
記住我



The method and interest in using network description of tissues is clearer when the library is applied to specific cases. Below we provide some illustrations of the implementation of Griottes on three types of inputs: 2D and 3D images as well as a data table. These examples also correspond to different biological samples: the adult zebrafish pallium and mesenchymal stem cell spheroids. Each example showcases a different use case for the program.
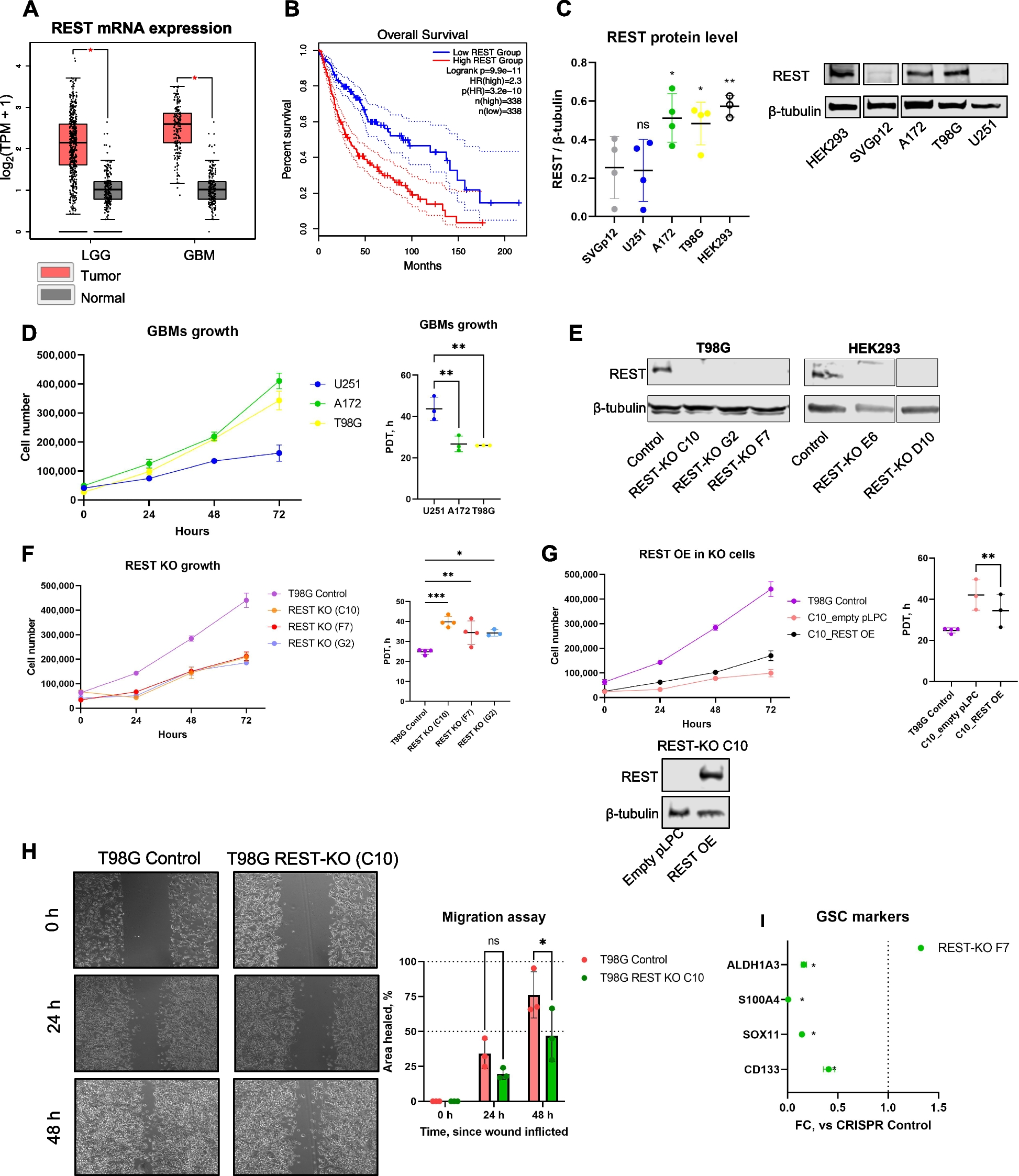
Zebrafish telencephalon 2D imageDeciphering the generation of neural stem cells (NSCs) in the adult zebrafish brain requires a deeper understanding of the connections between individual cells in the tissue. High-quality in vivo images have recently been obtained by Dray et al. [20], as shown in Fig. 2a. This image consists of a projected 3D stack of whole-mount immunostained pallium from 3 months post-fertilization zebrafish. Visualization of GFAP (glial fibrillary acidic) expression allows the identification of the neural stem cells, while PCNA (proliferating cell nuclear antigen) highlights dividing cells and ZO1 (Zonula Occludens 1) shows the apical domain of the cells. These cells have been shown to coordinate their behavior in part via local cell-cell interactions (see Ref.[20] for details).
