
記住我










The human solute carrier 22 family (SLC22), also termed the organic ion transporter family, consists of 28 distinct multi-membrane spanning proteins, which phylogenetically cluster together according to their charge specificity for organic cations (OCTs), organic anions (OATs), and organic zwitterion/cations (OCTNs). Some SLC22 family members are well characterized in terms of their substrates, transport mechanisms, and expression patterns, as well as their roles in human physiology and pharmacology, whereas others remain orphans with no known ligands. Pharmacologically, SLC22 family members play major roles as determinants of the absorption and disposition of many prescription drugs, and several, including the renal transporters, OCT2, OAT1 and OAT3, are targets for many clinically important drug-drug interactions. In addition, mutations in some of these transporters [SLC22A5 (OCTN2) and SLC22A12 (URAT1)] lead to rare monogenic disorders. Genetic polymorphisms in SLC22 transporters have been associated with common human disease, drug response, and various phenotypic traits. Three members in this family were deorphaned very recently: SLC22A14, SLC22A15, and SLC22A24, and found to transport specific compounds, such as riboflavin (SLC22A14), anti-oxidant zwitterions (SLC22A15) and steroid conjugates (SLC22A24). Their physiologic and pharmacological roles need further investigation. This review aims to summarize the substrates, expression patterns and transporter mechanisms of individual SLC22 family members and their roles in human disease and drug disposition and response. Gaps in our understanding of SLC22 family members are described.
SIGNIFICANCE STATEMENT In recent years, three members of the SLC22 family have been deorphaned and found to play important roles in the transport of diverse solutes. New research has furthered our understanding of the mechanisms, pharmacological roles, and clinical impact of SLC22 transporters. This minireview provides an overview of the physiologic and pharmacologic roles and impact of genetic variants in the SLC22 family on disease and drug response, and summarizes recent studies deorphaning SLC22 family members.
Brief Historical PerspectiveThe SLC22 family belongs to the major facilitator superfamily (MFS). In the human genome, the SLC22 family includes 28 members: 23 SLC22A family members and five atypical SLC22B, which are designated synaptic vesicle proteins (SV2A-C) and synaptic vesicle 2-related proteins (SVOP and SVOPL) (Fig. 1) (Perland et al., 2017; Meixner et al., 2020). In 2004, the phylogenetic tree of the human SLC22 family only included 18 members (Koepsell and Endou, 2004). However, in 2007, the family was expanded to include vesicular proteins SV2 and SVOP, the unknown substrate transporters on chromosome 11q12.3, and others (Jacobsson et al., 2007; Fredriksson et al., 2008; Wu et al., 2009). Recently, a member, MFSD10, was annotated as SLC22A32, resulting in the current 28 members in the human SLC22 family (Meixner et al., 2020). All share a predicted 12-transmembrane domain structure with a large extracellular loop between transmembrane domain 1 and transmembrane domain 2 (Fredriksson et al., 2008). With the exception of MFSD10, which was crystallized in an outward-facing state at 2.6 Å resolution by X-ray crystallography (Pascoa et al., 2021), no other SLC22 family member has been crystallized or structurally resolved.
In 1994, the rat organic cation transporter (Oct1), Slc22a1, was the first member in the SLC22 family to be molecularly identified through expression cloning (Grundemann et al., 1994). Subsequently, in 1997, the human ortholog SLC22A1 (OCT1) and a paralog, SLC22A2 (OCT2), were identified (Gorboulev et al., 1997; Zhang et al., 1997). Soon after the identification of OCT1 and OCT2, two zwitterion transporters, OCTN1 (SLC22A4) and OCTN2 (SLC22A5), were cloned and characterized (Tamai et al., 1997; Wu et al., 1998). OCTN1 was at first thought to be a weak carnitine transporter and was later found to be an excellent transporter of ergothioneine (Grundemann et al., 2005). OCTN2 was immediately recognized as an important carnitine transporter in the intestine and kidney and was later discovered to be responsible for carnitine transporter deficiency (Nezu et al., 1999), a life-threatening autosomal recessive human disorder. Although it was reported that a novel kidney transporter in mice existed in 1997 (Lopez-Nieto et al., 1997), the human organic anion transporters were identified and functionally characterized in 1999 [SLC22A6 (OAT1) and SLC22A8 (OAT3)] (Race et al., 1999) and in 2001 [SLC22A7 (OAT2)] (Sun et al., 2001) (Fig. 2, Supplemental Table 1).
After publication of the sequence of the human genome in 2001, many more organic ion transporters in the SLC22 family were cloned and characterized. In 2003, the HapMap Consortium developed the first human genome-wide maps of common variation in three major ancestral populations(International HapMap, 2003; Claussnitzer et al., 2020). From 2000 to 2015, investigators associated with the Pharmacogenomics of Membrane Transporters project, funded by the National Institutes of Health (NIH) identified and functionally characterized genetic variants in membrane transporter genes, including many members of the SLC22 family (Fig. 2) (Shu et al., 2006; Kroetz et al., 2010). Variants were identified in DNA samples from four major ethnic groups (Fig. 2, Supplemental Table 1).
