1.5% isoflurane in 30% O2 and 70% N2O administered via face mask, and gently fixed to the head coil to reduce motion and breathing artifacts. To maintain body temperature constant, animals were placed on a bed with circulating heated water. Respiration was monitored using a pressure-sensitive pad placed on the thorax (Small Animal Instruments Inc, Stony Brook, NY).
Magnetic resonance imaging data were registered to the Allen mouse brain atlas (ABA), as described elsewhere,38 in a blinded manner using an in-house developed MATLAB toolbox ANTx (available under https://github.com/ChariteExpMri/antx2). The T1 relaxation times of each single ABA brain structure were calculated using the back-transformed atlas, which matched the individual magnetic resonance images. The quantitative relaxometry assessment of Gd retention does not require any reference region. Differences in T1 relaxation time between baseline MRI and MRI at the different time points pfi of GBCAs were quantified as T1 relaxation time change (T1 change (%) = (T1pfi of GBCA – T1baseline)/T1baseline * 100). In animals with enlarged ventricles due to neuroinflammation, atlas transformation was corrected by subtracting the individual ventricle masks segmented on T2-weighted images using Analyze 10.0 (AnalyzeDirect, Inc).
Tissue Processing and HistologyMice were sacrificed with an overdose of ketamine/xylazine either 1, 10, or 40 days pfi of GBCAs (n = 4–5 mice/time point). Blood was collected and animals were immediately transcardially perfused with 0.1 M PBS at a continuous perfusion speed of 40 rpm (U/min; Ismatec ISM444B-115 V Analog Peristaltic Pump). Brains were extracted, fixed in 4% paraformaldehyde (PFA) and 30% sucrose, embedded and frozen (−80°C) for subsequent histological examinations. For histology, brains were cut coronally separating the forebrain, midbrain, and cerebellum. Frozen brain sections were cut in the coronal plane into 10-μm-thick cryosections and stored at −80°C. Consecutive frozen sections of those chosen for LA-ICP-MS were stained with hematoxylin and eosin (H&E) according to the standard protocol to assess inflammation. Hematoxylin and eosin images were acquired at ×2 and ×10 magnifications with a Keyence Fluorescence Microscope (BZ-X800, Germany) using the bright field mode.
LA-ICP-MS of Cerebellar SlicesThe LA-ICP-MS analysis of 10-μm-thick cerebellar cryosections was performed on a commercial LA system (NWR-213; ESI, Bozeman, MT) equipped with a 2-volume sample chamber coupled to a sector field ICP-MS (Element XR; Thermo Fisher Scientific, Bremen, Germany). A Y-piece was used to add argon to the helium carrier gas before entering the plasma. The ICP-MS was tuned daily for maximum ion intensity, signal stability (relative standard deviation <5%), and oxide ratio (ThO/Th <1%) during continuous ablation on a microscopic glass slide. Instrumental parameters of the ICP-MS system include radio frequency (RF) power: 1350 W; plasma gas flow (Ar): 16 L min−1; sample gas flow (Ar): 0.660 L min−1; auxiliary gas flow (Ar); 1.05 L min−1; mass resolution: 300 m/∆m; scanning mode: line by line; detected isotopes: 31P, 34S, 44Ca, 55Mn, 57Fe, 65Cu, 66Zn, 153Eu, 158Gd, and 160Gd. Parameters of the LA system include wavelength: 213 nm; helium gas flow: 1 L min−1; laser fluence: 1.9 J cm−2; spot size: 100 μm; scan speed: 100 μm s−1; repetition rate: 20 Hz; and line overlap: 20 μm. Data visualization was done in Origin 2018 (OriginLab Corporation, Northampton, MA). Calibration for 65Cu, 66Zn, and 158Gd (translation of count per spot of ablation [CPS] into fg/pixel) and drift correction was carried out by measuring matrix-matched standards cast on glass slides using analyte spiked agarose gels (analyte contents 0–310 fg/pixel).39 For further analysis of Gd retention in the whole cerebellar slice (mean), DN, and CN, manual regions of interest (ROIs) were applied to the calibrated LA-ICP-MS images of 158Gd. The analysis of each region was conducted 3 times in an independent randomized manner using ImageJ software40 for better reliability, and obtained Gd concentrations were averaged. Calibrated concentrations of 66Zn and 158Gd (fg/pixel) were translated into molar values (μM) where 1 pixel corresponded to 80 μm × 22 μm × 10 μm (1.76 × 10−11 L) of brain volume.