The image shown in Fig. 2a is first segmented using Cellpose [21], which yields a multi-channel 2D image containing the masks resulting from the cell segmentation and the fluorescence image in the GFAP and PCNA channels. This image is then analyzed using Griottes to generate a network representation of the tissue, as shown in Fig. 2b. Different network construction methods generate different graphs. This feature is illustrated in Fig. 2b, where a graph is constructed for a sub-section of the tissue using the contact, the Delaunay and the geometric construction rules. For the geometric and Delaunay graphs, the maximum cell-cell distance authorized is 15 μm, to avoid generating spurious connections between cells. While the Delaunay and contact graphs give broadly similar structure, the geometric graph shows clear differences in neighborhood, due to the heterogeneous sizes of the cells and their arrangements. The differences in graph structure can be measured quantitatively, e.g., by observing the degree distribution (number of links per cell) as a function of the network construction method. The distribution of degrees in the geometric graph is much broader than in the contact of Delaunay graphs (Fig. 2c).
From the input image, Griottes can also extract the fluorescence in the PCNA channel of the individual cells in the tissue. The distribution of fluorescence intensities is shown in Fig. 2d. A threshold value is then selected manually in order to classify cells with a higher intensity as cell type PCNA+ and those below as cell type PCNA− (Fig. 2e). This binary classification now allows us to set the cell type as an attribute for each node and construct the network representation of the tissue (Fig. 2f). We verify that the brightest cells in the PCNA channel are indeed classified as PCNA+ (Fig. 2f).
In images where the whole cells are segmented, as is the case here, it is possible to include the individual cell-cell contact surfaces in the network representation of the tissue. Indeed, cells can have different shapes leading to a high heterogeneity of cell contact surfaces within the tissue (Fig. 2g). Cell-cell contact can be pertinent to understand tissue behavior, since both chemical and mechanical signals are transmitted through membrane contacts. Extracting properties of these contact surfaces can shed light on key biological processes such as notch signaling [22]. Griottes allows easy retrieval of the contact length between all pairs of neighboring cells. The size of the contacts can be represented visually for each link (Fig. 2g) and the quantitative distribution of these values can be obtained (Fig. 2h) for downstream analysis.
Analysis of 3D spheroid imagesThe network representation can also be applied to 3D cultures, in the form of spheroids or organoids. These model systems recapitulate many aspects of the in vivo tissue structure, as shown, e.g., by comparing cryo-sections of organs with organoids [7, 23]. In these tissue models, the structural organization in 3D was shown to couple back on the functional behavior of the cells within the organoid. This was quantitatively shown in the case of mesenchymal stromal cells (MSCs), which were found to organize in a hierarchical manner, depending on their level of commitment, and to express different levels of chemokines depending on their position in the organoid [24, 25]. Previous analysis however was limited to coarse-grained quantities on averaged 2D images [26, 27]. This can now be overcome by imaging the organoids using a light-sheet microscope, which provides very good resolution over the complete depth of the organoid. Some representative slices of the 3D reconstruction are shown in Fig. 3a.