Direct species orthologs of most of the genes in the human SLC22A and SLC22B family exist; however, some of the genes have no direct species orthologs (Fig. 1 and 2). For example, the genes in the human chromosomal region 11q12.3 (hg38, chr11:63,079,940-63,410,294), which includes SLC22A9, SLC22A10, SLC22A24, and SLC22A25 have low sequence identity to the parallel mouse syntenic region localized on mouse chromosome 19. Notably, the genes in mouse chromosome 19 (chr19:7,643,247-8,392,204, mm39) are designated as Slc22a19, Slc22a26, Slc22a27, Slc22a28, Slc22a29, and Slc22a30 (Wu et al., 2009) and lack direct human orthologs. In addition, the orphan transporter SLC22A31 has no mouse ortholog. Conversely, the mouse transporters Slc22a21 (Octn3) and Slc22a20 (Oat6) (Wu et al., 2015a) have no human orthologs. In fact, the human SLC22A20 is annotated as a pseudogene by the National Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov/nuccore/NR_033396.1).
The major events that involved the discovery and characterization of members of the human SLC22 family are shown in Fig. 2, which begins with discovery of the molecular identities of members of the SLC22 family and includes functional characterization of their polymorphisms, knockout mice, and recent deorphaning studies.
Key Recent AdvancesIn this section, we summarize the function, tissue distribution and transport mechanisms of individual transporters in the SLC22 family. Further, we describe recent advances in deorphaning members of the SLC22 family, and present new information on genomics, pharmacogenomics and disease genetics of members of the SLC22 family.
Organic Cation Transporters: Function; Tissue Distribution and Transport MechanismThree transporters with known preference for organic cations are members of the SLC22 family: OCT1 (SLC22A1), OCT2 (SLC22A2), and OCT3 (SLC22A3). In the human genome, the genes encoding these transporters are located on chromosome 6 and are about 70% identical to one another. Localized primarily to the plasma membrane, OCT1, OCT2 and OCT3 are facilitated transporters that are independent of sodium or proton gradients. These transporters have been extensively studied and excellent review articles have been recently published (Lai et al., 2018; Koepsell, 2020; Samodelov et al., 2020; Koepsell, 2021b; Koepsell, 2021a). Their function, tissue distribution and substrate selectivity are briefly summarized below.
OCT1 (SLC22A1)As the first member of the human SLC22 family to be cloned and functionally characterized (Fig. 2), OCT1 is among the most highly expressed SLC transporters in the human liver (Fig. 3, Supplemental Table 2). Studies have shown significant correlation of OCT1 transcript and protein levels in liver tissues or hepatocytes (Nies et al., 2009; Fattah et al., 2017) and OCT1 protein expression is lower in neonates and significantly higher in adults (Prasad et al., 2016). In contrast, OCT1 has extremely low expression levels in all other tissues except whole blood, where its expression levels are detectable and higher than its two paralogs, OCT2 and OCT3. Proteomic studies showed protein expression levels of OCT1 in the small intestine although much lower levels than other known transporters, such as OATP2B1 (Drozdzik et al., 2019; Kiss et al., 2021). OCT1 promiscuously transports structurally diverse organic cations, including naturally occurring compounds, such as thiamine (vitamin B1) and histamine, and many synthetic drugs (e.g., metformin and sumatriptan) (Table 1). The transport mechanism is bidirectional and is driven by the substrate concentration and potential difference across the plasma membrane (Fig. 4). Inhibitors of the transporter include a diverse array of prescription drugs (Ahlin et al., 2011; Chen et al., 2017; Meyer et al., 2019; Koepsell, 2021b). Canonical substrates of OCT1 used in functional assays in cells are TEA (tetraethylammonium) and MPP+ (N-methylpyridinium). ASP+ (4-(4-(dimethylamino)styryl)-N-methylpyridinium iodide) has been used as a fluorescent substrate of the transporter and its paralogs (Gorboulev et al., 1997; Zhang et al., 1997; Ciarimboli et al., 2004). Canonical inhibitors of OCT1, which can be used in cellular assays, include verapamil and quinidine.
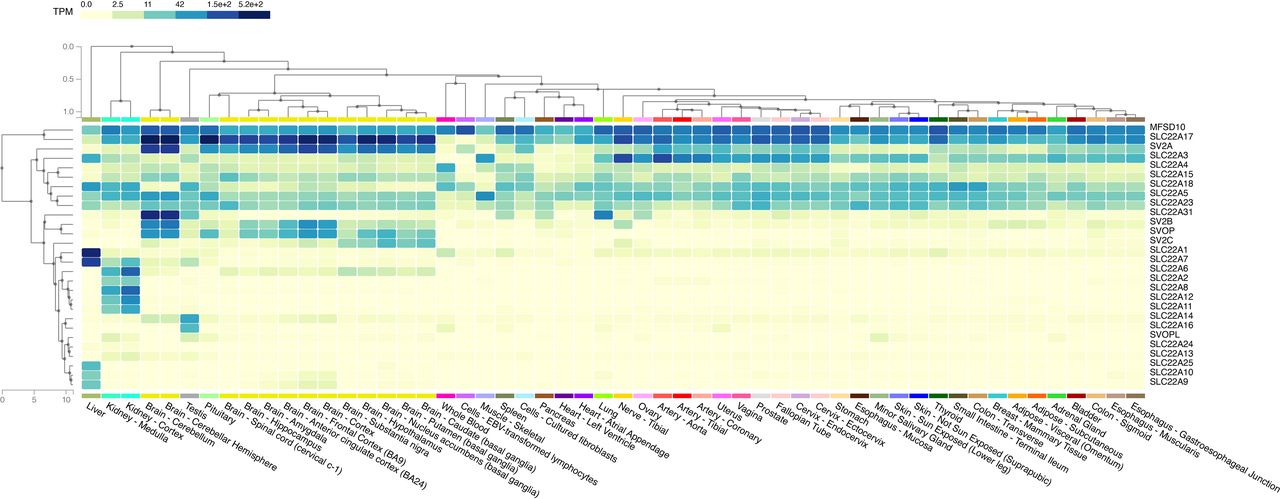 Fig. 3.
