
記住我




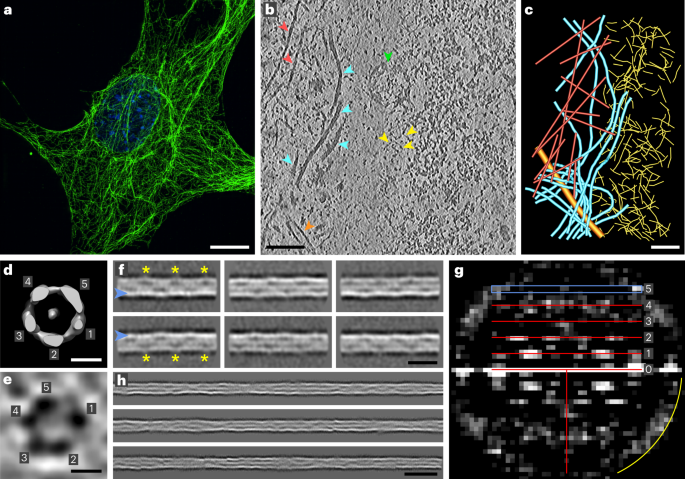
A cryo-EM reconstruction of a recombinant human CRL7FBXW8 complex, prepared by coexpressing CUL7, RBX1, SKP1 and FBXW8 in human human embryonic kidney 293S (HEK293S) cells, refined to 2.8 Å resolution overall. The density allowed fitting previous structures of RBX1 and SKP1, and building atomic models of CUL7 and FBXW8 de novo. The final coordinates were generated after multiple cycles of manual rebuilding and refinement (Fig. 1, Table 1, Extended Data Fig. 1 and Extended Data Table 1). Although AlphaFold was released subsequent to structure determination, it is noteworthy that most of the individual domains in CUL7 and FBXW8 superimpose with those predicted with Cα-r.m.s.d. values of <1.3 and <0.8 Å, respectively (Extended Data Fig. 2)34.
Fig. 1: Structure of CRL7FBXW8.
a, Cryo-EM map of CRL7FBXW8 (calculated with DeepEMhancer54). b, Structure of CRL7FBXW8 in the orientation of the cryo-EM map in a. c, Structure showing substrate ubiquitination by CRL1β-TRCP and E2 (PDB 6TTU)35, with SKP1 aligned in the orientation of CRL7FBXW8 in b. d, WD40 domain and C-terminal extension of FBXW8. Blades are labeled and color coded from 1–8 and a–d strands are labeled on blade 8.
Table 1 Cryo-EM data collection, refinement and validation statisticsCRL7FBXW8 adopts a diagonal T-shaped structure. CUL7–RBX1 forms a ≅160 Å long bar that crosses the ≅115 Å long CUL7-FBXW8–SKP1 base of the ‘T’ at a ≅50° angle (Fig. 1a). The T-junction is formed by five domains from CUL7 wrapping around FBXW8’s WD40 β-propeller and stalk helix (Fig. 1b). SKP1’s N-terminal region projects away from CUL7–RBX1 and was poorly resolved in the maps, but was visualized at the edge of the complex by docking previous SKP1–F-box protein complex structures into the low-resolution density (Extended Data Fig. 3a). Overall, the structure is well resolved, allowing unambiguous placement of RBX’s RING domain and building of residues for most of the protein complex (Extended Data Fig. 3b–e).
The CRL7FBXW8 T-shape differs from the more ovoid assemblies formed by canonical CRLs (Fig. 1c). CUL7 does not directly bind SKP1. This is explained by CUL7 lacking a cullin-repeat 1 (CR1) domain that is used by cullins 1–5 to bind their cognate substrate receptor modules. Notably, CR1 domain interactions are central to the characteristic architecture of prototypic CUL1-based E3 ligases6. CUL1–RBX1 forms half an oval. At one end, CUL1’s CR1 domain and SKP1 curve around to place the F-box protein’s substrate-binding domain facing RBX1 bound at the other end of CUL1. The oval is closed when RBX1’s RING domain binds a ubiquitin-carrying enzyme to deliver ubiquitin to an F-box protein-bound substrate35,36 (Fig. 1c). Below, we describe the distinct structural details of CRL7FBXW8, and how E3 ligase activity can be achieved despite its unique assembly.
