
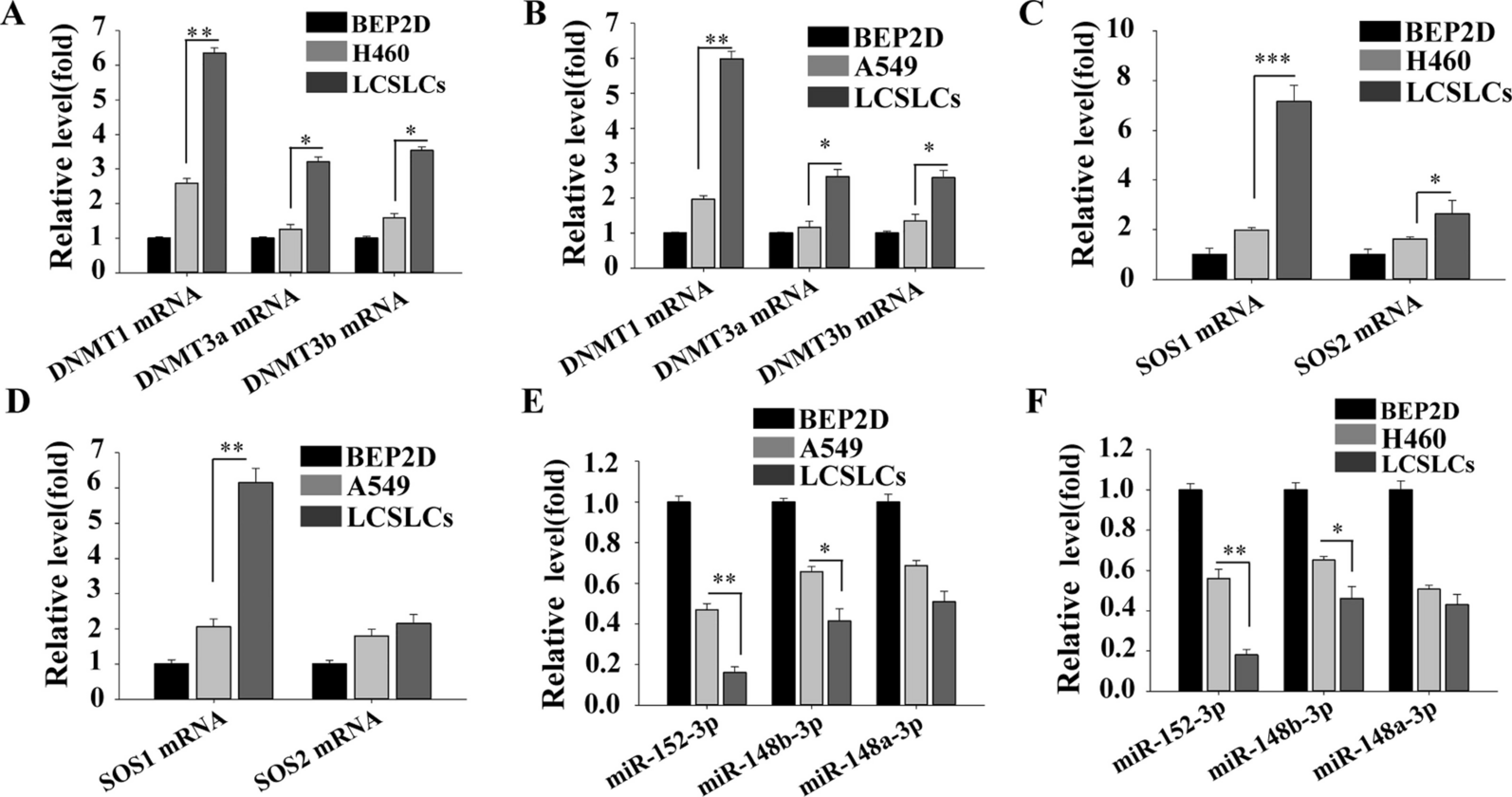
記住我
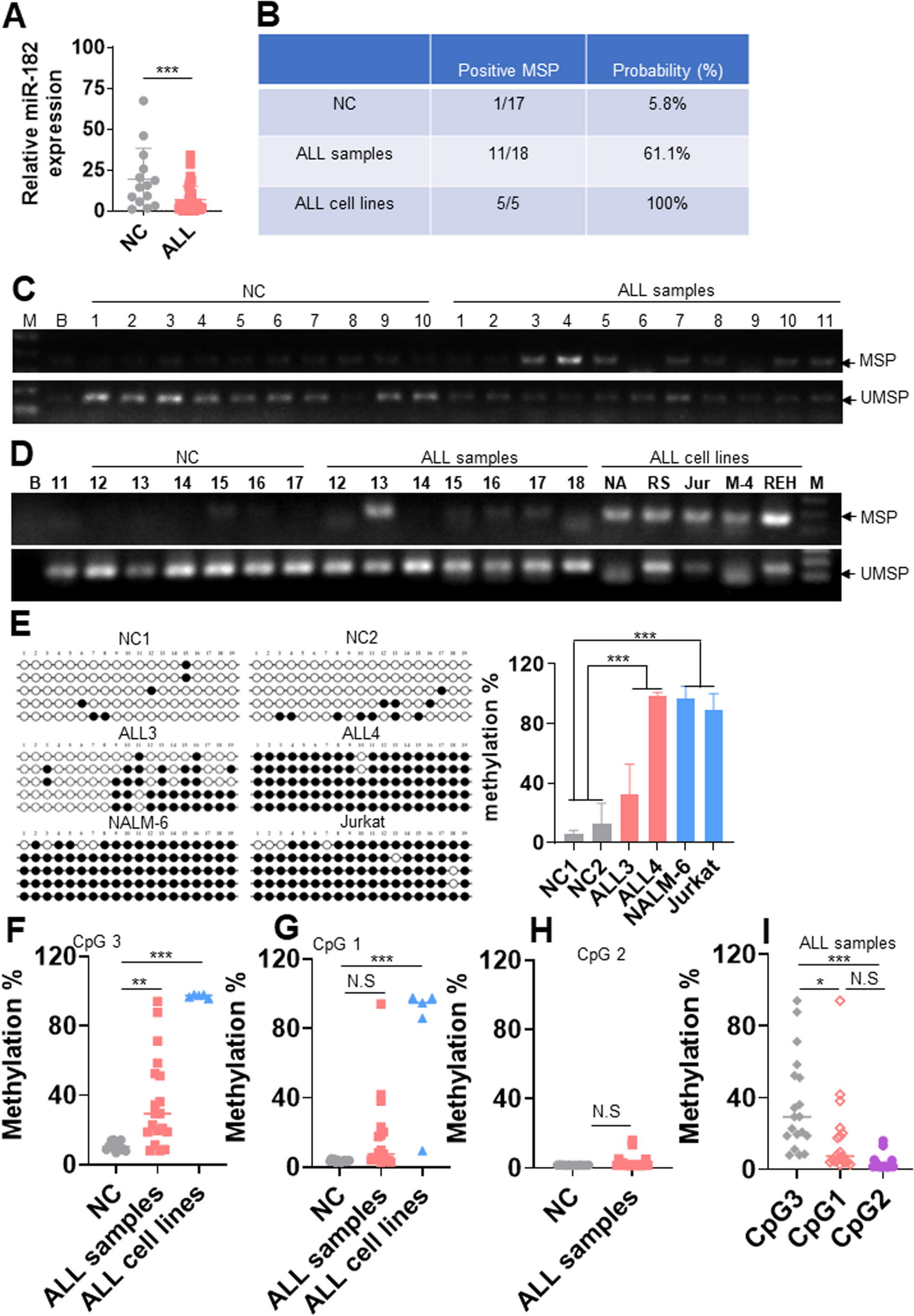
Firstly, we evaluated the differential expression of BCAM in tumor and normal tissues at pan-cancer RNA level from TCGA (The Cancer Genome Atlas) database (Fig. 1A). Compared with the median expression level of corresponding normal tissues, BCAM expression remarkably decreased in all three classic subtypes of RCC (low-expressed proportions: ccRCC: 518/530, 97.74%; pRCC, 285/288, 98.96%; chRCC, 59/65, 90.77%; p < 0.001). The low expression of BCAM in RCC was sequentially validated in the other six external datasets (Fig. 1B). Moreover, its expression profile in ccRCC tumor samples was further verified from GSE53757 (n = 72), GSE40435 (n = 101) and GSE66272 (n = 26) datasets, all with a p < 0.001 (Fig. 1C). The results mentioned above confirmed that BCAM expression at mRNA level was lower than normal tissues in all the three subtypes of RCC (p < 0.05).
