
記住我



The severe acute respiratory syndrome coronavirus 2 (SARS-Cov-2) pandemic represents an extraordinary challenge for global health. It has affected hundreds of millions of people worldwide and had already led to more than five million deaths as of December 2021. SARS-CoV-2 seroprevalence in people with HIV (PWH) varies considerably between countries, from 0.72 to 8.5% [1–4]. Conflicting findings have been published concerning the effect of HIV infection on the risk of severe coronavirus disease 2019 (COVID-19) but three recent meta-analyses all found that HIV infection was associated with a higher risk of death from COVID-19 [5–7].
Vaccination against COVID-19 is an essential weapon in the fight to limit the consequences of the SARS-CoV-2 pandemic. Its effectiveness for reducing a wide range of COVID-19-related outcomes, including mortality, which has been proven in the healthy population in the real-world setting, would, of course, be expected for the most vulnerable individuals, such as immunocompromised PWH [8].
One of the vaccines authorized in France is an mRNA encoding the spike protein of SARS-CoV-2 formulated in lipid nanoparticles, the BNT162b2 vaccine (Pfizer-BioNTech, Pfizer-France, Paris, France). Phase II/III studies of this vaccine have shown it to be highly effective and well tolerated [9,10]. Here, we studied PWH, for whom data concerning the effects of this vaccine are lacking.
Another key issue is the worldwide emergence of SARS-CoV-2 variants, some of which partially escape neutralizing antibodies [11], because of amino-acid deletions and substitutions in functional regions of the receptor-binding domain (RBD). NAb titers after mRNA vaccination were one-third lower for the alpha (B.1.1.7) variant of concern (VOC) [12] than for the historical B.1 virus [13–15]. The beta VOC (B.1.351) had a greater capacity to escape neutralization, with NAb titers for this variant 14-fold lower than those for the historical B.1 virus in both vaccinated and convalescent individuals [13–15]. NAb titers for delta VOC (B.1.617.2) are 3-fold to 5-fold lower than those for the alpha variant in vaccinated individuals [16]. There is a need to obtain more data concerning the efficacy of the BNT162b2 vaccine in populations of frail individuals, such as PWH.
We conducted a multicenter cohort study of people at risk of developing severe forms of COVID-19, including PWH, with the primary objective of evaluating the neutralizing capacity of antibodies against different variants of SARS-CoV-2 before BNT162b2 vaccination (day 0) and 1 month after completion of the vaccination schedule (M1).
Materials and methods General and ethical considerationsThe study followed the STROBE reporting guidelines (see checklist in supplemental Table 1, https://links.lww.com/QAD/C554) [17] and was registered with ClinicalTrial.gov (NCT04844489).
The study was approved by ethical and scientific committee of Ile de France number 8 under the registration number IDRCB 2021-A00469–32. All research patients participating in the study were informed and gave written consent for participation. The study obtained CAPNET approval, as it dealt with research in a priority topic during the COVID pandemic. The COVIVAC-ID study is a prospective cohort study of immunocompromised participants vaccinated with BNT162b2 vaccine, including several sub-cohorts defined on the basis of different types of immunosuppression. This report focuses on PWH.
People with HIVWe included PWH followed in the infectious disease departments of the AP-HP Sorbonne Université with a CD4+ T-cell count less than 500 cells/μl and a viral load less than 50 copies/ml, on stable antiretroviral therapy for at least 3 months and eligible for COVID-19 vaccination. The exclusion criteria were a prior known SARS-CoV-2 infection in the preceding 3 months (positive RT-PCR, LAMP PCR or antigen test), vaccination against another disease in the 15 days preceding recruitment, known or suspected allergy to any component of the vaccine or a history of severe allergy, evocative signs of COVID-19, intercurrent infection, pregnancy either underway or planned within the 3 months following vaccination. Two shots of the BNT162b2 vaccine were administered to each patient, 3–6 weeks apart, according to the vaccination schedule recommended in France at the time. Participants were studied before vaccination (day 0), after the first shot but before the second shot (day 28) and 1 month after the last shot (M1). The secondary research objectives, evaluated at the same time, included evaluations of seroconversion rate, the percentage of participants who had been infected before vaccination, changes in antibody [10] level after vaccination, a comparison of the neutralizing capacity of antibodies between the different variants of SARS-CoV-2, and an analysis of the correlation between antibody level and neutralization titer after vaccination, changes in major lymphocyte subpopulations.
SARS-CoV-2 serology, neutralizing antibodiesThe qualitative detection of nucleocapsid protein (anti-N) IgG Abs and the quantification of antispike RBD IgG Abs (SARS-CoV-2 IgG II Quant; Abbott, Rungis, France) were performed with an automated chemiluminescence assay on the Abbott Alinity i platform, in accordance with the manufacturer's instructions. An IgG index at least 0.8 indicates a positive serological result for anti-N antibodies. The cutoff for positivity for anti-S antibodies was 7.1 binding Ab units per milliliter (BAU/ml), as recommended by the manufacturer.