SKP1–FBXW8 resembles canonical SKP1-FBXW assembliesWithin CRL7FBXW8, although SKP1 does not contact with CUL7, the SKP1–FBXW8 subcomplex shows all the hallmark features of canonical SKP1 complexes with WD40 domain-containing F-box proteins: (1) the compact three-helix F-box is encased by SKP1’s C-terminal helices (Extended Data Fig. 4a); (2) a ≅30 Å long stalk helix rigidly projects from the F-box and (3) the stalk helix continues into a β-propeller (Extended Data Fig. 4b,c). FBXW8’s eight-bladed β-propeller overall superimposes well with those of other F-box proteins, most notably human FBXW7 (Protein Data Bank (PDB) 2OVP) and yeast Cdc4p (PDB 1NEX) (Extended Data Fig. 4b)37,38. As is typical of WD40 proteins, the β-propeller is closed by the N-terminal strand completing the otherwise C-terminal-most blade (Fig. 1d). However, FBXW8 is unique in that its C-terminal ≅35 residues extend beyond the propeller to form a triangular platform between the propeller and stalk helix. This C-terminal structure makes three-way interactions between FBXW8’s first propeller blade and two disparate CUL7 domains (Fig. 1b).
Unique domains and cullin-RING arrangement in CUL7–RBX1The CUL7–RBX1 portion of the structure contains four distinct regions (Fig. 2a,b). CUL7’s atypical N-terminal region and C-terminal cullin region bound to RBX1 together form the elongated bar shape that crosses the ‘T’. CUL7’s N-terminal region comprises a unique collection of domains that cap the N-terminal edge of the cullin region. A small beta-domain (SBD) connects via a poorly resolved 50-residue linker to the first of three short armadillo repeat units (ARM1, ARM2 and ARM3). The ARM3 unit covers the seam between the ARM1 and ARM2 domains, and proceeds into the first helix of the cullin region. The junction between the N- and C-terminal regions is mediated by a 64 Å long bent helix that is both the last helix of the ARM3 unit and the first helix of the cullin region. Altogether, the SBD, ARM1, ARM2 and ARM3 domains pack against each other to wrap around and completely encircle this junction helix. The N-terminal-most domain in the cullin region (the CR2 domain) is embraced on one side by the SBD and on the other by the ARM2 domain (Fig. 2b).
Fig. 2: Architecture of CUL7–RBX1.
a, Domains of CUL7 and RBX1. b, Structure of CUL7–RBX1 from CRL7FBXW8 (excluding FBXW8–SKP1 for a clearer view of CUL7), with domains colored according to a. c, Superposition of CUL7 and CUL1 (PDB 1LDJ)6 aligned on the WHB domain showing closeup of α/β-to-WHB linker region. d, Superposition of CUL7 and CUL1 (PDB 1LDJ) aligned on WHB domain showing relative orientations of RBX1 RING domains in CRL7FBXW8 and CUL1–RBX1 (PDB 1LDJ)6. e, C terminus of RBX1 anchored to WHB and 4HB of CUL7 shown in DeepEMhancer map.
CUL7’s cullin region initiates with CR2 and CR3 domains, which are five-helix bundles arranged in tandem, and proceeds with a four-helix bundle (4HB), a mixed α-helix/β-sheet (α/β) region, an α/β-to-WHB linker and the C-terminal WHB domain (Fig. 2c,d and Extended Data Fig. 5a). Individually, most of these CUL7 domains resemble those in other cullins6 (Extended Data Fig. 5a). However, CUL7’s α/β-to-WHB linker differs in such a way that prevents neddylation as described in detail below.
RBX1’s N-terminal region forms a β-strand, which is inserted in CUL7’s α/β region. Together, these CUL7 and RBX1 elements form an intermolecular cullin/RBX (C/R) domain that requires both proteins for proper folding. However, the relative orientation of RBX1’s ensuing RING domain differs from the model arrangement in CUL1–RBX1 (Fig. 2d). RBX1’s ten C-terminal residues extend beyond the RING domain, fill a crevice between CUL7’s 4HB and WHB domains, and cement a CRL7-specific domain arrangement (Fig. 2e). Although the WHB domain is the C terminus of canonical cullins, CUL7 has an additional 50 residues not overtly visible in the cryo-EM density. It is tempting to speculate that CUL7’s C terminus corresponds to patchy cryo-EM density covering RBX1’s RING domain, although the map does not allow unambiguous attribution of this region (Extended Data Fig. 3b).