Analysis of Blood Samples by ICP-MSBlood samples were thawed and digested by a high-pressure asher (HPA-S, Anton Paar GmbH, Graz, Austria). In brief, 0.5 mL subboiled nitric acid (HNO3, 65%) was added to 100 mg of each blood sample and incubated overnight. Thereafter, samples were incinerated for 3 hours at 300°C. After cooling, the digested samples were diluted to a volume of 5 mL with MilliQ water. Samples were measured with an Element 2 ICP sector field mass spectrometer (Thermo Fisher Scientific) in combination with the autosampler 4DXF-73A (ESI Elemental Service & Instruments GmbH, Mainz, Germany). Calibration was carried out by standard addition of diluted multielement ICP standards. Blood control samples were obtained from an unmanipulated age- and sex-matched mouse. Instrumental parameters of the ICP-MS system for blood analysis include RF power: 1250 W; plasma gas flow (Ar): 16 L min−1; sample gas flow (Ar): 1.215 L min−1; auxiliary gas flow (Ar); 1.05 L min−1; mass resolution: 4000 m/∆m; detected isotopes: 55Mn, 115In, 155Gd, 156Gd, 157Gd, 158Gd, and 160Gd.
Generation of Chronic Organotypic Hippocampal Slices and Applied Culture ConditionsSJL/J pups (7–10 days) were decapitated, brains removed, and hippocampi extracted and sectioned coronally into 350-μm-thick slices using a McIlwain Tissue Chopper (Mickle Laboratory Engineering Co Ltd, UK). Six single slices from different mice were placed on 1 membrane of a cell culture insert (Millicell inserts, Merck, Germany) in a 6-well plate with 2 mL prewarmed (37°C) and oxygenized modified Opitz-Araya slice culture medium (SCM; 75% minimum essential medium (MEM) ×1, 20% normal horse serum heat-inactivated, 30 mM N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid (HEPES), 13 mM d-glucose, 5.2 mM NaHCO3, 2 mM MgSO4, 1 mM l-glutamine, 1 mM CaCl2, 1 mg/L insulin-transferrin-selenium supplement, 0.004% ascorbic acid, pen/strep) per well.41 Chronic slices were cultured for 12 days at 37°C and 5% CO2; SCM was changed every 2 days. From day 13 on, treatment with tumor necrosis factor α (TNF-α; mouse recombinant, lyophilized; Invitrogen) and the 2 different GBCAs (gadopentetate and gadobutrol) was initiated as displayed in Figure 2. To mimic inflammation, TNF-α at 50 ng/mL was added to half of the slices for a 48-hour preincubation period. On day 15, medium was exchanged and either gadopentetate or gadobutrol diluted in SCM was added on the top of the membranes at 1, 10, or 50 mM, respectively. In control cultures, no GBCA was applied. In addition, TNF-α at 50 ng/mL was added again to the corresponding slices for simultaneous coincubation with the GBCAs for an additional 48 hours. In positive control slices only, N-methyl-D-aspartic acid (NMDA) at 50 μM was added on day 16 for 4 hours to induce neurotoxicity.
 FIGURE 2:
FIGURE 2: Schematic illustration of the ex vivo study design. Organotypic hippocampal slices were incubated for 12 days after initial preparation. On day 13 treatment with TNF-α at 50 ng/mL was started in half of the slices (total incubation time: 96 hours). On day 15, treatment with either gadopentetate or gadobutrol was initiated (total incubation time: 48 hours), whereas simultaneous incubation with TNF-α at 50 ng/mL was continued. Incubation with NMDA for 4 hours was performed in positive control slices only on day 16. On day 17, the chronic cultures were terminated, followed by live staining with PI and further fixation in PFA and sucrose. Cell viability was assessed using fluorescence microscopy; IMC was conducted to visualize Gd tissue content.