Fig. 1
BCAM mRNA and protein expression was lower in RCC tissues than in normal kidney tissues. A BCAM mRNA expression in tumor and normal tissues from pan-cancer data of The Cancer Genome Atlas (TCGA). *p < 0.05, **p < 0.01, ***p < 0.001. B BCAM mRNA expression in tumor and normal tissues from ccRCC, pRCC and chRCC obtained from 6 external datasets, including Higgins Renal, Gumz Renal, Beroukhim Renal, Yusenko Renal, Lenburg Renal and Jones Renal. C BCAM mRNA expression in tumor and normal tissues from ccRCC obtained from Gene Expression Omnibus (GEO) database, including GSE53757, GSE40435 and GSE66272. ****p < 0.0001. D Representative microphotographs of BCAM immunohistochemical staining in normal kidney tissue and ccRCC tissue by IHC. E BCAM mRNA expression in the single-cell-type clusters identified in normal kidney tissues. F BCAM protein expression in tumor and adjacent normal tissues from 6 patients in West China Hospital cohort by immunoblotting. The normalized data after quantitative analysis were shown on the right. **p < 0.01. G BCAM protein expression in tumor and normal tissues from ccRCC data of Clinical Proteomic Tumor Analysis Consortium (CPTAC). ***p < 0.001
We then investigated the expression pattern of BCAM in RCC and adjacent normal tissues at the protein level using paraffin-embedded samples from HPA (The Human Protein Atlas) database. Firstly, immunohistochemical staining showed that for the BCAM protein in 12 ccRCC tumor tissues and 3 normal renal tissues, BCAM protein could be detected in all three normal tissues with moderate intensity and 75–25% quantity in membrane and cytoplasm of renal tubules, but was almost (11 of 12) undetectable in ccRCC tumor tissues (Fig. 1D). Using single-cell RNA sequencing (scRNA-seq) data of human kidney from GSE131685, a total of 25,279 cells from kidney tissues were divided into 14 clusters, including proximal tubular cells (c-0, c-1, c-2, c-3, c-4, c-5, c-6, c-8, c-10), distal tubular cells (c-11), collecting duct cells (c-12), T cells (c-7) and macrophages (c-8), B cells (c-13) [20]. Markers for collecting duct cells were AQP2, CLDN8, PVALB and TMEM213. Markers for distal tubular cells were SLC12A1, TMEM72 and UMOD. Markers for proximal tubular cells were MIOX, SLC22A8 and TMEM174. Other markers are listed in Additional file 1: Table S1. We identified that BCAM mRNA was mainly expressed in epithelial cell clusters (Fig. 1E). The expression level of BCAM mRNA was the highest in collecting duct cells c-12, followed by proximal tubular cells c-8 and c-5, which was consistent with the result that BCAM protein was mainly distributed in renal tubules of normal tissues according to IHC staining. Subsequently, we conducted western blotting with frozen samples in our center. The normalized results were consistent with the previous clues, suggesting the downregulation of BCAM protein in ccRCC (Fig. 1F). Finally, we utilized CPTAC (Clinical Proteomic Tumor Analysis Consortium) database to reconfirm the expression difference of BCAM between tumor and normal tissues in ccRCC at the protein level (Fig. 1G). A total of 110 primary tumor tissue samples were included and 84 normal tissue samples as the negative control. The result showed that 90.91% (100/110) of ccRCC tissues had lower protein expression of BCAM (below the median BCAM expression level of normal kidney tissue), and the difference in BCAM protein level between ccRCC and normal tissues was statistically significant (p < 0.001, fold change = 2). The expression of BCAM at protein level was consistent with those at RNA level, revealing that BCAM expression was indeed downregulated in renal tumor tissues at both mRNA and protein levels.