The two other structured domains from CUL7—the ARM2-Ins (an extended loop insertion in ARM2) and the DOC domain—both bind FBXW8 (Fig. 3a). The CPH domain is not visible in the cryo-EM maps of CRL7FBXW8 and is presumably flexibly tethered to the ARM2 domain.
Fig. 3: CUL7-FBXW8 interface.
a, Structure of CRL7FBXW8 with CUL7 domains colored as in Fig. 2 and interfaces between CUL7 and FBXW8 highlighted in squares. FBXW8 is colored in a gradient to visualize the individual blades 1–8, the stalk helix and the C-terminal extension. b–d, Close-ups of CUL7-FBXW8 interfaces with corresponding cryo-EM density calculated with DeepEMhancer. b, Interface between CUL7 4HB and FBXW8 blades 5 and 6. c, Interface between FBXW8 C-terminal extension with CUL7 ARM2-Ins and DOC domain, between CUL7’s CR2 and CR3 domains, and an extended loop from FBXW8 blade 8. d, Interface between CUL7 DOC domain and FBXW8 stalk helix and C-terminal extension.
Unique cullin-F-box protein assemblyCUL7 binds FBXW8 through an elaborate multipart interface that differs entirely from CUL1 interactions with F-box proteins. First, the concave surface from CUL7’s CR2, CR3 and 4HB domains encases roughly half the rim of the top side of FBXW8’s β-propeller (Fig. 3a). Grooves between CUL7 CR3 and 4HB domain helices embrace intra- and inter-blade loops from blades 5–8 of FBXW8 (Fig. 3b, c). Extended loops between the central strands of blades 5 and 8 secure the two edges of this interface. Second, CUL7’s ARM2-Ins protrudes nearly 40 Å from the ARM2 domain, fills the groove between CUL7’s CR2 and CR3 domains, meanders around the crevice between the FBXW8’s β-propeller and C-terminal extension, and then returns to complete the ARM2 domain (Fig. 3c). Finally, CUL7’s DOC domain nestles in a furrow between the stalk helix and C-terminal extension of FBXW8 (Fig. 3d). The atypical assembly explains why CUL7 uniquely binds FBXW8 but no other F-box proteins: the interactions are with domains, domain insertions and sequences unique to FBXW8, and even those with the WD40 domain involve residues that are not conserved even with its closest homolog FBXW7 (Extended Data Fig. 5b,c).
The structure also provides a rationale for why our purification of FBXW8 required coexpression not only with SKP1 to bind its F-box, but also CUL7–RBX1 (Methods). The interface is highly hydrophobic; it seems likely that exposure of the hydrophobic surfaces would lead to aggregation of SKP1–FBXW8 purified on its own. In total, 85 residues from CUL7 interact with 91 residues—nearly 20%—from FBXW8 to bury roughly 3,500 Å2 of surface area (Extended Data Fig. 5d).
Comparing interface residues between the cryo-EM structure and those predicted using AlphaFold raises the possibility some key interacting regions in CUL7 and FBXW8 may fold and/or rearrange to form the interaction. In particular, the differences from the models suggest the CUL7 ARM2-Ins and DOC domain, and FBXW8 blade 5 and blade 8 loops and the C-terminal extension are considerably remodeled and/or reoriented in the complex (Extended Data Fig. 2b).