Fig. 3. Transcriptomic measurements of 28 members in the solute carrier 22 family across 54 tissues. Transporters which have similar transcriptomic profiles are clustered closer together. For example, major facilitator superfamily D10 and solute carrier 22A17 are expressed at high abundance (darker blue) across almost all tissues and thus are next to each other. See Supplemental Table 2 to obtain the median transcripts per kilobase million values for each transporter and tissue. The data used in this figure were obtained from the Genotype-Tissue Expression (GTEx) Multi Gene Query page (https://www.gtexportal.org/home/multiGeneQueryPage) (GTEx Consortium, 2020). This visualization page allows a user to enter a list of genes and to display the expression in a heat map across tissues. The heat map is generated using the hierarchical clustering method described in https://gtexportal.org/home/home/news?id=360 (section 2018-04-04). The GTEx Project was supported by the Common Fund of the Office of the Director of the National Institutes of Health, and by National Cancer Institute, National Human Genome Research Institute, National Heart, Lung, and Blood Institute, National Institute on Drug Abuse, National Institute of Mental Health, and National Institute of Neurological Disorders and Stroke.
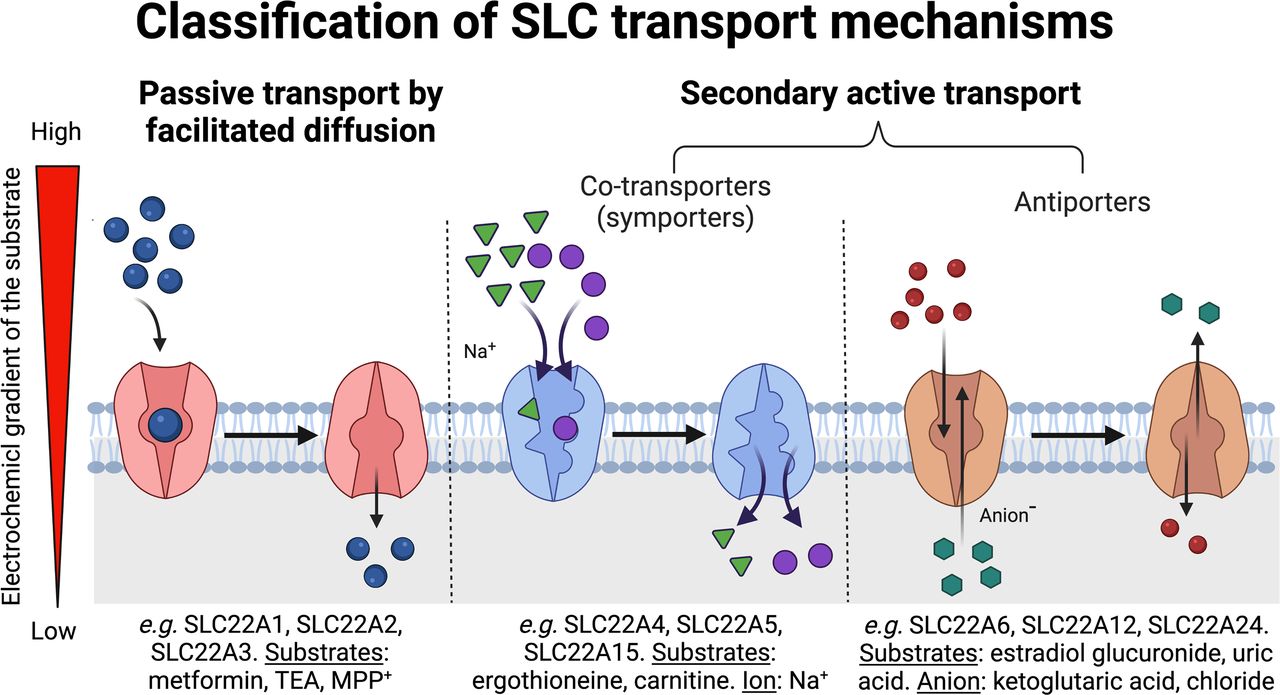 Fig. 4.
Fig. 4. Classification of solute carrier transport mechanisms. Circles depict substrate. The arrows show the general direction of flux. Triangles represent sodium and hexagons represent anions, which may include organic anions, such as glutaric acid. Sodium and anions are ions that provide a driving force for transport by moving down their concentration gradients. Examples of transporters in the solute carrier 22 family that function to move substrates across the plasma membrane by one of the mechanisms (passive transport or secondary active transport) from high to low concentrations are shown in the figure. Secondary active solute carrier transport can be classified further into co-transport (symport) and antiport. Figure was created with Biorender.com.
TABLE 1Substrates, membrane localization, genetic polymorphisms, and associated phenotypes of transporters in the solute carrier 22 family
Data not available or not known is noted as unknown.
OCT2 (SLC22A2)In contrast to OCT1, with its primary expression in the liver, OCT2 is highly expressed in the kidney, where it is localized to the basolateral membrane of the renal tubule. OCT2 expression in the kidney is largely in the proximal tubule (Fig. 3), but it is also present in the l of Henle and distal tubule (Humphreys, 2021). Targeted proteomic studies showed high abundance of OCT2 in human kidney and a strong correlation between transcript and protein levels of OCT2 in the kidney (Cheung et al., 2019). Similarly, to OCT1, OCT2 protein levels are lower in neonates (Cheung et al., 2019) but similar across children, adolescents, and adults (Li et al., 2019). It works in concert with apical membrane transporters, e.g., multi-drug and toxin extrusion proteins (MATEs, SLC47), to mediate secretory flux from blood to urine of many basic drugs. The substrate selectivity of OCT2 overlaps extensively with OCT1. That is, the transporter interacts with structurally diverse basic drugs and naturally occurring amines. Canonical substrates of OCT2 include the anti-diabetic drug metformin and the anti-cancer drug cisplatin. OCT2 is thought to play major roles in the renal elimination of basic drugs and the renal toxicity of cisplatin. Canonical inhibitors include cimetidine, trimethoprim, and pyrimethamine (Koepsell, 2020); however, these compounds also inhibit MATEs, albeit with different kinetic properties. Thus, in a clinical situation or in vivo in animals, it is often difficult to pinpoint the transporter implicated in a renal drug-drug interaction.