Neutralizing antibodiesA semi-quantitative ELISA (SARS-CoV-2 Surrogate Virus Neutralization Test, Genscript, USA) based on the antibody-mediated blockade of ACE-2–Spike protein interactions was performed as previously described [18]. Briefly, serum samples were incubated with horseradish peroxidase (HRP)-conjugated SARS-CoV-2 RBD and were then added to ACE-2-coated ELISA plates. The RBD-ACE2 interaction was quantified biochemically by adding TMB solution as a substrate. The larger the number of NAbs present in the samples, the greater the inhibition of labeled RBD to ACE-2 observed, resulting in a weaker signal. According to the manufacturer, an inhibition level greater than 30% should be considered a positive result.
The neutralizing activity of the serum samples was also assessed in a whole-virus replication neutralization test (VNT), as previously described [19], with three SARS-CoV-2 isolates: D614G (B.1), and the beta (B.1.351) and delta (B.1.617.2) variants (GenBank accession numbers MW322968 and MW580244 and GISAID accession ID 3431813). Briefly, the serum samples were decomplemented by heat inactivation (56 °C for 30 min), serially diluted and subjected to preincubation, in a 96-well plate, with 50 μl per well of diluted virus, for each virus tested (2 × 103 TCID50/ml), for 60 min at 37 °C. We then added 100 μl of Vero E6 cell suspension (3 × 105 cells/ml) to the mixture, which was then incubated at 37 °C under an atmosphere containing 5% CO2. Microscopy examination was performed on day 4, to assess the cytopathic effect (CPE). NAb titers greater than 1 : 10 were considered positive. The geometric mean titer (GMT) was calculated with a 95% CI for the M1 samples.
Flow cytometryBlood samples were collected into tubes containing EDTA in accordance with the study protocol: absolute numbers of cells were monitored for lymphocyte subsets at each patient visit: day 0, day 28 and M1. Blood cell subset [CD3+, CD4+, CD8+ T lymphocytes, CD19+ B lymphocytes and CD3-CD56+ natural killer (NK) cells] counts (cells/μl) and percentages were established from fresh blood samples with CYTO-STAT tetraCHROME kits, Flowcount fluorescent beads and tetra CXP software, on a FC500 cytometer (Beckman Coulter), in accordance with the manufacturer's instructions.
Data collectionAll data were collected on a web-based case report form. The following covariates were collected at inclusion: known history of COVID-19, presence of comorbid conditions, duration of HIV infection, history of opportunistic infections, history of virologic failure, duration of HIV suppression, clinical category of HIV infection, history of illnesses associated with HIV infection, immunovirological status (HIV viral load, CD4+ and CD8+ cell counts), blood cell count, liver and kidney function tests, C-reactive protein level and class of antiretroviral therapy. During follow-up, we also checked for signs of COVID-19 vaccination failure (documented infection with SARS-CoV-2 and variant, if known). All adverse events were also recorded.
Statistical analysis Sample sizeThe sample size of each subcohort of the COVIVAC-ID study was determined based on a hypothesis concerning the primary endpoint measured 28 days after the first vaccine shot. According to the published data available at the start of the study [20], we expected a neutralization rate of 80% against the alpha variant. We calculated that 97 participants would be required to measure this proportion with a precision (half the 95% CI) of 8% in each subcohort.
Descriptive analysisQuantitative variables are expressed as medians and interquartile ranges, and qualitative variables are expressed as numbers and percentages in this study.
Primary and secondary endpoint analysesThe primary endpoint was the proportion of participants with a NAb titer greater than 1 : 10 against the D614G strain, the beta variant or the delta variant, reported as a percentage with a 95% CI (Clopper–Pearson interval). Analyses were performed on the complete-case dataset. No imputation was performed regarding outcomes for PWH with missing data. Subgroups defined on the basis of CD4+ cell count at baseline were identified but the categories for analysis were restricted to CD4+ cell counts above or below 250 cells/μl, because of the limited number of PWH with very low values. For post hoc analyses of PWH according to their COVID-19 history, we restricted the sample of PWH to those with no known SARS-CoV-2 infection (i.e. with no anti-N antibodies at baseline, and with no known clinical history of COVID-19) receiving two shots, compared with PWH receiving only one shot but had a known history of COVID-19. Serological results were not available during follow-up, so participants with an unknown COVID-19 history received two shots of the vaccine. We compared primary and secondary endpoints between subgroups with appropriate tests according to the type of variable and its distribution: Wilcoxon rank-sum tests or Student's t tests for continuous variables, and chi-squared or Fisher's exact tests for categorical variables.
All analyses were performed with R software, version 4.0. All tests were two-tailed, and the threshold for statistical significance was 0.05 for each comparison. No correction was made for multiple comparisons, and the results for the subgroup analysis remain exploratory.