Purified recombinant CRL7FBXW8 assembly is inactiveCanonical CRLs exhibit a wide range of E3 ligase activities, in partnership with various NEDD8- and ubiquitin-linked carrying enzymes. These activities depend on multivalent interactions, at minimum involving the RBX RING domain, an E2 and its active site-linked NEDD8 or ubiquitin, and also other contacts, for example between a cullin-linked NEDD8 and a ubiquitin-carrying enzyme35,36,39. The structure of CRL7FBXW8 is inconsistent with even the most rudimentary of these canonical CRL NEDD8 and ubiquitin E3 ligase activities. A previous structure showed how CUL1 is neddylated on a specific lysine in the WHB domain40. Comparing the structure of CRL7FBXW8 showed two features that prevent it from attaining the arrangement required for neddylation despite CUL7’s WHB domain displaying a lysine corresponding to the CUL1 neddylation site. First, a long, rigid ‘Helix29’—which is conserved in structures of unneddylated cullins 1–5—acts like a wand to direct the targeted lysine into the active site of the RBX1-E2~NEDD8 intermediate (Fig. 4a)40. However, CUL7 lacks this long, rigid Helix29. Instead, the corresponding region forms the α/β-to-WHB linker, an extended amphipathic structure not found in the other cullins (Figs. 2a,c and 4a,b). The hydrophobic face of the α/β-to-WHB linker is embedded in a hydrophobic surface from CUL7’s 4HB domain (Fig. 4b,c). These interactions, along with additional contacts between CUL7’s WHB domain with the 4HB domain and with RBX1’s RING domain, fixes CUL7’s WHB domain in an orientation incompatible with neddylation (Fig. 4d–e). Second, previous studies also showed that neddylation requires a specific RBX1 RING-domain orientation. This is required to bind and position the E2~NEDD8 intermediate with its active site adjacent to the cullin’s target lysine (K720 in CUL1, Fig. 4d)41. However, in CRL7FBXW8, RBX1’s RING domain is secured in an inactive arrangement by intercalation of its C-terminal residues between CUL7’s 4HB and WHB domains (Fig. 2e). The position of RBX1’s RING domain in CRL7FBXW8 prevents binding an active E2~NEDD8 intermediate as shown in a model based on the structure visualizing CUL1 neddylation (Fig. 4e). The arrangement in CRL7FBXW8 also prevents RBX1’s RING domain from binding an active E2~ubiquitin intermediate as shown in a model based on a structure visualizing CRL1 substrate ubiquitination (Extended Data Fig. 6a,b).
Fig. 4: CUL7 is not neddylated under conditions allowing efficient neddylation of canonical cullins.
a,b, Structural arrangement of α/β-to-WHB linker region comparing CUL1 (PDB 4P5O)40 in a with CRL7FBXW8 in b, views are aligned on α/β domain. PDB 4P5O required use of a CUL1 version with the neddylation site K720 mutated to arginine to prevent transfer of NEDD8 and visualize the catalytic architecture. K720 was modeled in the structure here for clarity. c, Interface between CUL7’s α/β-to-WHB linker region and 4HB shown in cryo-EM map calculated with DeepEMhancer. d, Two views of structure visualizing CUL1 neddylation (PDB 4P5O). e, Modeling activated RBX1 RING–UBE2M~NEDD8 (PDB 4P5O) shows this clash with CRL7FBXW8, depicted by aligning RBX1 RING domain from both structures. f, In vitro neddylation assays using the indicated cullin-RING complex and NEDD8 E2 (UBE2M or UBE2F), NEDD8 E1 (NAE1-UBA3) and detecting fluorescently labeled NEDD8 in SDS–PAGE gels (samples of both gels derived from the same experiment and gels were processed in parallel, n = 2 technically independent experiments).
E3 ligase activities have not been reconstituted with biochemically pure CRL7FBXW8. We therefore tested activity with our purified recombinant components. In canonical CRLs, RBX1 promotes NEDD8 transfer to the cullin’s WHB domain lysine from either neddylating E2 UBE2M or UBE2F, while the homologous RBX2 only functions with UBE2F (ref. 42). Examining neddylation side-by-side with canonical cullin–RBX complexes showed that in control reactions, by the first time-point, fluorescent NEDD8 was fully transferred from UBE2M or UBE2F to CUL1–RBX1, and from UBE2F to CUL5–RBX2. However, CRL7FBXW8 was not substantially modified by either NEDD8 E2 (Fig. 4f and Extended Data Fig. 6c).