Fig. 3
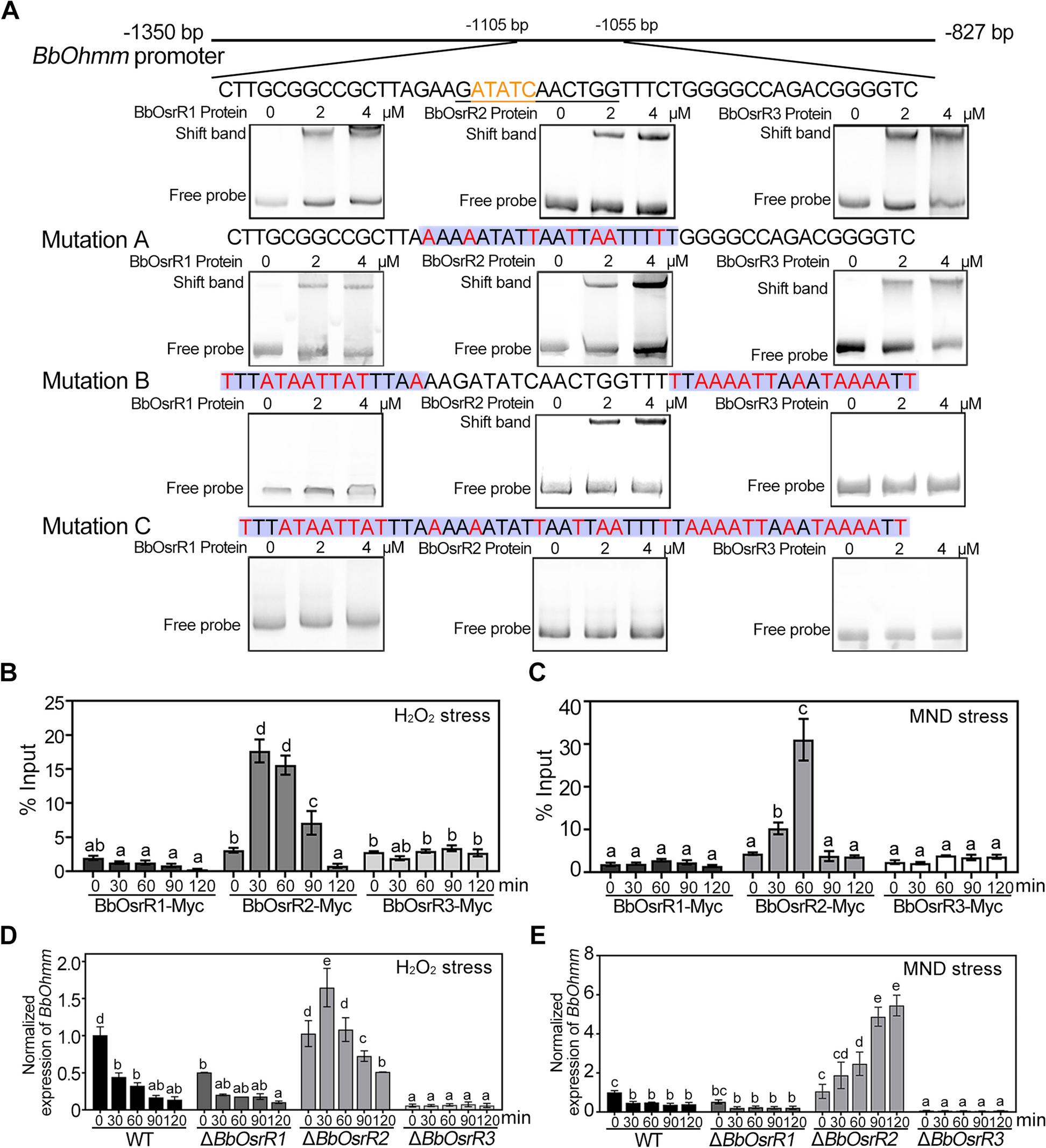
a Sections of a MSC spheroid imaged with a light-sheet microscope. The technique allows in-depth imaging of tissue structures. Scale bar is 50 μm. b 3D network representation of a MSC spheroid. Different cell types are identified based upon CD146 fluorescence measurements. c Comparison between the degree distribution of an example spheroid (panel b, bars) and the batch distribution (N = 5, red line). d Comparison between the link-length distribution of an example spheroid (bars) and the batch distribution (red line). e The network representation makes it possible to identify cells on the outer layer of the spheroid (red) from the inner cells (blue). We can “peel off” the outer layers successively, revealing the inner structure and composition of the spheroid. f Cell degree as a function of the layer number, the average degree is larger for the layers near the center of the spheroid. Blue dots show one example spheroid and red dashed line represents average over the experimental batch. g Distance between cell centers (in μm) as a function of the layer number. Blue dots show one example spheroid and red dashed line represents average over the experimental batch
The MSC spheroids, with CD146 and DNA staining (see the “Methods” section for the full protocol), were imaged in 3D thereby providing access to the cell nuclei and to the distribution of CD146-expressing cells. The images were then segmented using Cellpose [21] and fed into Griottes in order to generate the 3D network representation of the organoid for further analysis. Since only the nuclei were available in these images, the Delaunay construction method was used to estimate the 3D graph, as shown in Fig. 3b. For illustration purposes, the cells here are classified depending on the CD146 expression with the same methodology as in the section for zebrafish, above.