OCT3 (SLC22A3)In contrast to both OCT1 and OCT2, which are highly expressed in a single tissue, OCT3 is expressed in many tissues, including arterial tissues, tibial nerve, prostate, ovaries, and uterus (Fig. 3). It is also expressed in tissues important in drug absorption and elimination, including the liver, kidney, and intestine, in both transcript and proteomic studies (Nies et al., 2009; Drozdzik et al., 2019; Oswald et al., 2019). In the intestine, OCT3 appears to be localized to the apical membrane and to play a role in drug absorption (Chen et al., 2015), whereas in the liver and kidney, the transporter appears to co-localize with its paralogs, OCT1 and OCT2 on the basolateral membrane. In organs of elimination, OCT3 is expressed at substantially lower levels than its paralogs, suggesting that it plays a lesser role in the elimination of most drugs. Like OCT1 and OCT2, the transport mechanism for OCT3 is facilitated and based on the electrochemical gradient across the plasma membrane. The substrate selectivity of OCT3 overlaps substantially with OCT1 and OCT2. Endogenously, the transporter appears to be involved in the reuptake of catecholamines, thus regulating local concentrations of neurotransmitters in the vicinity of their receptors (Song et al., 2019). In cancer, inhibition of OCT3 transport has been shown to be protective against doxorubicin-induced cardiac injury (Huang et al., 2021). In addition, loss of OCT3 appears to play a role in promoting hepatic fibrosis, where Oct3−/− mice showed increasing expression levels of transforming growth factor β1 (both protein and mRNA), as well as upregulation of transforming growth factor β-dependent genes (Vollmar et al., 2019).
Organic Anion Transporters: Function, Tissue Distribution, and Transport MechanismThe human organic anion transporter subfamily of the SLC22 family consists of eight transporters: OAT1 (SLC22A6), OAT2 (SLC22A7), OAT3 (SLC22A8), OAT4 (SLC2A11), OAT7 (SLC22A9), OAT10 (SLC22A13), SLC22A24, and URAT1 (SLC22A12). SLC22A24, which was recently deorphaned, will be described in the section Recent Deorphaning Studies of Transporters in the SLC22A Family. Localized primarily to the plasma membrane, these transporters have distinct mechanisms to mediate transmembrane flux of their substrates. Below, their function, specificity, tissue distribution and other molecular characteristics are described. Excellent reviews, which describe most of these transporters, have been recently published (Nigam, 2015; Nigam, 2018; Kuang et al., 2021; Zhang et al., 2021).
OAT1 (SLC22A6)The first member of the organic anion transporter subfamily, OAT1 is highly and predominantly expressed in the human kidney (Fig. 3). To handle the high plasma levels of organic anions, the transporter is exclusively localized to the basolateral membrane of the proximal tubule (Breljak et al., 2016; Humphreys, 2021). Proteomic studies showed significant correlation with transcript levels and high abundance in the kidney (Nakamura et al., 2016; Cheung et al., 2019; Li et al., 2019; Oswald et al., 2019). The mechanism by which OAT1 transports its solutes across the plasma membrane is through exchange with the dicarboxylic acid, alpha-ketoglutarate, which is transported into the renal epithelia by sodium-dependent transporters in the SLC13 family. Thus, the transport mechanism is considered to be tertiary active. That is, the primary active transporter, sodium-potassium ATPase, creates an inwardly-directed sodium gradient, which in turn, drives the intracellular accumulation of alpha-ketoglutarate. Alpha-ketoglutarate then flows down its concentration gradient in the efflux direction via OAT1 in exchange for an OAT1 substrate (in the influx direction) (Lu et al., 1999; Uwai et al., 2017). Typical substrates of OAT1 include drugs (e.g., penicillins, cephalosporins, non-steroidal inflammatory drugs, and anti-viral drugs), toxins (ochratoxin), and a range of compounds derived from gut bacterial or human metabolism, such as uric acid, para-amino hippuric acid, and indoxyl sulfate (Table 1). Like OCT2, the transporter is a target for renal drug-drug interactions, and therefore is studied extensively during drug development (Giacomini et al., 2010; Zamek-Gliszczynski et al., 2018).