We next probed automodification as a readout for intrinsic ubiquitin E3 activity. Under conditions when the control canonical CUL–RBX1 complexes (wild-type or neddylated) were autoubiquitinated in the presence of E1 (UBA1) and E2 (UBE2D3), CRL7FBXW8 was relatively inactive, similar to the nonneddylatable CUL1 K720R mutant (Extended Data Fig. 6d). One possibility could be that CRL7FBXW8 preferentially uses an alternative E2. However, autoubiquitination was also not observed with 26 tested E2s (Extended Data Fig. 6e).
In complexes with neddylated canonical cullins, RBX1’s RING domain can also promote ubiquitin chain formation by UBE2G1 and by UBE2R-family E2s43. RBX1 associated with unneddylated canonical cullins can also promote such polyubiquitination with UBE2R1 (ref. 44), which is stimulated by interactions between an acidic C-terminal region of UBE2R1 and a basic canyon on canonical cullins45 that is notably lacking in CUL7 (Extended Data Fig. 6f). These activities can be assayed even in the absence of a substrate, by monitoring transfer of fluorescent ubiquitin from a preformed E2~ubiquitin intermediate to an unlabeled acceptor ubiquitin (~ here refers to the thioester linkage between E2 catalytic cysteine and ubiquitin C terminus). However, CUL7–RBX1 was inactive under conditions when RBX1 complexes with CUL1 promote di-ubiquitin synthesis (Extended Data Fig. 6g). Taken together, the results are consistent with the structural findings that RBX1’s RING domain in CRL7FBXW8 is hindered from binding an active E2~NEDD8 or E2~ubiquitin intermediate.
FBXW8–SKP1 bridges CUL7–RBX1 and CUL1–RBX1In vivo, CUL7 was found to associate with CUL1 in an FBXW8-dependent manner18. The CRL7FBXW8 structure suggests the mechanism: the canonical CUL1-binding residues in CUL7-bound SKP1 and the F-box are fully exposed and thus could mediate the interaction of CUL7 with CUL1. Comigration in size-exclusion chromatography (SEC) showed CRL7FBXW8 directly binds full-length CUL1–RBX1 (Fig. 5a), but not an N-terminally truncated version lacking the CR1 domain (Extended Data Fig. 7a). Furthermore, when bound to CRL7FBXW8, CUL1–RBX1 is excluded from binding another SKP1–F-box protein complex (harboring a monomeric version of the F-box protein β-TRCP1) (Extended Data Fig. 7b–d). Indeed, a cryo-EM map of a complex between CRL7FBXW8 and the N-terminal region of CUL1 (comprising the CR1, CR2 and CR3 domains), refined to a resolution of 4.6 Å visualized the interactions: SKP1–FBXW8 within the CRL7FBXW8 complex binds CUL1’s CR1 domain in a canonical manner (Fig. 5b).
Fig. 5: CRL7FBXW8 forms an active E3 ligase with NEDD8–CUL1–RBX1.
a, SEC experiments comparing migrations of CRL7FBXW8, CUL1–RBX1 and the mixture of both through a Superose 6, 5/150GL column, visualized by total absorbance (280 nm) and the corresponding elution fractions on Coomassie stained SDS–PAGE gels. b, Cryo-EM map of CRL7FBXW8–CUL1 N-terminal region (residues 1–410) in two orientations. c, In vitro assay showing TP53 ubiquitination by CRL7FBXW8 and neddylated CUL1–RBX1. Assays detected TP53 with TAMRA appended at the N terminus, and contained indicated versions of CUL1, CUL5 and CUL7 complexes, E1 (UBA1), E2 (UBE2D3) and ubiquitin (n = 2 technically independent experiments). d, Model of the active E3 ligase formed by interaction between CRL7FBXW8 and neddylated CUL1–RBX1. N8, NEDD8; UB, Ubiquitin.