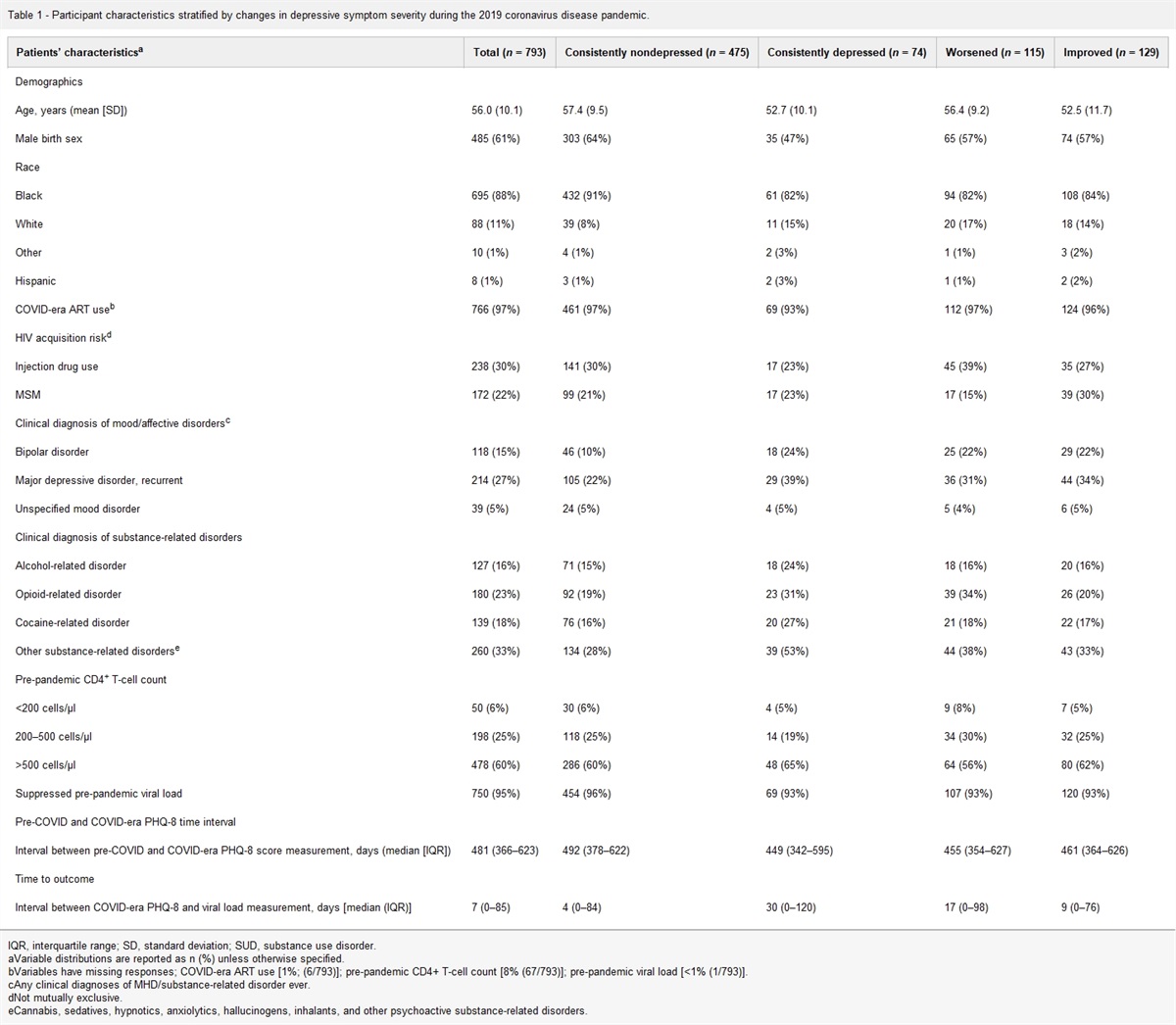
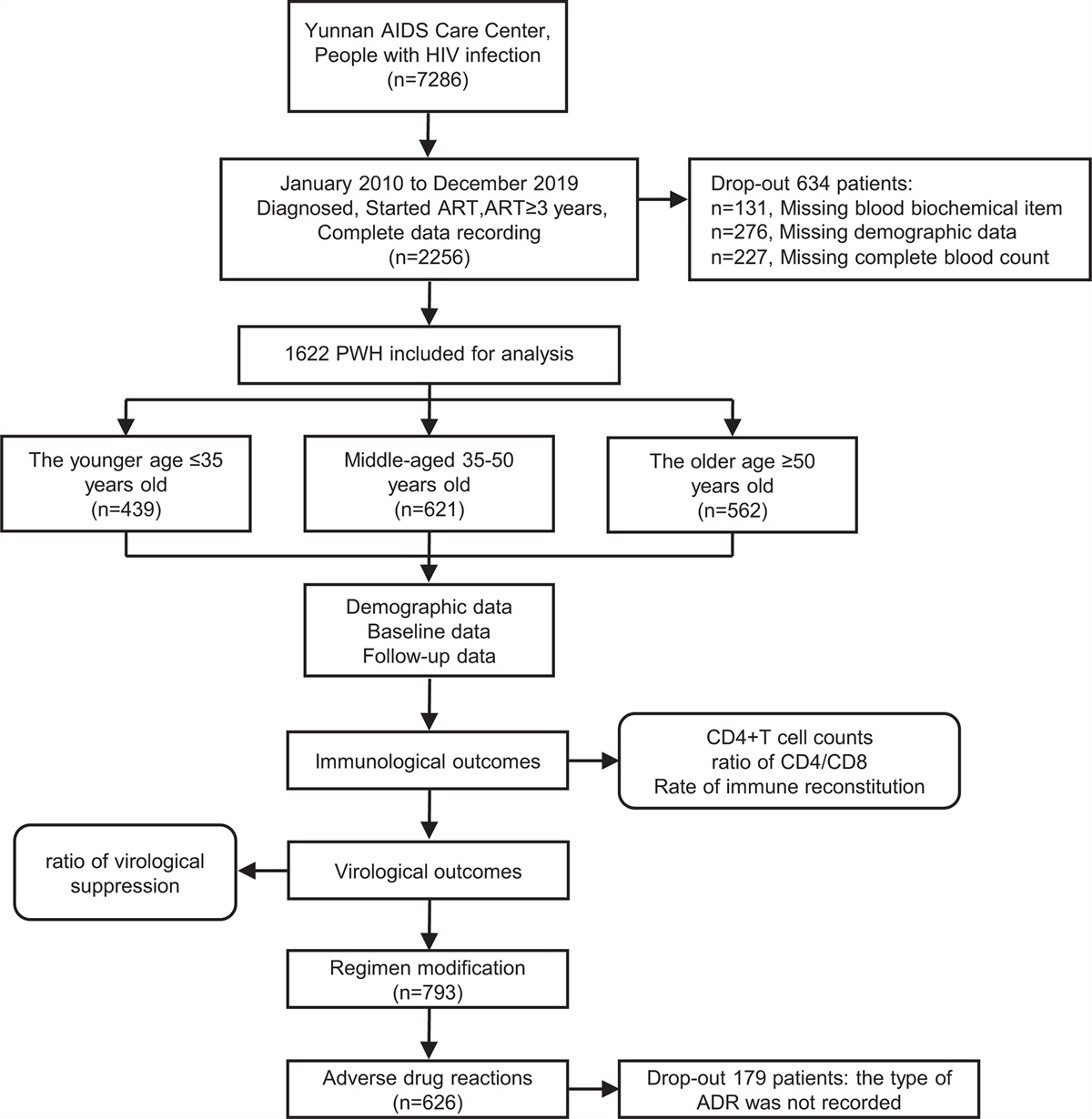
Results People with HIVWe enrolled 97 PWH in this study (see Supplementary Fig. 1, https://links.lww.com/QAD/C554 for flow diagram). Their baseline characteristics are described in Table 1. At the time of vaccination, all PWH had a viral load less than 50 copies/ml on ART, with a median CD4+ T-cell count of 401 cells/μl and a median time since HIV diagnosis of 11 years. Most PWH were on treatment regimens including integrase inhibitors (71%). In total, 85 participants received two shots of vaccine. The remaining 12 PWH received only one shot of vaccine: 11 because of prior symptomatic COVID-19 and one because of early exit from the study. The median time between the two shots was 28 [IQR: 28–29] days. We were able to evaluate 90 PWH at the M1 timepoint. Overall tolerance was very good and no COVID-19 was reported before M1.
Table 1 - Baseline characteristics of people with HIV vaccinated with the BNT162b2 vaccine (n = 97). Median or number IQR or % Age (years) 48 39–56 Sex (male) 85 87.6% BMIa 24 22.0–27.1 Other chronic diseases 39 40.2% Obesity (BMI > 30 kg/m2) 11 11.3% Arterial hypertension 17 17.5% Chronic obstructive pulmonary disease 2 2.1% Diabetes (1 and 2 types) 2 2.1% Allergy 5 5.2% Hemoglobinb 14.8 13.7–15.7 