This network representation allows us to retrieve the distribution of number of neighbors (degree) for each cell within the tissue. By repeating the measurement on N = 5 spheroids, the distribution of degrees is found to be well-described by a normal distribution, centered at 〈k〉=11.4±3.2 neighbors per cell (Fig. 3c). Other properties, such as the distribution of distances between every two neighbors within the samples, can readily be obtained as well (Fig. 3d). In the MSC spheroids, cells are on average separated by 〈l〉=23±2.4 μm (Fig. 3d).
It is also possible to assign a layer number to each of the cells in the network and to analyze the cells based on their distance from the edge or center of the organoid. This allows us to virtually peel off the cells composing the outer layers of the spheroid or to treat each layer individually (Fig. 3e). Analysis of the average degree of cells composing each layer, see Fig. 3f, shows an increase from an average of 〈k0〉=8.0±2.4 neighbors per cell in the outer layer (layer 0) to 〈k6〉=13.3±2.9 neighbors in the inner layers (layer 6 in this case). In contrast to the increase in the number of neighbors as a function of the layer, the cell-cell distance remains stable across the spheroid depth. Indeed, the mean distance, defined by length from one nuclei barycenter to another, varies between 〈l3〉=22.6±2.3 μm and 〈l0〉=23.8±2.6 μm (Fig. 3g). In the future, the layer analysis framework could be combined with categorical information on the cells, such as their phenotype, to assess the formation of the spheroids and organoids in greater detail.
Building a network from a data tableImage processing software can often yield cell information in a table format containing the cell positions as well as other attributes such as the cell type. The data table can contain the positions of the cells in the x−y plane, as well as further information about the cells, such as their cell type. Below we show how such data can be integrated and analyzed using Griottes. In the current example a region of interest of the zebrafish telencephalon was segmented and analyzed [20], attributing a cell type to each of the visible cells in the sample. Understanding cellular organization and cell-cell relationships of this tissue will be critical to better understand how this stem cell population coordinates its behavior to sustain large-scale and long term homeostasis.
The cell positions can serve to place them on a two-dimensional scatter plot where the different cell phenotypes are indicated by different colors (Fig. 4a). Griottes is then used to build the network representation of the tissue, here using the Delaunay triangulation to connect cells to each other (Fig. 4b).