Tissue Processing and Histology of the Chronic Hippocampal SlicesTreatment of slices with GBCAs ± TNF-α was stopped on day 17 by washing the membranes with fresh SCM. To assess cell death, propidium iodide (PI) was added to each well at 10 μg/mL for 30 minutes at 37°C. Further steps were performed without direct illumination to prevent photobleaching of the PI fluorophore. Subsequently, the membrane inserts were washed twice in prewarmed SCM for 10 minutes, followed by 1-hour fixation in 4% PFA. Thereafter, the membranes were transferred into sucrose (30%) in 0.1 M PBS for 2 days for further fixation. After fixation, slices were removed from the membranes and processed for cryosectioning. Cryostat slicing was performed at a thickness of 20 μm for assessment of the cell death ratio, at 10 μm for imaging mass cytometry (IMC) measurements. To analyze the cell death rate, hippocampal cryosections were counterstained with 4′,6-diamidin-2-phenylindol (DAPI) and subsequently mounted in a mounting medium (Thermo Fisher Scientific, Shandon Immu-Mount). Negative and positive controls were pooled for further analysis using fluorescence microscopy. Three independent rounds of experiments were conducted according to the experimental setup shown in Figure 2.
Fluorescence Microscopy and Toxicity AssayFluorescence images were taken from the section of the middle layer of slices and acquired using a Keyence Fluorescence Microscope (BZ-X800, Germany). Overview snapshots at ×2 magnification (CFI Plan Apo Lambda 2x; numerical aperture: 0.10, working distance: 8.50 mm) were taken in the DAPI filter line (BZ-X filter DAPI, model OP-87762; excitation wavelength: 360/40 nm, emission wavelength: 460/50 nm) for orientation. Thereupon, the dentate gyrus (DG) was imaged at ×20 magnification (CFI Plan Apo Lambda 20x; numerical aperture: 0.75, working distance: 1.00 mm). Only those DGs that passed the following criteria were imaged: (1) DG and hippocampal C1 region were distinguishable; (2) the original DG shape was maintained; and (3) neuronal cells of DG were in a good focus and distinguishable. Resulting DGs were imaged for DAPI fluorescence in the DAPI filter line and PI fluorescence in the Texas Red filter line (BZ-X filter TexasRed, model OP-87765; excitation wavelength: 560/40 nm, emission wavelength: 630/75 nm). Photobleaching was minimized by reducing laser power and exposure time between image acquisitions. Per treatment group 8 to 12 DG images were obtained. All images were blindly analyzed using ImageJ software.40 In stacks combining PI fluorescence and DAPI fluorescence images, manual ROIs around the single DG were drawn and the cell death rate was calculated (%, proportion of PI+ cells to DAPI+ cells). Recounting by independent investigators revealed a mean count-recount difference of 0.18% in cell death rate and a maximal count-recount difference of 1%.
Imaging Mass CytometryGadolinium-based contrast agent penetration within the chronic slices during the 48 hours of incubation was quantified by IMC of 10-μm-thick cryosections. Two untreated control sections, 2 sections treated with 10 mM gadopentetate, and 2 treated with 10 mM gadobutrol were ablated. Before the ablation process, nucleic acids were labeled with Iridium-Cell-IDTM Intercalator (125 μM; Fluidigm, Canada) according to IMC staining protocols (available at www.fluidigm.com) for subsequent colocalization of nucleic acids with elemental Gd. Imaging mass cytometry was performed on a CyTOF2/upgraded to Helios specifications coupled to a Hyperion Tissue Imager (Fluidigm, Canada), using CyTOF software version 7.0.8493. The instrument was tuned according to the manufacturer's instructions, using the 3-Element Full Coverage Tuning Slide (Fluidigm, Canada). The dried slide was loaded into the imaging module and ROIs were selected for each sample on a preview (panorama). Optimal laser power was determined for each sample to obtain complete ablation of the tissue. Laser ablation was performed at a resolution of 1 μm and a frequency of 200 Hz in a mass range of 75 to 209 Da. Data were stored as MCD files. For semiquantitative comparison of the elemental content of Gd inside the ablated hippocampal slices, the mean ion count of 6 ROIs per DG of defined size (100 μm × 100 μm) was calculated using the MCD Viewer v1.0.560.6. The mean ion counts of the 2 ablated hippocampal slices per treatment group were averaged.