Lymphocytesb 1592 1350-1870 ALATc 25 19–36 ASATc 24 19–30 Duration of HIV infection (years) 11.1 7.6–22.2 Prior history of opportunistic infections 47 48.5% Prior history of virological failure 31 32.0% Duration of ARV therapy (years)d 10.4 6.5–17.4 Duration for which viral load less than 50 copies/ml (months)a 78 28-123 Tritherapy 69 71.1% Bitherapy 22 22.7% CD4+ T cells/μl 401 303–471 CD4+/CD8+ T-cell ratio 0.7 0.4–0.9ARV, antiretroviral drug; IQR, interquartile range.
aOne patient with missing data.
bFive PWH without complete blood counts.
cThirty-seven PWH without liver function test results.
dTwo PWH with missing data.
The SARS-CoV-2 antibody responses observed after the first and second doses are summarized in Table 2. At baseline, 15 of 97 (15.5%) participants had anti-N IgG and 22/97 (23%) had anti-RBD IgG, suggesting prior COVID-19. The seroconversion rate for anti-RBD IgG was 97% (95% CI 90–100%) at M1. Median (IQR) anti-RBD IgG titer was 0.97 (0.97–5.3) BAU/ml at D0 and 1219 (602–1929) at M1 (Fig. 1). Only two participants had not developed anti-RBD IgG by M1. These two participants were aged 56 and 59 years; both were nonobese men with a history of opportunistic infections and virological failure, plasma HIV viral load undetectable for only 4 months and a low CD4+/CD8+ T-cell ratio (0.4 and 0.2).