OAT3 (SLC22A8)Similar to OAT1, OAT3 is highly and predominantly expressed in the human kidney, with much lower levels of expression in other tissues (Fig. 3). Proteomic studies showed similar abundance of OAT1 and OAT3 in the kidney, with lower expression levels in the neonates (Cheung et al., 2019; Li et al., 2019; Oswald et al., 2019). Its high levels of expression in the kidney is consistent with the transporter’s role in eliminating many naturally occurring and synthetic compounds (Nigam, 2015; Nigam, 2018; Kuang et al., 2021; Zhang et al., 2021). Single-cell RNA-seq data place OAT3 in all three segments of the proximal tubule (S1, S2 and S3) (Humphreys, 2021), and, along with OAT1, OAT3 has been localized to the basolateral membrane serving in the secretion of its substrates (Breljak et al., 2016). In the choroid plexus, the transporter appears to move its substrates from the CSF back to the blood (Uchida et al., 2020). Like OAT1, OAT3 is a tertiary active transporter moving its substrates into the proximal tubule cells in exchange for alpha-ketoglutarate (Burckhardt et al., 2005; Burckhardt, 2012; Uwai et al., 2017) (Fig. 4). Pharmacologically, OAT3 interacts with structurally diverse organic anions and has overlapping but broader substrate specificity compared with OAT1 (Table 1). Recent studies suggest that both OAT1 and OAT3 potently interact with drug metabolites, including sulfate and glucuronide conjugates as well as oxidized metabolites (Wu et al., 2017; Zou et al., 2021); however, OAT3 has a broader selectivity for metabolites than OAT1 (Astorga et al., 2011). Consistently, OAT3, in contrast to OAT1, can interact potently with certain non-anionic compounds, including some basic drugs, such as cimetidine, where it may play a role in their renal elimination (Tahara et al., 2005).
OAT2 (SLC22A7)In contrast to OAT1 and OAT3, which have a predominant localization to the kidney, OAT2 is expressed abundantly in both the kidney and liver, with lower levels of expression in other tissues and organs (Fig. 3). This is similar to proteomic studies of the liver and kidney (Nakamura et al., 2016; Oswald et al., 2019). The transporter interacts with many organic anions such as para-aminohippuric acid, and various anti-viral drugs, as well as other compounds. For example, OAT2 is also capable of transporting various nucleobases, nucleosides, and nucleotides, including the cyclic nucleotide cGMP as well as nucleoside analog drugs (e.g., ganciclovir, warfarin) (Bi et al., 2018; Mathialagan et al., 2018). Recent studies suggest that the transporter works with other hepatic uptake transporters to mediate drug disposition (Zamek-Gliszczynski et al., 2018). Its role can be distinguished from other liver organic anion transporters with the use of a specific inhibitor, ketoprofen (Bi et al., 2019), and penciclovir has been proposed as a specific substrate for use in human hepatocytes (Mathialagan et al., 2018).
OAT4 (SLC22A11)Most highly homologous to URAT1 (SLC22A12) in the SLC22 family (Fig. 1), OAT4 is highly and selectively expressed in the kidney; however, its transcript and proteomic levels are more akin to OCT2 than to the much higher levels of OAT1 or OAT3 (Fig. 3) (Li et al., 2019) . Unlike OAT1, OAT2 and OAT3, OAT4 has an apical localization in the proximal tubule where it plays a role in renal reabsorption (Miyazaki et al., 2005). It appears to be localized to segments 1 and 3 of proximal tubule epithelial cells and at lower levels in other cell types in the kidney (Humphreys, 2021). The transporter is also found in human placenta, where it appears to be responsible for the removal of sulfated conjugates of steroids from the fetus (Ugele et al., 2003). OAT4 also exhibits a diverse substrate specificity with a preference for organic anions. Typical substrates include uric acid, the toxin ochratoxin A, and estrone sulfate (Table 1). Studies showed that substitution of chloride in the uptake buffer enhanced uptake of estrone sulfate in cells expressing OAT4 (Hagos et al., 2007). Recent studies showed that OAT4 efflux glutamate and aspartate, similar to OAT10 (Skwara et al., 2017). There is no mouse ortholog for this gene.
OAT7 (SLC22A9)OAT7 has not been well-studied. It is expressed in the liver at approximately 6% of the transcript levels of OAT2 according to GTEx portal, with lower expression levels in the brain and negligible expression levels in other tissues (Fig. 3). Protein levels of OAT7 in the human liver showed similar abundance as OAT2 (Vildhede et al., 2018). Specific substrates for the transporter have been difficult to identify. Recent studies have demonstrated that estrone 3-O-sulfate and dehydroepiandrosterone 3-O-sulfate are excellent substrates but not specific, as OATPs are known to take up sulfated steroids (Mathialagan et al., 2018). Orthologs of OAT7 are found in primates but not rodents.
OAT10 (SLC22A13)With its highest expression levels in the kidney according to GTEx (Fig. 3), OAT10 has been the subject of recent studies of uric acid disposition (Wang et al., 2020a). The transporter has been recognized as a uric acid reabsorptive transporter localized to the apical membrane of the renal tubule (Burckhardt, 2012; Otani et al., 2020). Single-cell RNA-seq data, however, suggest that in comparison with URAT1 (SLC22A12), OAT10 is expressed at much lower levels in the proximal tubule (Humphreys, 2021). The transporter has a high affinity for nicotinic acid and a lower affinity for uric acid (Bahn et al., 2008). It also takes up the typical OAT substrate, para-amino hippuric acid, with optimal uptake at pH 5, suggesting that it may act as a proton-organic anion symporter.
URAT1 (SLC22A12)With higher transcript and protein expression levels in the kidney and much lower levels in other tissues, URAT1 is a selective uric acid transporter (Fig. 3) (Nakamura et al., 2016; Cheung et al., 2019). Expressed on the apical membrane of the renal proximal tubule, URAT1, together with other transporters, serves in the reabsorption of uric acid. The transporter acts as an anion exchanger taking up uric acid in exchange for inorganic (e.g., Cl-) and organic anions (e.g., lactate, nicotinate). It is the target of many uricosuric drugs including lesinurad and newer drugs (Bardin and Richette, 2018). URAT1 has been extensively discussed in many review articles on gout and uric acid disposition (see Koepsell, 2013; Nigam and Bhatnagar, 2018; Dong et al., 2019) and therefore will not be further reviewed here.