Low BCAM expression was associated with adverse clinicopathological parameters and poor prognosisWe then thoroughly investigated the potential functional roles of BCAM dysregulation in RCC. Clinicopathological parameters were collected and analyzed, including age, gender, pT stage, pN stage, metastatic status and tumor grading (ISUP grading). Firstly, we explored the relationship between BCAM mRNA expression and clinicopathological characteristics in the whole RCC cohort (Additional file 2: Table S2). The results revealed that BCAM mRNA expression negatively correlated with age, pT stage, metastatic status and tumor grade (all p < 0.05). The association of BCAM expression at the mRNA level with clinicopathological parameters was further analyzed and stratified by RCC histological types. For ccRCC, the expression of BCAM was again inversely correlated with pT stage, metastatic status and tumor grade (Fig. 2A–D). For pRCC and chRCC, there was no relationship between BCAM and clinicopathological variables except for the pN stage (Additional file 3: Fig. S1A–E). Finally, the association of BCAM mRNA expression from the TCGA database with overall survival (OS) was also analyzed, and the result demonstrated that low expression of BCAM in ccRCC was negatively related to OS, while within pRCC and chRCC, BCAM expression did not affect survival status (Fig. 2G).
Fig. 2
Low BCAM expression was associated with several clinicopathological characteristics and poor prognosis in ccRCC. A BCAM mRNA expression was associated with pT stage in ccRCC. B BCAM mRNA expression was not associated with pN stage in ccRCC. C BCAM mRNA expression was associated with metastatic status in ccRCC. D BCAM mRNA expression was associated with tumor grading in ccRCC. E BCAM protein expression was associated with tumor grading in ccRCC. F BCAM protein expression was associated with pan-cancer subtype in ccRCC. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. G Kaplan–Meier analysis of the association between BCAM expression and OS in ccRCC, pRCC, chRCC
After that, we analyzed data from the CPTAC database and found that the relationship between BCAM expression at the protein level and tumor grading was consistent with that at the mRNA level (Fig. 2E). Moreover, according to mass-spectrometry-based proteomic classification (K1–K10), 194 ccRCC enrolled in the database could be subclassified into 8 subgroups (except K4 and K6) [21]. Each subgroup had notable features. For example, K2 and K3 were featured by the activation of immunity signaling pathways. In general, BCAM expression level among different proteomic subtypes was remarkably lower than that in adjacent normal tissues, and it was noted that those who belonged to K2 subtype, which was associated with adaptive immune response and T cell activation, had the lowest expression level of BCAM, indicating the potential association between BCAM downregulation and the activation of immune-related signatures (Fig. 2F).
Potential mechanisms of BCAM dysregulation in ccRCCGenetic analysis indicated that DNA mutation and CNV had no certain effects on BCAM downregulationSince the expression of BCAM was correlated with various clinical characteristics and predicted the prognosis in ccRCC, it might be of clinical significance to divide ccRCC samples into BCAM-low and BCAM-high subgroups. We then set the median expression of BCAM as the cutoff value and further seek for multi-omics differences between the BCAM-high and BCAM-low subgroups. To find out the potential regulatory mechanism of the BCAM gene in ccRCC, we initially detected the DNA alteration and copy number variation (CNV) status, based on the differential expression of BCAM within ccRCC data in the TCGA cohort. The extremely low frequency (0.3%) of somatic mutation with BCAM itself did not explain its high percentage of low expression among ccRCC (Additional file 3: Fig. S1F). As shown in Fig. 3A, a much higher frequency of BAP1 mutation was the only alteration related to BCAM low expression, predicting the poor prognosis (17.24% vs 4.47%, p < 0.05). In BCAM-low subgroup, patients with BAP1 mutation had a worse OS than those without BAP1 mutation (Additional file 3: Fig. S1G).