Table 2 - SARS-CoV-2 antibody response and neutralizing activity before and after BNT162b2 vaccination Day 0 (n = 97) Day 28 (n = 85) Month 1 (n = 90) P value for the test comparing M1 and D0 Positive for anti-N IgG [n (%)] 15 (15.46%) 10 (12.05%) 12 (13.33%) 0.2276c Indeterminatea anti-N IgG [n (%)] 6 (6.19%) 5 (6.02%) 4 (4.44%) Anti-N IgG index [median (IQR)] 0.08 [0.02–0.41] 0.05 [0.02–0.21] 0.07 [0.03–0.36] 0.0013d Positive for anti-S IgG [n (%)] 21 (22.68%) 75 (90.36%) 88 (97.78%) <0.0001c Anti-RBD IgG titer (BAU/ml) [median (IQR)] 0.97 [0.97–5.25] 56.52 [20.80–138.52] 1218.89 [602.12–1969.26] <0.0001d Positive for neutralizing antibodiesb [n (%)] 14 (14.58%) 47 (56.63%) 83 (94.32%) <0.0001cBAU, binding antibody units; IQR, interquartile range.
a0.50 less than anti-N IgG index less than 0.80.
bGenScript ELISA.
cMcNemar's test for paired data.
dWilcoxon test for paired data.
 Fig. 1:
Fig. 1: Change in anti-RBD IgG titer relative to baseline after the first and second doses of BNT162b2 vaccine.
NAb capacity improved between D0 (15%; 95% CI 8–23%) and M1 (94%; 95% CI 87–98%), as shown by the GenScript assay (P < 0.0001). VNT assessments showed that, at M1, NAbs were present in 82% of PWH for D614G, 77% for the beta variant and 84% for the delta variant, with a GMT of 20.6 (95% CI 13–28.1) for D614G, 15.7 (95% CI 11.7–19.8) for the beta variant and 17.3 (95% CI 14.3–20.3) for the delta variant.
The planned subgroup analysis at M1 revealed a seroconversion rate of 91% and a median anti-RBD IgG titer of 852 BAU/ml in participants with CD4+ cell counts less than 250 cells/μl (n = 12) and of 98% and 1270 BAU/ml, respectively, in participants with CD4+ cell counts greater 250 cells/μl (n = 78) (significant difference in the change between D0 and M1 between subgroups; P = 0.0770) (Supplemental Table 2, https://links.lww.com/QAD/C554). At M1, NAbs were detected in 73% of participants with CD4+ cell counts less than 250 cells/μl and 97% of those with CD4+ cell counts greater than 250 cell/μl (P = 0.0130). For D614G and the delta variant, no difference in neutralizing capacity was found between PWH with CD4+ T-cell counts above and below 250 cells/μl. However, for the beta variant, neutralizing capacity was 50% in participants with CD4+ T-cell counts less than 250 cells/μl and 81% in participants with CD4+ T-cell counts greater than 250 cells/μl (P = 0.0292).
Analysis according to history of prior COVID-19 diseaseWe compared the changes in anti-RBD IgG levels and neutralization after vaccination according to the participants’ history of prior COVID-19. We compared participants known to have had COVID-19 before vaccination, who received a single shot of vaccine (n = 11) with participants who received two shots of vaccine and had no anti-N and/or anti-S antibodies, or only extremely low levels of these antibodies at D0 (n = 71). At M1, the participants with a history of prior COVID-19 had higher anti-RBD IgG levels than COVID-19-naive participants: 2345 BAU/ml (2054–4413) versus 1012 BAU/ml (534–1480), respectively (P ≤ 0.0001) (Fig. 2). However, both groups of participants had high neutralization rates at M1: 100% (95% CI 76–100%) for participants with a history of prior COVID-19 and 92% (95% CI 83–97%) for COVID-19-naive participants.
 Fig. 2:
Fig. 2: Change in anti-RBD IgG titers after the second dose of BNT162b2 vaccine as a function of history of prior coronavirus disease 2019.
Major lymphocyte subpopulationsNo changes were observed in CD4+, CD8+ T, and NK cell counts or CD4+/CD8+ T-cell ratio. Vaccination induced the production of antibodies as already reported for healthy individuals but there was also a slight but significant decrease in CD19+ B-cell counts in these PWH (208 ± 124 cells/μl at D0 vs. 188 ± 112 cells/μl at M1, P < 0.01) [21].
DiscussionThis study on PWH with CD4+ T-cell counts less than 500 cells/μl and viral load less than 50 copies/ml showed that a complete BNT162b2 vaccination schedule induced high rates of seroconversion and neutralization of the D614G strain and the delta variant.
Several studies on the immunogenicity of COVID-19 vaccines in various immunocompromised populations have been published in the last few months. However, few data are available for PWH.