Zwitterion Transporters: Function, Tissue Distribution, and Transport MechanismFour family members, which primarily transport zwitterions cluster together phylogenetically in the human SLC22 family. These include OCTN1 (SLC22A4), OCTN2 (SLC22A5), SLC22A15, and SLC22A16 (fly-like putative transporter 2). SLC22A15 was recently deorphaned (see section Recent Deorphaning Studies of Transporters in the SLC22A Family). As with other SLC22 family members, the transporters all contain 12 transmembrane domains and a large extracellular loop between the first and second transmembrane domain. A few review articles about OCTNs have been recently published (Pochini et al., 2019; Betterton et al., 2021; Sweet, 2021). Below, we briefly describe OCTN1, OCTN2, and SLC22A16.
OCTN1 (SLC22A4)OCTN1 is expressed in a variety of tissues throughout the body and in particular, in whole blood (Fig. 3). Protein expression levels of OCTN1 in the kidney are lower than OCT2 and OATs (Li et al., 2019) and its protein is also expressed in the small intestine (Nakamura et al., 2016). OCTN1 is sometimes termed the ergothioneine transporter (Grundemann et al., 2005). Ergothioneine is a naturally occurring zwitterion, which is derived from fungi and some bacterial species. Its levels in whole blood are dependent on OCTN1 expression, consistent with a major role of OCTN1 in ergothioneine disposition. Ergothioneine is thought to function as an important anti-oxidant in the body. Though not a true vitamin with a deficiency syndrome, ergothioneine has been proposed to be a “longevity” vitamin (Ames, 2018) because of its potent anti-oxidant activity and high levels in the human body. OCTN1 transports ergothioneine in a sodium-dependent fashion, thus promoting the intracellular accumulation of ergothioneine in tissues in which it is expressed. In addition, OCTN1 transports various organic cations, including choline, acetylcholine and tetraethylammonium (Pochini et al., 2019). There are some data suggesting that the transporter may function biologically in the release of acetylcholine. OCTN1 appears to have a complex transport mechanism that depends on both the direction of transport (influx or efflux) as well as the charge of its substrates. In particular, though influx of ergothioneine is sodium-dependent, influx of acetylcholine, a cation, is inhibited by sodium. In contrast, acetylcholine efflux is not sodium-sensitive. Proteoliposomes have been used to characterize OCTN1 function, which provides excellent support for its transport mechanism as well as its substrate selectivity (Pochini et al., 2019). Drugs that have been identified as substrates of OCTN1 include metformin, sulpiride, gabapentin, and cytarabine, although these substrates are controversial (Tschirka et al., 2018) (Table 1).
OCTN2 (SLC22A5)In contrast to OCTN1, OCTN2 is primarily a carnitine transporter. The transporter gene is located together with OCTN1 in a locus on chromosome 5. With ubiquitous expression in most tissues in the body, including the proximal tubule and intestine (Kato et al., 2006), OCTN2 serves in carnitine homeostasis providing 70 to 80% of the body’s supply of carnitine through intestinal dietary absorption and renal reabsorption mechanisms. In humans, transport of carnitine across the intestinal basolateral membrane transporter is not well-established. Studies suggest that the rodent ortholog of OCTN3, which is localized to the basolateral membrane in intestinal epithelia, may play a role in carnitine absorption (Duran et al., 2005; Garcia-Miranda et al., 2005). The remainder of the body’s carnitine is synthesized. Essential in fatty acid oxidation, carnitine is needed by virtually all tissues. Carnitine transporter deficiency (CTD) is an autosomal recessive disease caused by mutations in OCTN2 (see section Rare Diseases Involving Mutations in SLC22 Family). OCTN2 is a sodium co-transporter and serves in carnitine influx. It also interacts with carnitine acyl esters as well as various synthetic molecules. The transporter can serve in a sodium-independent fashion in efflux of its substrates and in particular, acyl-carnitine esters when their intracellular concentrations exceed their extracellular concentrations. Drugs that are substrates of OCTN2 include mildronate and sulpiride (Grigat et al., 2009; Li et al., 2017) (Table 1). OCTN2 and CTD have been the subject of many review articles (Longo, 2016; Almannai et al., 2019; Pochini et al., 2019).
Fly-like Putative Transporter 2 (SLC22A16)Sometimes termed CT2 or fly-like putative transporter, SLC22A16 is a high-affinity carnitine transporter (Enomoto et al., 2002b). Only a few studies characterizing the function of SLC22A16 have been published (Table 1). The transporter is highly expressed in human testes, with much lower levels of expression in other tissues. It appears to be localized to Sertoli cells within the testis and to the luminal membrane of epididymal epithelium. Carnitine is essential in the regulation of motility of spermatozoa as well as to their maturation. Because of its polarity, carnitine clearly needs a transporter to foster its intracellular accumulation. In the testes, SLC22A16 is a highly specialized transporter. SLC22A16-mediated carnitine uptake is inhibited by acyl-carnitine derivatives as well as betaine. SLC22A16 is over-expressed in some cancers, such as acute myeloid leukemia (Wu et al., 2015b), and its over-expression is tied to the growth and viability of the tumor cells, suggesting that it may be a viable target for the treatment of cancers.