Fig. 3
BCAM expression was associated with gene mutation features and DNA methylation modification in ccRCC. A Alteration frequency of the genes with the highest alteration frequency in the whole ccRCC cohort between the BCAM-low and BCAM-high subgroups, and the correlations with these genes and the OS prognosis of ccRCC patients. B The association between BCAM expression and tumor mutational burden (TMB) and microsatellite variation (MSI). C The association between BCAM expression and copy number variation (CNV). D The correlation of BCAM expression and the expression of DNA methylation-related genes
We also analyzed the correlation between BCAM gene expression and tumor mutation burden (TMB)/microsatellite instability (MSI). In ccRCC, lower expression of BCAM was correlated with higher TMB (Fig. 3B). At the same time, there was no relation between BCAM and MSI status. In the end, we investigated the association of CNV with BCAM expression. We found that there was no correlation (r = 0.042) (Fig. 3C). Taken together, these results suggested that genetic alteration could not explain the dysregulation of BCAM; instead, low BCAM expression could be associated with a higher frequency of BAP1 mutation in ccRCC.
Hypermethylation was closely related to BCAM downregulationNext, we explored whether the downregulation of BCAM in ccRCC might be regulated by epigenetic modification using the TCGA database. We initially took the DNA methylation modification into account and exhibited the correlation between BCAM gene and several genes involved in DNA methylation (Fig. 3D). As the results illustrated, BCAM gene positively correlated with the expression of most methylation related enzymes in ccRCC, such as MECP2 (r = 0.399, p < 0.001), MBD3 (r = 0.28, p < 0.001) and TET2 (r = 0.244, p < 0.001). We then generated a waterfall plot to associate methylation levels with gene subregions visually based on the sequencing results of 450 k DNA methylation chips in the TCGA project (Fig. 4A). Several BCAM-related probes had a high extent of methylation, including cg03074126, cg14037553, cg17489534, cg24122751, cg22640961, cg12249345, cg21978694, cg05670193 and cg23318764. We further thoroughly investigated the specific correlations between the methylation level of these probes and BCAM expression in ccRCC. The results revealed that, in the 333 samples of ccRCC, the methylation level of 8 out of 9 probes negatively correlated with BCAM gene expression, in which CpG island-related probe cg22640961 (r = − 0.54, p < 0.001) showed the strongest negative correlation, followed by cg12249345 (r = − 0.52, p < 0.001), cg21978694 (r = − 0.51, p < 0.01), cg24122751 (r = − 0.47, p < 0.001), cg14037553 (r = − 0.33, p < 0.001), cg17489534 (r = − 0.32, p < 0.001), cg23318764 (r = − 0.24, p < 0.001) and cg05670193 (r = − 0.17, p < 0.01). However, cg03074126 (r = 0.53, p < 0.001) was the exception and its methylation level strongly positively correlated with BCAM expression. Also, when integrating the results of CpG island-related probes, the aggregation result showed a correlation coefficient of − 0.54 (p < 0.001), indicating that the low expression of BCAM could be attributed to its CpG island-related methylation modification. Furthermore, we studied whether the methylation degree of these probes differed between tumor and normal tissues. We found that all the methylation level of probes, except for cg03074126 and cg05670193, was significantly higher in tumor tissues than that in normal tissues in ccRCC (Fig. 4B). Given the above-analyzed results that low BCAM expression coexisted with a higher frequency of BAP1 mutation in ccRCC, we further tried to explore the relationship between BAP1 mutation status and BCAM methylation level. To our surprise, three probes of BCAM, including cg06522456 (p < 0.01), cg08319238 (p < 0.001) and cg14037553 (p < 0.01), showed higher level of methylation in ccRCC with BAP1 mutation compared to BAP1 wild-type ccRCC, indicating the potential mechanism of BAP1 mutation regulating BCAM expression level (Additional file 3: Fig. S1H).