Some studies on PWH reported low nonresponse rates, ranging from 0 to 4% [11,22], or antibody titers similar to those for an HIV-negative population [23,24]. Our data confirm the results of a previous study showing that the BNT162b2 mRNA vaccine elicits robust humoral immune responses in PWH [24]. We extend these data here, by showing that the neutralizing activity of antibodies, as determined with the gold-standard VNT assay with a historical strain (B.1 lineage with D614G spike substitution) and VOCs in our population, was not significantly weaker for the delta variant.
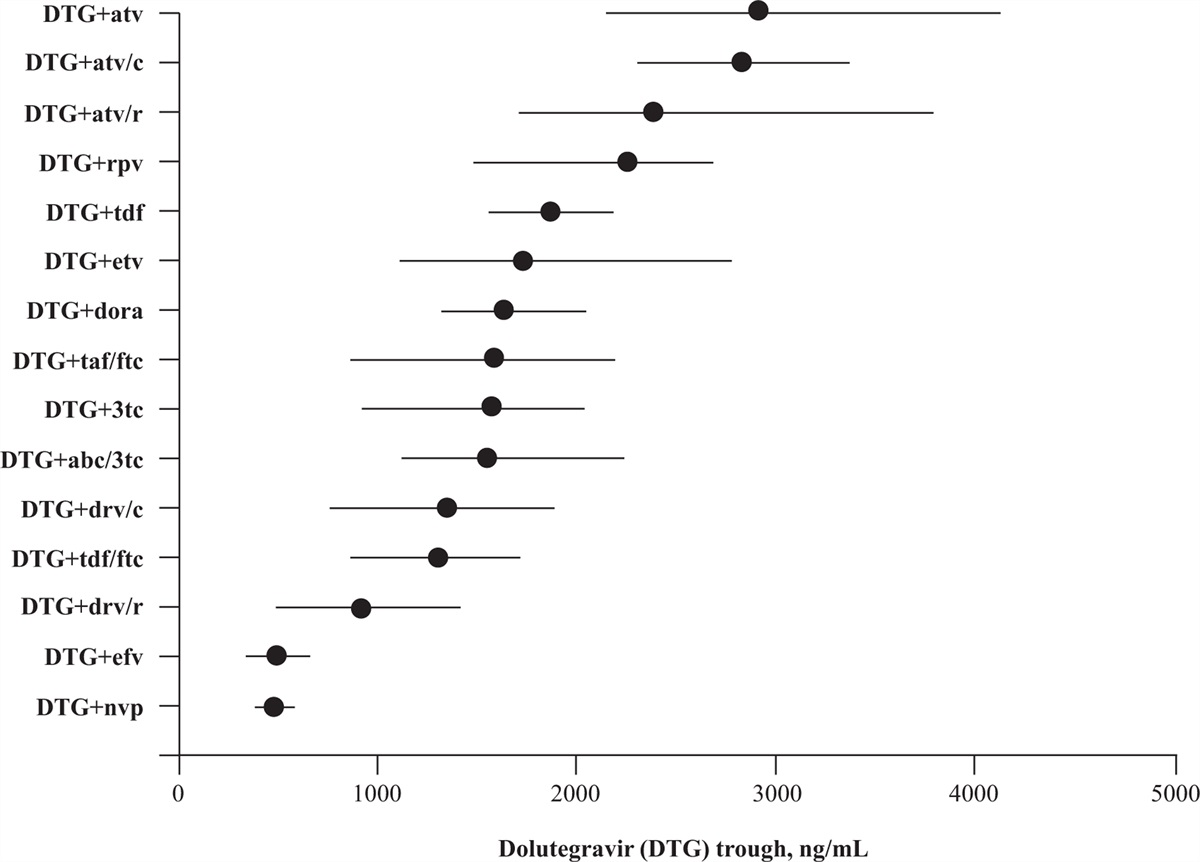
In a recent study, mRNA-containing vaccination schemes, being female individual, and having a higher CD4+ T-cell count were found to be associated with a higher concentration of antibodies in PWH. The antibody response was particularly poor in the subgroup of individuals with CD4+ T-cell counts less than 200 cells/μl [25]. Similarly, an Italian study compared immunogenicity after two shots of mRNA vaccine between PWH with different CD4+ T-cell counts (<200 cells/μl, 200–500 cells/μl and greater than 500 cells/μl) [26]. Regardless of CD4+ T-cell count category, IgG anti-S RBD antibody levels increased, resulting in significant differences between D0 and the time of the second injection and 1 month after the second injection, as in our cohort, with significantly lower levels of antibody in all groups of PWH than in the unpaired control group [26]. In the Yale New Haven Health System, the efficacy of BNT162b2 vaccine was evaluated in PWH aged 55 years and over, at the same time points; 97.5% of PWH displayed a response to the vaccine and CD4+ T-cell count had no impact on vaccine response [27]. The PWH enrolled in our study had CD4+ T-cell counts less than 500 cells/μl. Our data concerning vaccine efficacy in this specific population are reassuring, with no impact of CD4+ T-cell counts on seroconversion rate or median anti-RBD IgG titer after a complete vaccination schedule. However, although PWH with CD4+ T-cell counts less than 250 cells/μl retained the capacity to neutralize both the historical strain and the delta variant, they had a lower neutralizing capacity for the beta variant.