Fig. 4
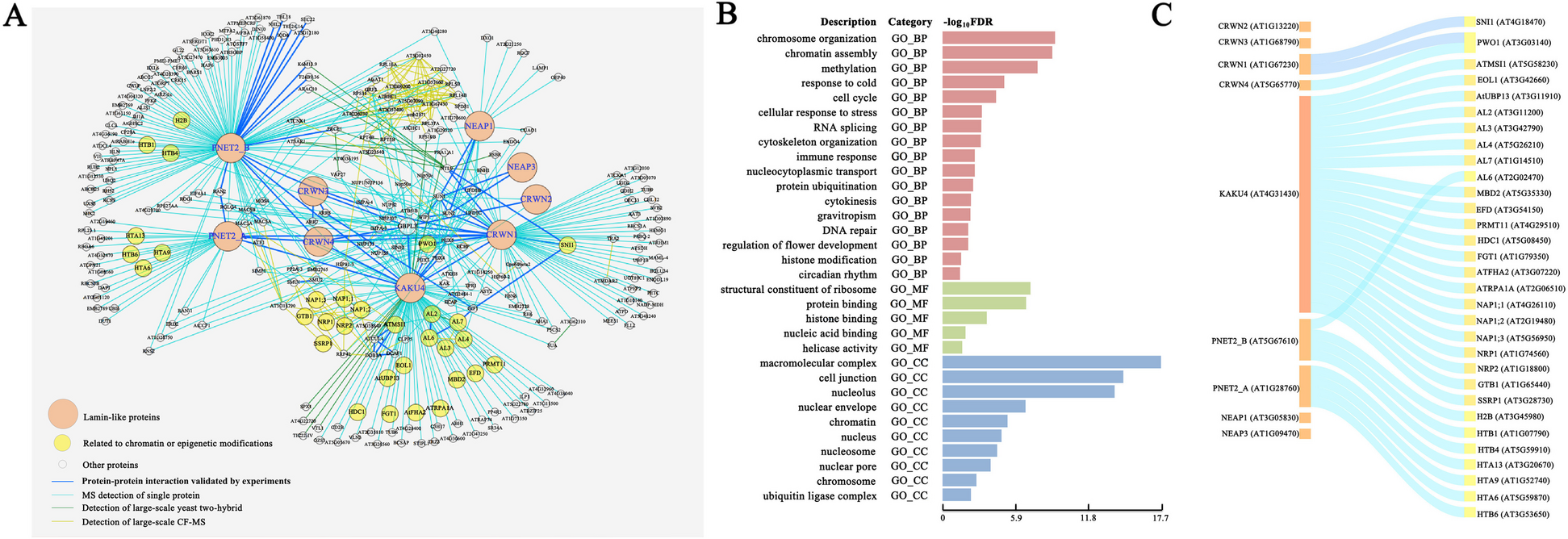
a Reconstruction of the zebrafish telencephalon from a data table. Colors represent the different cell types entered in the table by the user. b Network construction with Griottes from a point-cloud using the Delaunay construction rule. c Degree distribution of cells composing the zebrafish telencephalon after the network construction using the Delaunay rule. d Composition of the zebrafish telencephalon: a vast majority of the cells are GFAP+/PCNA−. e From the network representation of the tissue we can extract clusters of any given cell type (left). Distribution of PCNA+ cluster sizes for clusters of two cells or more (right). f Percentage of cells that belong to a cluster of their cell type of size larger than 2. All GFAP+/PCNA− cells are connected and belong to the same cluster. Conversely, a majority of GFAP+/PCNA− and GFAP+/PCNA+ cells aren’t connected to any other cell of the same type
The telencephalon is structured in a quasi-2D structure, the cells spreading out and forming a sheet. Analysis of the degree distribution of the cells in the tissue (Fig. 4c) shows a similar distribution to that recovered from the cell membranes in Fig. 2c. The tissue contains multiple cell-types necessary for maintaining the tissue homeostasis (Fig. 4d) [20]. A key question to understand the extrusion of cells from the zebrafish telencephalon is the spatial organization of cell phenotypes within the tissue. One method is to study the distribution and properties of clusters of cells of similar phenotype, i.e., a group of cells of the same cell type that are connected to each other. Such clusters can be identified by analyzing the network (Fig. 4e), which makes it possible to compute the proportion of cells of each type that belong to clusters of size larger than 1 (Fig. 4f). We can look at the distribution of cell clusters within the tissue to determine the statistical properties of the organization of the tissue (Fig. 4e).
Looking at the proportion of clusters strictly larger than 1, we see differences depending on the cell types. GFAP+/PCNA− cells, which compose 73% of the tissue, all belong to the same cluster. On the other hand, despite differing presence within the tissue, GFAP+/PCNA+ and GFAP−/PCNA+ cells have a similar chance of belonging to a cluster. Using the network representation makes it possible to investigate experimental data in a new light, whilst highlighting the cell-cell interactions within the samples.
留言 (0)