CRL7FBXW8 can recruit a neddylated CRL1 substrate in vitroGiven that CRL7FBXW8 lacks E3 activity in our assays, but it binds CUL1–RBX1, we asked if CRL7FBXW8 could in principle serve in a substrate receptor manner for a neddylated CRL1 complex. If this were the case, then a CRL7FBXW8-interacting protein could potentially serve as a substrate in vitro. The only such interaction that has been reconstituted with pure proteins is CUL7’s CPH domain binding to TP53. Indeed, we found that CRL7FBXW8, TP53 and CUL1–RBX1 can form a complex based on comigration by SEC, and this depends on CUL7’s TP53-binding CPH domain (Extended Data Fig. 7e–j). The CPH domain is connected to the ARM2 domain by equal to or roughly 45- and 28-residue long linker sequences, which are presumably flexible given that this domain is not observed in the cryo-EM map. In principle, the CPH domain—which itself spans equal to or roughly 25 Å—could extend over a wide radius relative to the rest of the complex to deliver TP53 to the ubiquitination active site (Extended Data Fig. 8a).
Previously, we found that immunoprecipitated CUL7 was shown to mediate mono- or oligoubiquitination of TP53 (ref. 21). Notably, a CUL7 mutant unable to bind RBX1 showed similar activity21. We were able to reconstitute such TP53 ubiquitination using entirely purified components, dependent on CRL7FBXW8 and neddylated CUL1–RBX1 (Fig. 5c, lanes 1–4). We thus, used TP53 as model substrate to probe mechanistic implications of the CRL7FBXW8–CUL1–RBX1 complex. Several observations are consistent with CUL7 recruiting TP53 for neddylated CUL1–RBX1-mediated ubiquitination. First, ubiquitination depends on CUL7’s TP53-binding CPH domain (Fig. 5c, lane 5). Second, neddylated CUL5–RBX2, which does not bind SKP1–F-box protein substrate receptor modules, cannot substitute for neddylated CUL1–RBX1 (Fig. 5c, lane 6). Finally, the CRL7FBXW8-dependent TP53 ubiquitination is decreased with catalytically impaired neddylated CUL1–RBX1 mutants (nonneddylatable CUL1 K720R, Fig. 5c, lane 7), or CUL1–RBX1 modified with Q40E mutant NEDD8 (the Q40E mutation blocks noncovalent interactions between NEDD8 and CUL1 that allosterically activate CUL1)35 (Fig. 5c, lane 8). Future studies will be required to investigate potential cellular functions of TP53 ubiquitination by the distinctive CRL7FBXW8-neddylated CUL1–RBX1 assembly. Nonetheless, the data demonstrate that CRL7FBXW8 can biochemically act as a CRL1 substrate receptor (Fig. 5b,c and Extended Data Fig. 8a,b).
3M syndrome mutationsThe physiological importance of CUL7 is underscored by its mutation causing developmental defects resulting in the human hereditary short stature disorder 3M syndrome23. CUL7 mutations account for 70% of 3M syndrome cases, with the remainder caused by mutations in OBSL1 and CCDC8 occurring in a mutually exclusively manner between these three genes. CUL7 interacting with OBSL1 and CCDC8 may be compatible with CUL7 incorporation into CRL7FBXW8 or binding to TP53 (refs. 15,17,21,23,24,46,47), although it is unclear if such interactions are functionally related to the 3M complex since neither FBXW8 nor TP53 are mutated in the disease (Extended Data Fig. 8c). Nonetheless, the CRL7FBXW8 assembly allows obtaining insights into potential structural effects of 3M syndrome mutations. Mapping mutations onto the structure shows that most are premature truncations that would affect CUL7 binding to RBX1 and presumably formation of the 3M complex, as this depends on C-terminal regions of CUL7 binding to OBSL1 (ref. 15). Although missense mutations map throughout the structure, several do map to interprotein interfaces in CRL7FBXW8, including with FBXW8 (P861S in the DOC domain, and Q1246G in the 4HB) and RBX1 (W1448R, G1452D, H1464P, Q1469R in the α/β-domain), as well as the interdomain interface between the CR2 and CR3 domains (L1014R) (Extended Data Fig. 8d,e). In agreement with the structure, mutation of H1464 and Q1469 located in CUL7’s β-sheet that binds RBX1’s N terminus, or the frame-shift at E1594 that truncates the RBX1-binding region of the WHB domain, were shown to eliminate binding to RBX1 (ref. 24). Thus, although FBXW8 has not been found to be mutated in 3M syndrome patients, the CUL7 mutations at the interface with FBXW8 may suggest a role in the function of the 3M E3 ligase complex.
留言 (0)