In our study, 11.3% of the PWH had a history of prior COVID-19 infection. This finding is consistent with a previous study conducted in HIV negative workers in Paris France (May–July 2020), which showed that 11% of them had detectable anti-N and anti-S IgG antibodies [28]. Similarly, in three regions in France, including Paris, the rate of seropositivity for anti-SARS-CoV-2 antibodies in adults in May–June 2020, after the first lockdown, was 10% or less; the rate of seropositivity for our cohort of HIV PWH studied 10 months later was in a similar range [29]. Some patients have been infected by COVID-19 prior vaccination in this study related to the number of patients with positive anti-N IgG and anti-S IgG at baseline. The difference between these two parameters could be explained either by a loss of anti-N IgG overtime either a possible access but not declared, to prior vaccination. We found that the antibody responses after vaccination of PWH with prior COVID-19 infection and a single shot of vaccine and of COVID-19-naive PWH who received two shots of vaccine were similar to that in the general population, with higher anti-RBD IgG levels recorded for PWH with a history of prior COVID-19
(Supplemental Figure 2, https://links.lww.com/QAD/C554).
One of the limitations of this study is the relatively small number of participants and the absence of a control group. In addition, the follow-up period is not yet long enough to assess the persistence of antibodies in this population. However, we determined neutralizing activity against different VOCs in PWH.
In conclusion, we show here that PWH with CD4+ T-cell counts less than 500 cells/μl and viral load less than 50 copies/ml have a very good response to the BNT162b2 vaccine, which is unaffected by CD4+ T-cell count. Following a complete BNT162b2 vaccination schedule, these PWH presented high seroconversion rates and a high neutralization capacity for the delta variant.
AcknowledgementsWe thank the COVIVAC-ID staff, including project manager Assitan Kone Haidara and data manager Mehdi Thamri, and Sophie Sayon her technical assistance. The sponsor was Assistance Publique – Hôpitaux de Paris (Direction de la Recherche Clinique et de l’Innovation).
Sources of support: this study was funded by a grant from Assistance Publique – Hôpitaux de Paris. Others analyses (not reported here) in the COVIVAC-ID study are supported by Roche.
Contributors: V.P., L.B., M.R., D.K., V.C., N.S.T., A.G.M. contributed to the conception and design of the study. V.P., L.B., C.S., M.R., S.M., I.M., I.M., K.Z., D.K., V.C., N.S.T., A.G.M. contributed to the acquisition, analysis, and interpretation of data. K.L., N.V., G.P., R.C., R.T., M.A.V. enrolled PWH in the study. V.P., L.B. and A.GM.. wrote the original draft and M.R. and N.S.T. substantively revised the manuscript. All the authors have read and approved the final manuscript.
Conflicts of interestThere are no conflicts of interest.
References 1. Berenguer J, Diez C, Martin-Vicente M, Mican R, Perez-Elias MJ, Garcia-Fraile LJ, et al. Prevalence and factors associated with SARS-CoV-2 seropositivity in the Spanish HIV Research Network Cohort. Clin Microbiol Infect 2021; 27:1678–1684. 2. Lombardi F, Ricci R, Belmonti S, Fabbiani M, Borghetti A, Baldin G, et al. Seroprevalence of SARS-CoV-2 antibodies in HIV-infected PLWHIV in Rome, Italy during the COVID-19 Outbreak. Diagnostics (Basel) 2021; 11:1154. 3. Noe S, Schabaz F, Heldwein S, Mayer W, Ruecker K, Tiller FW, et al. ArcHIV study group. HIV and SARS-CoV-2 co-infection: cross-sectional findings from a German ’hotspot’. Infection 2021; 49:313–320. 4. Spinelli MA, Lynch KL, Yun C, Glidden DV, Peluso MJ, Henrich TJ, et al. SARS-CoV-2 seroprevalence, and IgG concentration and pseudovirus neutralising antibody titres after infection, compared by HIV status: a matched case-control observational study. Lancet HIV 2021; 8:e334–e341. 5. Dong Y, Li Z, Ding S, Liu S, Tang Z, Jia L, et al. HIV infection and risk of COVID-19 mortality: a meta-analysis. Medicine (Baltimore) 2021; 100:e26573. 6. Sarkar S, Khanna P, Singh AK. Impact of COVID-19 in PLWHIV with concurrent co-infections: a systematic review and meta-analyses. J Med Virol 2021; 93:2385–2395. 7. Ssentongo P, Heilbrunn ES, Ssentongo AE, Advani S, Chinchilli VM, Nunez JJ, Du P. Epidemiology and outcomes of COVID-19 in HIV-infected individuals: a systematic review and meta-analysis. Sci Rep 2021; 11:6283. 8. Dagan N, Barda N, Kepten E, Miron O, Perchik S, Katz MA, et al. BNT162b2 mRNA covid-19 vaccine in a nationwide mass vaccination setting. N Engl J Med 2021; 384:1412–1423. 