Statistical AnalysisData were analyzed using the GraphPad Prism software (version 8.4.3; GraphPad Software, San Diego, California). Normal distribution of the variables was assessed using 3 normality tests (Shapiro-Wilk normality test, D’Agostino and Pearson normality test, and Anderson-Darling test); all data shown here were considered nonparametric. Values are expressed as geometric means and standard deviations. Two-group comparisons were performed by Mann-Whitney tests applying the Bonferroni correction for multiple testing over 3 consecutive time points. Because of the low sample size (n = 2 per group), merely descriptive statistics are shown for 158Gd and 66Zn concentrations measured using LA-ICP-MS and ICP-MS. As this was contemplated as an exploratory pilot study, no formal hypothesis testing was initially planned. Comparisons between more than 2 groups were accomplished by the Kruskal-Wallis test for nonparametric data followed by Dunn's post hoc test for multiple rank comparisons (α power = 0.05). Correlations among T1 relaxation times and the amount of detected Gd in cerebellar tissue were computed by the nonparametric Spearman rank correlation. P values <0.05 were indicative of significant group differences. The significance level of shown data was further displayed as * implying P < 0.05, ** implying P < 0.01, *** implying P < 0.001, and **** implying P < 0.0001.
RESULTS Inflammation Promotes Long-term Gd Retention After Gadopentetate but Not Gadobutrol ApplicationTo assess the effects of neuroinflammation on Gd retention after multiple injections of either linear gadopentetate or macrocyclic gadobutrol, T1 map RARE-VTR magnetic resonance images were acquired at day 1, 10, and 40 pfi (Fig. 3A). T1 relaxation time change (%) was calculated for the different brain areas that have been reported to retain Gd, including cerebellum (C), DN, CN, globus pallidus, thalamus, pons, medulla, hippocampus, and DG (supplementary Figure SDC 1, https://links.lww.com/RLI/A703). Whereas in HC animals, gadopentetate but not gadobutrol applications led to a negative T1 time change in the studied brain areas until 40 days pfi, in the EAE brain areas, both GBCAs appeared to be retained with similar regional distribution, although less pronounced for gadobutrol. Significant differences in T1 time change between inflamed and healthy brains at 24 hours pfi were observed for both linear gadopentetate and macrocyclic gadobutrol. Areas affected included the whole cerebellum, C, DN, CN, globus pallidus, thalamus, pons, medulla, hippocampus, and DG (supplementary Figure SDC 1, https://links.lww.com/RLI/A703).
 FIGURE 3:
FIGURE 3: 7 T MRI relaxometry in vivo. A, Representative longitudinal MRI scans (T1 map RARE-VTR sequence) of animals sacrificed 40 days pfi of GBCAs. Relaxometry revealed decreasing T1 relaxation times within the CN (red arrow) toward day 40 pfi of gadopentetate in EAE and HC animals. Gadobutrol-treated animals did not show prominent qualitative T1 shortening within the CN. B, T1 relaxation time change (%) differences between EAE and HC animals within the C, DN, and CN for both tested GBCAs. Mann-Whitney tests were computed for EAE versus HC after gadopentetate or gadobutrol treatment and the Bonferroni correction was applied for multiple testing over 3 consecutive time points. Data are displayed as mean ± SD.
Especially the cerebellar ROIs (C, DN, and CN) were identified as regions showing differences in T1 relaxation time change (%) between EAE and HC animals, as displayed in Figure 3B. Strikingly, differences between cerebellar ROIs of EAE and HC animals remained significant until 40 days pfi of gadopentetate (C, −10.61% ± 2.87% vs −2.76% ± 3.03%, P = 0.0477; DN, −15.13% ± 4.72% vs −3.64% ± 5.94%, P = 0.0477; CN, −13.69% ± 4.53% vs −2.64% ± 4.31%, P = 0.0477). Within the EAE group, T1 time change even increased when comparing 10 and 40 days pfi of gadopentetate, in particular within the DN and CN, suggesting long-term Gd retention within these areas. Contrary to that, as shown in Figure 3B, 10 and 40 days pfi of gadobutrol, no significant differences between EAE and HC ROIs regarding T1 changes were detected (C, −1.95% ± 3.30% vs 0.83% ± 1.67%; DN, 1.95% ± 4.03% vs 4.95% ± 7.75%; CN, −1.93% ± 3.96% vs −0.33% ± 2.89%), suggesting a continuous clearance of gadobutrol from the brain toward day 40 pfi also in the inflamed tissue.
LA-ICP-MS Confirmed Regional Patterns and Kinetics of Gd Retention in Inflamed Versus Healthy BrainsGadolinium retention inside the brain, as well as the distribution of different elements including zinc (Zn), copper (Cu), calcium (Ca), phosphorus (P), sulfur (S), manganese (Mn), and iron (Fe), was confirmed and quantified by LA-ICP-MS in representative 10-μm-thick cerebellar cryosections (n = 2 per group). Consecutive H&E-stained sections were imaged for localization of cerebellar ROIs (Fig. 4A).