SLC22B Synaptic Vesicular Glycoprotein 2The SLC22B subfamily is comprised of three synaptic vesicle glycoprotein 2 members [SV2A (SLC22B1), SV2B (SLC22B2) and SV2C (SLC22B3)] and SVOPL and SVOP. These proteins are most closely aligned with SLC22A18 and MFSD10 in the SLC22 family. Multiple sequence alignments of the five SLC22B family members with MFSD10 and SLC22A18 result in approximately ∼13 to 19% identity, suggesting that SLC22A and SLC22B are related but do not meet the 25% criteria for sequence homology of family members. The SV2 family has recently been reviewed (Janz et al., 1998; Bartholome et al., 2017; Stout et al., 2019) and is briefly described here. Consistent with other SLC22 family members, SV2 family members are 12-transmembrane proteins, which have about 65% sequence homology with one another. These proteins are expressed in secretory vesicles, such as synaptic vesicles, and seem to be important in neurotransmission, yet their exact function remains unknown (Bajjalieh et al., 1993; Crowder et al., 1999). Nevertheless, the proteins, and in particular SV2A, have been targeted by small molecules. For example, levetiracetam targets SV2A to treat epilepsy. The drug was approved for clinical use before its target was identified. Within the brain, the three proteins have distinct expression patterns, with SV2A being the most ubiquitous followed by SV2B which is found primarily in the trigeminal and motor nuclei followed by SV2C, which is found in the striatum, midbrain, and ventral pallidum. Despite extensive experimentation, the precise function of these proteins is unknown. The proteins have been thought to function in vesicular transport similar to the monoamine transporters in the SLC18 family; however, to date, evidence for vesicular transport function has not been found. SV2A has been shown to transport galactose when vesicles are fused with the plasma membrane (Madeo et al., 2014). SV2 proteins in vesicles that are fused to the plasma membrane may interact with botulinum and tetanus neurotoxins (Dong et al., 2006; Stout et al., 2019). The three proteins are implicated in Alzheimer’s disease and Parkinson’s disease67, although until their precise functions are identified, the mechanisms for their roles in neurodegeneration will remain elusive. SVOP (SLC22B4) and SVOPL (SLC22B5) are more distantly related to SV2, but nevertheless in the SLC22B family. The proteins are both vesicular. SVOP has been studied in knockout mice and is clearly not essential for the fertility or viability of the mice, which have no obvious phenotypes (Yao et al., 2013). Like SV2A, SVOP is ubiquitously expressed in all brain regions. SVOPL is a paralog of SVOP (Jacobsson et al., 2007). Little is known about the gene and protein; however, the gene appears to be maternally imprinted with the paternal allele inactivated through DNA methylation (Yao et al., 2013). Expressed earlier in development, SVOPL expression levels in the brain decline with aging.
Recent Deorphaning Studies of Transporters in the SLC22A FamilyAmong the 28 members of the SLC22 family, ten are orphans. Six of the orphans are SLC22A members and four are atypical SLC22B members. These ten orphan transporters have no known substrates. Three of the 28 SLC22 members were deorphaned in the last few years (Fig. 2). These are SLC22A24 (Yee et al., 2019), SLC22A15 (Yee et al., 2020) and SLC22A14 (Kuang et al., 2021). Notably, untargeted metabolomic approaches were used as the primary method to identify substrates of these recently deorphaned transporters. Other methods were used in the deorphaning studies, for example, microscopy methods to localize the transporter as well as structural method to detect the charge specificity of the transporters.
SLC22A24In the phylogeny tree, SLC22A24 clusters with the organic anion transporters, SLC22A6, SLC22A8, SLC22A11, and SLC22A12. SLC22A24 substrates were discovered using a combination of genome-wide association metabolomic studies (GWAS-metabolomic) and untargeted metabolomic methods in HEK293 cells overexpressing SLC22A24 (Yee et al., 2019). In the GWAS-metabolomic studies, a common nonsense mutation in SLC22A24, p.Tyr501Ter was found to be associated at genome-wide levels of significance with lower plasma levels of three steroid metabolites, progesterone, etiocholanolone glucuronide, and androsterone glucuronide. In the untargeted metabolomic studies, cells recombinantly expressing SLC22A24 were exposed to media containing fetal bovine serum and found to take up several steroid sulfate and glucuronide conjugates, bile acids and dicarboxylic acids. The transport mechanism was determined to be independent of sodium and involve exchange with glutaric acid. Because the variant, p.Tyr501Ter is common, transcript levels of SLC22A24 are difficult to detect presumably because of nonsense mediated decay. However, according to data from single cell RNAseq (Humphreys, 2021), SLC22A24 is expressed in segment 3 of the renal proximal tubule. Further, because the p.Tyr501Ter, which was found to be non-functional, was associated with reduced plasma levels of steroid conjugates, the transporter was presumed to function in the reabsorption of its substrates and play a role in steroid homeostatic mechanisms. Sequencing studies have revealed that approximately 50–80% of individuals in all populations have at least one allele of SLC22A24 containing the nonsense polymorphism (rs11231341), suggesting the transporter is disappearing from the human genome. In fact, SLC22A24 was also found to have interesting evolutionary pattern, with great apes having a direct ortholog of SLC22A24 but old and new world monkeys lacking a direct ortholog. Further, the gene was not found in many mammals, but was found in mice, lemurs, horses, and rats. However, species differences in the substrate specificity of the SLC22A24 orthologs were notable, suggesting that the transporter is under evolution pressure. Future studies are needed to determine the membrane localization of SLC22A24 and its pharmacological role in drug disposition.