Fig. 4
BCAM expression was associated with DNA methylation status in ccRCC, and the BCAM-low and BCAM-high subgroups had different enriched functions and pathways. A The association of methylation level with gene subregions. B The DNA methylation level of different probes between normal and tumor tissues. C Differential genes between the BCAM-low and BCAM-high subgroups and 50 upregulated genes and 50 downregulated genes with the largest differential changes. D KEGG pathway enrichment analysis and GO enrichment analysis of genes upregulated in the BCAM-low subgroup and genes upregulated in the BCAM-high subgroup
In general, these results indicated that BCAM-related epigenetic modification was obviously associated with its gene regulation and RCC patient prognosis.
Different expressions of BCAM indicated different transcriptomic characteristicsWe attempted to further explore the transcriptomic differences between BCAM-low and BCAM-high groups and find clues for potential treatment schemes, respectively. Therefore, we first observed the differentially expressed genes between the BCAM-low and BCAM-high subgroups based on the TCGA database (Fig. 4C). The results demonstrated that, compared to BCAM-high subgroup, there were 145 up-expressed genes and 539 down-expressed genes in BCAM-low subgroup (Additional file 4: Table S3).
Low BCAM expression was associated with high immunogenicityBoth the KEGG pathway (Up) enrichment analysis and the GO (Up) enrichment analysis demonstrated that the up-expressed genes among the BCAM-low subgroup were mostly enriched in the immune-related signaling pathway, including “cytokine–cytokine receptor interaction,” “chemokine signaling pathway,” “cell adhesion molecules” and “antigen processing and presentation” (Fig. 4D). To further investigate the immune status in ccRCC, we compared the immune infiltration between BCAM-low and BCAM-high subgroups. After mining data from TCGA cohort, we found that in ccRCC samples, most immune cell types were increased in BCAM-low subgroup, such as M2 macrophage (p < 0.001), monocyte (p < 0.001), myeloid dendritic cell (p < 0.001), CD8 + T cells (p < 0.001) and CD4 + Th2 cells (p < 0.001) (Fig. 5A). The composition of these immune cell subpopulations in tumor microenvironment (TME) of ccRCC was also calculated (Fig. 5B) [22]. Data from CPTAC database further demonstrated that, when compared with metabolic immune desert, VEGF immune desert and CD8− inflamed subgroups, the CD8 + inflamed subgroup had a lowest BCAM mRNA (the proportion of Z-Score > 0 = 18.5%) and protein (the proportion of Z-Score > 0 = 16.0%) expression level, and a highest BCAM methylation level (the proportion of Z-Score > 0 = 55.6%), consistent with the above results (Fig. 5D). Additionally, the VEGF immune desert subgroup had the highest BCAM mRNA (the proportion of Z-Score > 0 = 44.0%) and protein (the proportion of Z-Score > 0 = 68.0%) expression level and the lowest BCAM methylation level (the proportion of Z-Score > 0 = 4.2%) among the 4 immune subgroups. We also evaluated the expression of 8 immune checkpoints of BCAM-low and BCAM-high subgroups using the TCGA database to see whether there were differences in expression level in ccRCC (Fig. 5C). The results revealed that, compared to BCAM-high subgroup, the expression of CD274 (p < 0.001), CTLA4 (p < 0.001), HAVCR2 (p < 0.001), LAG3 (p < 0.001), PDCD1 (p < 0.001), PDCD1LG2 (p < 0.001) and TIGIT (p < 0.001) was elevated by varying degrees in BCAM-low subgroup. Further analysis revealed that these 7 immune checkpoints all had lower methylation levels in BCAM-low subgroup compared to the BCAM-high subgroup (p < 0.01) (Additional file 5: Fig. S2A). The results demonstrated that immune checkpoints might lead to immune escape of tumor cells in ccRCC.