 FIGURE 4:
FIGURE 4: Analysis of Gd distribution using LA-ICP-MS. A, On the left, H&E bright field image of cerebellar slice containing the CN corresponding to LA-ICP-MS tissue layer at ×2 magnification; CNs are additionally displayed at ×10 magnification. On the right, Allen brain atlas correlation image of CN formation. Nuclei from lateral to medial: DN, interposed nucleus (IP), vestibulocerebellar nucleus (VeCB), fastigial nucleus (FN). Scale bars: 1 mm. B, LA-ICP-MS images showing Gd distribution in cerebellar slices. Red areas display high and blue areas low Gd content (μM). Scale bars: 1 mm. C, Gd concentrations measured within the whole C, DN, or CN in μM of brain volume. Because of the low sample size, merely descriptive statistics are shown (mean ± SD).
Confirming MRI data, differences in Gd content between EAE and HC animals were obvious for both administered GBCAs (Fig. 4B). Whereas HC tissues retained Gd strictly limited to the areas of CN (including DN), the EAE cerebella presented Gd additionally localized within the granular layers of the cerebellar cortex, reaching mean Gd concentrations of 41.24 ± 6.97 μM on day 40 pfi of gadopentetate (Fig. 4B). In contrast, administration of gadobutrol led to a qualitatively much lower and diffuse Gd retention around the fourth ventricle (periventricular) and within the choroid plexus, as well as among marginal parts of the cerebellar cortex (Fig. 4B). The highest Gd levels were detected in brains of EAE animals receiving gadopentetate at all given time points of ablation. Gadolinium levels were 2- to 3-fold higher in EAE animals compared with HC mice at all measured time points, that is, 24 hours and 10 and 40 days pfi of gadopentetate, in particular inside the whole cerebellar slice (2.05–3.65-fold), the DN (1.66–2.03-fold), and CN (1.81–2.12-fold) (Fig. 4C). Interestingly, there was a trend of increasing Gd levels at day 40 pfi of gadopentetate in both EAE and HC mouse brains (EAE vs. HC; C, 11.51 ± 0.64 μM vs 3.15 ± 0.24 μM; DN, 49.03 ± 4.73 μM vs 29.46 ± 0.57 μM; CN, 55.06 ± 0.16 μM vs 30.44 ± 4.43 μM).
Regarding gadobutrol, Gd levels inside the C, DN, and CN were approximately 2- to 3-fold higher in EAE animals than HC mice 24 hours pfi. These levels were reduced at day 10 within all assessed brain areas but still higher in EAE. On day 40 pfi, Gd levels diminished to less than 0.40 μM regarding both EAE and HC animals as shown in Figure 4C (EAE vs HC: C, 0.28 ± 0.03 μM vs 0.14 ± 0.04 μM; DN, 0.37 ± 0.08 μM vs 0.17 ± 0.04 μM; CN, 0.38 ± 0.08 μM vs 0.17 ± 0.03 μM).
In particular, in the inflamed CN, administrations of gadopentetate resulted in retention of Gd 24 hours pfi approximately 4-fold higher compared with gadobutrol (DN, 4.43-fold; CN, 4.13-fold) and 7 to 8-fold higher regarding HC (DN, 8.53-fold; CN, 7.14-fold) (Fig. 4C). Differences between both GBCAs became more apparent on day 40, as Gd amounts pfi of gadopentetate were even increased, in contrast to strongly reduced Gd concentrations found after gadobutrol administration (Fig. 4C).
In addition, as shown in the supplementary Figure SDC 2A, https://links.lww.com/RLI/A704, we demonstrated an inverse correlation of Gd concentrations and mean T1 relaxation times for all 3 analyzed cerebellar ROIs, that is C (rs = −0.93, P < 0.0001), DN (rs = −0.74, P = 0.0078), and CN (rs = −0.78, P = 0.0043) (see Supplemental Digital Content, Figure SDC 2A, https://links.lww.com/RLI/A704).