SLC22A15In the phylogenetic tree, SLC22A15 clusters together with zwitterion transporters, SLC22A4, SLC22A5 and SLC22A16. To deorphan this transporter, untargeted metabolomic approaches were used in cells recombinantly expressing SLC22A15. These approaches showed that SLC22A15 transports zwitterions such as ergothioneine, carnitine, betaine, as well as to a lesser extent cations including MPP+ and neurotransmitters. Prior to characterization of SLC22A15 (Yee et al., 2020), only SLC22A4 was a known specific ergothioneine transporter (Grundemann et al., 2005); however, questions remained about how ergothioneine, a hydrophilic zwitterion found in abundance in the brain, entered brain parenchyma. The deorphaning studies show that SLC22A15 transports ergothioneine, but with a higher Km (affinity) than SLC22A4. Like SLC22A4, SLC22A15 transports its substrates in a sodium dependent fashion, and is thus able to concentrate its substrates intracellularly. Though both transporters are expressed in multiple tissues, SLC22A15 is expressed more abundantly in various regions of the brain (Fig. 3) suggesting it may play a role in the CNS disposition of ergothioneine and carnosine, two important anti-oxidants. SLC22A15 has been shown to play a role in cell proliferation (Zhu et al., 2019; Fang et al., 2021).
SLC22A14SLC22A14 is a specific transporter in the testes. Male Slc22a14 knockout mice and also male mice with a deletion on chromosome 9, which includes Slc22a14 along with several other genes, are phenotypically infertile (Runkel et al., 2008; Maruyama et al., 2016). However, the mechanism for the infertility remained unclear until a recently published deorphaning study revealed the substrate of SLC22A14 (Kuang et al., 2021). Unlike SLC22A15 and SLC22A24, which are expressed on the plasma membrane, SLC22A14 and its mouse ortholog Slc22a14 are expressed in the mitochondria. Therefore, mitochondrial lysates from HEK293 cells overexpressing empty vector, and human and mouse SLC22A14 were isolated and subject to untargeted metabolomic analyses (Kuang et al., 2021). Riboflavin was the top metabolite and was present at significantly higher levels in the mitochondrial lysate from HEK293 cells overexpressing human or mouse SLC22A14 compared with empty vector. Other metabolites relevant to the role of riboflavin in energy metabolism were also modulated. These include fatty acids and triacylglycerides. In further elegant studies, the mechanisms by which riboflavin deficiency in Slc22a14 knockout mice caused infertility were revealed and included reduced sperm motility leading to infertility. It is not known whether SLC22A14 transports other anions that may overlap with SLC22A13 since they cluster together phylogenetically (Fig. 1).
Pharmacogenetics of SLC22 TransportersGenetic polymorphisms in the coding and non-coding regions of ten members in the SLC22 family have been discovered and functionally characterized (Fig. 2, Supplemental Table 1). At about the same time as the publication of the draft human genome sequence, common genetic variants in SLC22A1 and SLC22A2 were discovered through sequencing approximately 200 DNA samples from ethnically diverse populations. These variants were functionally characterized to determine their effects on transporter activity or expression levels (Kerb et al., 2002; Leabman et al., 2002; Leabman et al., 2003). Several of these coding variants have been significantly associated with drug disposition in candidate gene studies (Supplemental Table 1) (Yee et al., 2018). The first candidate gene study of an SLC22 family member was performed by the Giacomini Laboratory to determine the effect of OCT1 reduced function variants (p.R61C, p.G401S, p.M420Del, p.G465R) on metformin disposition and response (Shu et al., 2007; Shu et al., 2008). Since then, many other studies have documented the impact of reduced function variants in OCT1 on the disposition, toxicity, and response of a wide range of basic drugs. These include metformin (Song et al., 2008), morphine (Tzvetkov et al., 2013; Balyan et al., 2017), ondansetron (Tzvetkov et al., 2012), sumatriptan (Matthaei et al., 2016), and, more recently, amitriptyline (Matthaei et al., 2021). Importantly, these reduced function variants are common in individuals of European ancestry, which range from 2% (OCT1 p.G401S) to 18% (OCT1 p.M420Del). Though absent in East Asians, OCT1 p.M420Del is present other ethnic groups, albeit at variable allele frequencies (African ancestry 5%, Latino 29%, South Asians 15%, Central Asia 20%, Middle East 10%) (Seitz et al., 2015). The common missense variant in East Asian is OCT1 p.P341, although predicted to be deleterious, but did not show any functional differences among the substrates screened (Shu et al., 2007; Seitz et al., 2015). This growing body of literature led the International Transporter Consortium to designate polymorphisms in OCT1, along with OATP1B1 (SLCO1B1) and breast cancer resistance protein (ABCG2), as important determinants of drug disposition and response (Yee et al., 2018).
Unlike OCT1 which has many common missense polymorphisms, other members of the SLC22 family have fewer common missense variants and only a few of these have shown significant associations with drug disposition or response. Although the missense variant OCT2 p.A270S (rs316019) has been widely studied, the effect of this variant is inconsistent across different ethnic groups with respect to metformin disposition (Wang et al., 2008; Chen et al., 2009; Tzvetkov et al., 2009; Zolk et al., 2009; Yoon et al., 2013; Moon et al., 2018; Kuhlmann et al., 2021) and is weakly or not associated with the disposition of other drugs (Borghetti et al., 2019; Yamamoto et al., 2019; Costa et al., 2021). The allele frequency of OCT2 p.A270S ranges from 5% in Latino to 15% in African American (https://gnomad.broadinstitute.org/).
However, SLC22A2 polymorphisms, including rs316019, have shown significant associations with serum creatinine and estimated glomerular filtration rate in genomewide association studies, perhaps due to larger sample sizes, usually of at least 5000 individuals (Shin et al., 2014; Pattaro et al., 2016; Gorski et al., 2017; Kanai et al., 2018). The impact of genetic variations in organic cation transporters on drug disposition has been recently reviewed (Koepsell, 2020; Zazuli et al., 2020; Kolz et al., 2021).
留言 (0)