Fig. 5
Low BCAM expression was associated with high immunogenicity; distinct BCAM expression patterns could indicate potential therapeutic strategies in ccRCC. A Immune cell score in the BCAM-low and BCAM-high subgroups. *p < 0.05, **p < 0.01, ***p < 0.001. B The percentage abundance of tumor-infiltrating immune cells in each sample. C The expression of immune checkpoints in the BCAM-low and BCAM-high subgroups. ***p < 0.001. D The association between different immune subgroups and the mRNA and protein expression and methylation level of BCAM, and the proportion of Z-Score > 0 of the 4 immune subgroups in BCAM methylation level, mRNA expression level and protein expression level, respectively. E The expression of angiogenesis-related genes in the BCAM-low and BCAM-high subgroups. *p < 0.05, **p < 0.01, ***p < 0.001. F Tumor Immune Dysfunction and Exclusion (TIDE) score of the BCAM-low and BCAM-high subgroups. G Distribution of Sorafenib, Pazopanib and Axitinib IC50 scores in the BCAM-low and BCAM-high subgroups. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
High BCAM expression was associated with the enrichment of angiogenesisOn the other hand, the KEGG and GO enrichment analysis for the BCAM-high subgroup showed that the angiogenesis-related pathways were remarkably enriched (Fig. 4D). We then attempted to further confirm whether the extent of angiogenesis activation was higher in the BCAM-high subgroup of ccRCC sample. The expression level of 36 related genes between BCAM-low subgroup and BCAM-high subgroup was compared, and the results illustrated that 16 angiogenesis related genes, including JAG2, JAG1, MSX1, SLCO2A1, APP, PDGFA, PTK2, VEGFA, NRP1, STC1, FGFR1, LPL, KCNJ8, LRPAP1, CCND2 and THBD, were significantly upregulated in BCAM-high subgroup (p < 0.001) (Fig. 5E), and only four were upregulated among BCAM-low subgroup. We also found that for JAG1, MSX1, SLCO2A1, APP, PTK2, NRP1, STC1, FGFR1, KCNJ8, LRPAP1, CCND2 and THBD, the methylation degree in BCAM-high subgroup was lower than that in BCAM-low subgroup (p < 0.05) (Additional file 5: Fig. S2B). All the findings revealed that more angiogenesis related genes upregulation was enriched in BCAM-high ccRCC patients.
These transcriptomic results suggested that immune-related signaling pathways were overactivated in the BCAM-low subgroup. In contrast, more angiogenesis-related signaling pathways were activated in the BCAM-high subgroup, which implied the different therapeutic strategies in practice.
Potential therapeutic strategies based on distinct BCAM expression patterns in ccRCCFinally, we looked at the predictive value of BCAM in the treatment of ccRCC. The sensitivity of tyrosine kinase inhibitor (TKI) drugs was calculated based on the GSDC database, comparing within differential BCAM expression subgroups. The results showed that the half-maximal inhibitory concentration (IC50) of Sorafenib (p < 0.0001), Pazopanib (p < 0.0001) and Axitinib (p < 0.01) among the BCAM-high subgroup was significantly lower than those within BCAM-low group (Fig. 5F). These results indicated that anti-angiogenesis therapy should be considered for ccRCC patients with BCAM-high expression.
Tumor Immune Dysfunction and Exclusion (TIDE) algorithm modeled immune evasion in tumors by combining both T cell dysfunction and exclusion signatures. The higher the TIDE prediction score was, the worse the immune checkpoint inhibitor (ICI) response was. Using the TCGA database, we calculated and compared the TIDE scores of the BCAM-low and BCAM-high subgroups. In contrast with the BCAM-high subgroup, the TIDE score of the BCAM-low subgroup was much lower (p = 0.00052) (Fig. 5G), indicating that the BCAM-low subgroup might respond better to ICI-based immunotherapy. Considering the upregulation of multiple different immune checkpoints, including CD274, CTLA4, HAVCR2, LAG3, PDCD1, PDCD1LG2 and TIGIT, double or triple ICIs combination therapy might be promising among ccRCC with BCAM-low expression.
留言 (0)