Gd Was Detectable in Blood Samples 40 Days pfi of Both Tested GBCAsTo evaluate the potential mobilization of Gd from organs to the blood circulation during the observation period, Gd concentrations (μg/g blood) in blood samples of EAE mice (n = 2/time point) and HC mice (n = 2/time point) were assessed by ICP-MS. Elevated Gd levels were detected in all blood samples at all analyzed time points pfi of both GBCAs, gadopentetate and gadobutrol (supplementary Figure SDC 2B, https://links.lww.com/RLI/A704). In line with Gd content within cerebellar brain slices assessed using LA-ICP-MS, EAE animals showed elevated Gd levels compared with HC animals (except at day 10 pfi of gadopentetate). Maximal Gd concentrations detected corresponded to 0.0054% of the intravenously injected cumulative dose of 20 mmol/kg BW (24 hours pfi of gadopentetate in EAE mice). A negative control sample (untreated age- and sex-matched mouse) demonstrated a very low Gd concentration of 0.0002 μg/g, as shown in the supplementary Figure SDC 2B, https://links.lww.com/RLI/A704. Because of the low sample size (n = 2 per group; HC at day 1 pfi of gadopentetate: n = 1 due to sample contamination) and high variation, only descriptive statistics were performed (see Supplemental Digital Content, Figure SDC 2B, https://links.lww.com/RLI/A704).
LA-ICP-MS Revealed Elevated Zinc Content in Inflamed Mouse BrainsNext, we aimed to identify endogenous metals whose distribution may be affected by inflammation and could be related to Gd retention within cerebellar structures. Interestingly, elevated Zn levels were found in cerebellar slices of EAE animals compared with HC mice at all time points, irrespective of the applied GBCA (Fig. 5A, B). Zn was diffusely distributed in the EAE cerebellar sections at 24 hours pfi of GBCAs, whereas 10 and 40 days pfi, high Zn levels were predominantly located within and around the CN within the white matter and layers of the cerebellar cortex (Fig. 5A). Furthermore, cerebellar EAE slices, as well as HC slices, revealed a decreasing trend in Zn levels toward day 40 pfi. This trend was particularly pronounced in the inflamed DN and CN, irrespective of the applied GBCA, as shown in Figure 5B. Amounts of Zn in EAE brains regarding the C, DN, and CN decreased by 25%, 26%, and 26%, respectively, whereas in HC brains, Zn levels decreased by 14%, 21%, and 22%, respectively, toward day 40 pfi (Fig. 5B).
 FIGURE 5:
FIGURE 5: Analysis of elemental colocalization using LA-ICP-MS. A, Laser ablation images of cerebellar Zn distribution. Red areas display high and blue areas low Zn content (μM). The EAE animals in both GBCA groups showed qualitatively higher Zn levels compared with HC. Scale bars: 1 mm. B, Demonstration of Zn levels within the whole C, the DN, and CN (μM). The EAE animals consistently presented higher elemental Zn levels than HC animals irrespective of the applied GBCA with a tendency to decrease toward day 40 pfi of GBCAs.
The overall distribution of Cu, Ca, P, S, Mn, and Fe concentrations seemed to be unaffected by inflammation (supplementary Figure SDC 3, https://links.lww.com/RLI/A705). However, areas with high Gd content overlapped with areas of high Fe levels (see Supplemental Digital Content, Figure SDC 3, https://links.lww.com/RLI/A705).
Gadopentetate but Not Gadobutrol Exerts a Neurotoxic Effect on Living Organotypic Hippocampal SlicesTo assess the potential toxicity of GBCAs, we measured the cell death rate (%) after 48-hour treatment of chronic hippocampal slices with gadopentetate or gadobutrol at 1, 10, and 50 mM, respectively (Fig. 6). To simulate neuroinflammatory conditions, TNF-α at 50 ng/mL was added to half of the slices. Gadopentetate promoted cell death within the DG only at 50 mM (Fig. 6A, B). No toxicity was observed for gadobutrol. Tumor necrosis factor α at 50 ng/mL enhanced the gadopentetate-induced cell death rate significantly (with vs without TNF-α: 3.15% ± 01.18% vs 2.17% ± 1.14%; P = 0.0345), as shown in Figure 6B. NMDA incubation at 50 μM served as a positive control. Its neurotoxic effect was also increased by TNF-α (34.34% ± 11.14% vs 19.70% ± 9.19%; P = 0.0002) (Fig. 6B).