9. Chagla Z. The BNT162b2 (BioNTech/Pfizer) vaccine had 95% efficacy against COVID-19 >/=7 days after the 2nd dose. Ann Intern Med 2021; 174:JC15. 10. Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. C4591001 Clinical Trial Group. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med 2020; 383:2603–2615. 11. Madhi SA, Koen AL, Izu A, Fairlie L, Cutland CL, Baillie V, et al. Wits VIDA COVID team. Safety and immunogenicity of the ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 in people living with and without HIV in South Africa: an interim analysis of a randomised, double-blind, placebo-controlled, phase 1B/2A trial. Lancet HIV 2021; 8:e568–e580. 12. Polvere I, Parrella A, Casamassa G, D’Andrea S, Tizzano A, Cardinale G, et al. Seroprevalence of anti-SARS-CoV-2 IgG and IgM among adults over 65 years old in the south of Italy. Diagnostics (Basel) 2021; 11:483. 13. Garcia-Beltran WF, Lam EC, St Denis K, Nitido AD, Garcia ZH, Hauser BM, et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021; 184:2372.e9–2383.e9. 14. Marot S, Malet I, Leducq V, Abdi B, Teyssou E, Soulie C, et al. Neutralization heterogeneity of United Kingdom and South-African SARS-CoV-2 variants in BNT162b2-vaccinated or convalescent COVID-19 healthcare workers. Clin Infect Dis 2022; 74:707–710. 15. Planas D, Bruel T, Grzelak L, Guivel-Benhassine F, Staropoli I, Porrot F, et al. Sensitivity of infectious SARS-CoV-2 B.1.1. 7 and B. 1. 351 variants to neutralizing antibodies. Nat Med 2021; 27:917–924. 16. Planas D, Veyer D, Baidaliuk A, Staropoli I, Guivel-Benhassine F, Rajah MM, et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021; 596:276–280. 17. von Elm E, Altman DG, Egger M, Pocock SJ, Gotzsche PC, Vandenbroucke JP. STROBE Initiative. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: guidelines for reporting observational studies. J Clin Epidemiol 2008; 61:344–349. 18. Tan CW, Chia WN, Qin X, Liu P, Chen MI, Tiu C, et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat Biotechnol 2020; 38:1073–1078. 19. Marot S, Malet I, Leducq V, Zafilaza K, Sterlin D, Planas D, et al. Rapid decline of neutralizing antibodies against SARS-CoV-2 among infected healthcare workers. Nat Commun 2021; 12:844. 20. Xie X, Zou J, Fontes-Garfias CR, Xia H, Swanson KA, Cutler M, et al. Neutralization of N501Y mutant SARS-CoV-2 by BNT162b2 vaccine-elicited sera. bioRxiv 2021. 21. Fu Y, Chen F, Cui L, Zhao Y, Zhang H, Fu S, et al. Immunological analysis of people in Northeast China after SARS-CoV-2 inactivated vaccine injection. Vaccines (Basel) 2021; 9:1028. 22. Levy I, Wieder-Finesod A, Litchevsky V, Biber A, Indenbaum V, Olmer L, et al. Immunogenicity and safety of the BNT162b2 mRNA COVID-19 vaccine in people living with HIV-1. Clin Microbiol Infect 2021; 27:1851–1855. 23. Frater J, Ewer KJ, Ogbe A, Pace M, Adele S, Adland E, et al. Oxford COVID Vaccine Trial Group. Safety and immunogenicity of the ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 in HIV infection: a single-arm substudy of a phase 2/3 clinical trial. Lancet HIV 2021; 8:e474–e485. 24. Woldemeskel BA, Karaba AH, Garliss CC, Beck EJ, Wang KH, Laeyendecker O, et al. The BNT162b2 mRNA vaccine elicits robust humoral and cellular immune responses in people living with HIV. Clin Infect Dis 2021. 25. Noe S, Ochana N, Wiese C, Schabaz F, Von Krosigk A, Heldwein S, et al. Humoral response to SARS-CoV-2 vaccines in people living with HIV. Infection 2021; 50:617–623. 26. Antinori A, Cicalini S, Meschi S, et al. Immunogenicity of mRNA vaccination against SARS-CoV-2 in persons living with HIV (PLWHs) with low CD4 count or previous AIDS. 18th European AIDS Conference, EACS 2021, October 27–30, 2021, London. Abstract OS4/3. 27. Tuan JJ, Zapata H, Critch-Gilfillan T, Ryall L, Turcotte B, Mutic S, et al. Qualitative assessment of anti-SARS-CoV-2 spike protein immunogenicity (QUASI) after COVID-19 vaccination in older people living with HIV. HIV Med 2021; 23:178–185. 28. Anna F, Goyard S, Lalanne AI, Nevo F, Gransagne M, Souque P, et al. High seroprevalence but short-lived immune response to SARS-CoV-2 infection in Paris. Eur J Immunol 2021; 51:180–190. 29. Carrat F, de Lamballerie X, Rahib D, Blanche H, Lapidus N, Artaud F, et al. or the SAPRIS and SAPRIS-SERO study groups. Antibody status and cumulative incidence of SARS-CoV-2 infection among adults in three regions of France following the first lockdown and associated risk factors: a multicohort study. Int J Epidemiol 2021; 50:1458–1472.
留言 (0)