 FIGURE 6:
FIGURE 6: Toxicity of GBCAs on organotypic hippocampal slices. A, Representative fluorescence images of the DG; all nuclei in blue (DAPI), dead nuclei in red (PI). The DGs of slices coincubated with/without TNF-α (50 ng/mL) are displayed for all experimental conditions. The PI-positive cells appear in NMDA-treated (±TNF-α) and gadopentetate-treated slices at 50 mM (±TNF-α). Scale bars: 200 μm. B, Cell death rate (%) was elevated in NMDA (positive control)-treated slices and slices treated with 50 mM gadopentetate. Tumor necrosis factor α enhanced cell death rate significantly. Data were evaluated using Mann-Whitney tests (mean ± SD). C, IMC images of 10-μm-thick slices after gadopentetate or gadobutrol treatment at 10 mM and negative control. The DGs were surrounded by manually drawn ROIs (white), and thresholds were adjusted for optimal visualization. Scale bars: 200 μm.
To examine whether the lack of cytotoxicity of GBCAs at especially lower concentrations was caused by deficient penetration of the compounds into the living slices, IMC was performed using slices treated with 10 mM of GBCAs. Gadolinium was detectable inside hippocampal slices after treatment with both GBCAs, gadopentetate and gadobutrol (Fig. 6C). However, the mean 158Gd count inside the DG region was 26 times higher in slices treated with gadopentetate than in those treated with gadobutrol (6.07 ± 1.32 ion count vs 0.23 ± 0.02 ion count). Nevertheless, the mean ion count for 158Gd within the gadobutrol-treated slices was still on average 12 times higher compared with untreated control slices (0.23 ± 0.02 ion count vs 0.02 ± 0.01 ion count). Interestingly, in gadopentetate-treated slices, detected 191, 193Ir (labels nucleic acids) colocalized with areas of high 158Gd counts within the DG (Fig. 6C).
DISCUSSIONWe demonstrated previously that after repeated applications of gadopentetate, neuroinflammation favors the retention of Gd within the CNS in vivo.37 Here, in a murine model of multiple sclerosis as well as in living brain tissue, we compared the linear gadopentetate and the macrocyclic gadobutrol in terms of dynamics and patterns of inflammation-promoted Gd retention in vivo as well as potential cytotoxic effects ex vivo. Using T1 relaxometry and LA-ICP-MS, we found that gadopentetate was retained in the brain up to 40 days pfi, whereas gadobutrol was efficiently washed out toward day 40 pfi. Although for both GBCAs inflammation favored enhanced Gd retention within cerebellar structures, gadopentetate led to much larger Gd brain retention compared with gadobutrol. Finally, using an ex vivo model of neuronal tissue, we showed that gadopentetate but not gadobutrol induced cell death in the living brain tissue and that this process was enhanced in the presence of the inflammatory mediator TNF-α.
Our study revealed significantly decreased T1 relaxation times after multiple injections of GBCAs in EAE brains compared with HC. T1 relaxometry is a robust tool to study MRI alterations related to Gd retention after administration of GBCAs in a quantitative manner,42,43 without the need for a reference region, which in itself may be a confounding factor.13,44 Using the well-established ABA registration tool, we chose an automated approach to identify T1 relaxation decreases within different brain areas, ensuring higher validity and comparability. The LA-ICP-MS data confirmed the relaxometry findings, as cerebellar Gd levels in EAE mice were 2- to 3-fold higher than in HC mice and within the range previously reported by us and others.15,37,45 Maximal Gd contents inside the CN and DN after repeated injections of gadopentetate corresponded to approximately 4.51 μg/g in HC mice and 8.28 μg/g in EAE mice (30 μM and 55 μM, respectively).
Comparing the 2 applied GBCAs overtime, LA-ICP-MS data confirmed that neuroinflammation enhanced Gd retention especially after gadopentetate administration. We observed a long-term retention in the EAE cerebellum and its nuclei, and to a lower extent also in HC brains, up to 40 days pfi of the linear compound. However, gadobutrol administration led to a rather low and transient retention of Gd until day 10 pfi, which was also enhanced during inflammation but was completely cleared from the brain at day 40 pfi. Recent studies demonstrated a nonpermanent retention of macrocyclic GBCAs33,34,46,47 with clearance rates dependent on the specific washout kinetics.33,34 Frenzel et al suggested that gadobutrol remained in its intact form inside the body as it was exclusively present in the soluble fraction of the brain as a low-molecular-weight molecule. In contrast, linear GBCAs at day 24 pfi were present to a large extent in an insoluble form, whereas in its soluble fraction, a portion of Gd was bound to large macromolecules, suggesting that Gd had dissociated from the chelate.15 These findings were confirmed in recent rodent studies.45,48,49
We speculate that BBB disruption37 and/or alterations of the choroid plexus50 occurring during EAE contribute to the enhanced entry of GBCA into the brain, whereas inflammation-mediated tissue alterations51–54 may promote enhanced Gd retention. In healthy individuals, Gd retention inside the brain seems to be the result of an entry of GBCA through the blood-CSF barrier55 at the choroid plexus and from the CSF via the glymphatic system,56–59 into the brain. Along these lines, Öner et al60 reported on DN T1 hyperintensities after only a single intrathecal injection of gadopentetate in patients with normal renal function. It has been shown that alterations of the choroid plexus observed in multiple sclerosis61 and EAE62 may contribute to CNS pathology. Thus, an enhanced entry of GBCA into the CSF via the inflamed choroid plexus could be assumed.
Moreover, BBB breakdown is considered an early hallmark in multiple sclerosis that permits large CNS infiltration of peripheral activated autoreactive lymphocytes into the CNS. In addition, inflammation-related release of chemokines, free radicals, activated matrix metalloproteinases (MMP) and the consequent enhanced extravasation of immune cells may further compromise BBB integrity.63 Blood-brain barrier disruption occurs in a similar way in the EAE model, in which T lymphocytes have been demonstrated to induce tight junction alterations in BBB endothelium initiating barrier disruption and enhanced permeability.64,65 Whereas a leaky BBB may result in elevated GBCA diffusion, additional cellular and tissue alterations such as demyelination, microglial activation, changes in elemental metal distribution, or/and alterations of the extracellular matrix may contribute to enhanced retention. In this context, it has been suggested that inflammation-mediated alterations in the extracellular matrix composition such as glycosaminoglycans (GAGs) may favor GBCA dechelation and parenchymal retention of Gd.37 We previously reported that in EAE, inflammation-induced modifications of GAG sulfation on brain endothelial cells facilitate the uptake of very small superparamagnetic iron oxide nanoparticles.51 The GAG network on endothelial cells of vessels may act as an ion exchanger with complexing abilities.66 Interestingly, the DN was found to contain elevated amounts of microvessels that may bind Gd23 and to be enriched in endogenous metals such as Fe, Cu, and Zn that may facilitate transmetallation processes.67
Strikingly, we observed increasing Gd levels within the CN (DN included) from day 10 to day 40 pfi, which was observed also in HC but was more pronounced in the EAE animals. We speculate that this increase over time within the CN on day 10 and 40 pfi may be the result of a release of Gd3+ from other storage organs such as the bones.3,68–70 Stable Gd retention inside the bone has been reported for a period of more than 8 years most likely caused by the transmetallation of Gd3+ with resident calcium ions.69,71 Thakral and Abraham72 reported on increasing Gd amounts in sequential skin biopsies of patients with nephrogenic systemic fibrosis although no more GBCA was administered. Bone storage may also contribute to maintain Gd tissue levels by its mobilization into the bloodstream, since it was reported that Gd was still detectable in the red blood cell compartment 10 days pfi of principally linear GBCAs.18 Our ICP-MS analysis of blood samples revealed indeed that Gd was present in all samples up to day 40 pfi within the same range as detected in the cerebellum (approximately 0.2–2.0 μg/g). This supports the hypothesis of long-term retention, release, and recirculation of Gd through the body according to the Le Chatelier principle.73
We also showed that Zn levels in EAE brains were elevated compared with HC brains irrespective of the administered GBCA at all investigated time points. It has been suggested that disruption of brain Zn homeostasis may play an active role in the pathogenesis of multiple sclerosis and EAE.74,75 Zn acts as a cofactor for numerous enzymes like MMPs, is component of various proteins including myelin basic protein,74 and was reported to induce mitochondrial production of reactive oxygen species.75 Hence, increased neuronal Zn release may lead to MMP activation compromising BBB integrity and mediate neuronal injury. In this line, Choi et al75 reported on decreased BBB impairment and reduced immune cell infiltration and myelin destruction in the spinal cord of EAE mice after oral zinc chelator application.
We further observed that cerebellar Zn concentrations decreased until day 40 pfi, whereas Gd levels increased. Several studies reported on Zn depletion due to transmetallation with Gd and resulting zincuria after injections of linear GBCAs.31,76,77 Thus, increased Zn content in EAE brains may lead to elevated transmetallation rates with administered Gd and therefore facilitate its retention.
Furthermore, we investigated how GBCAs may affect cell viability using a model of organotypic hippocampal slice cultures in which the 3-dimensional histological architecture and
留言 (0)