
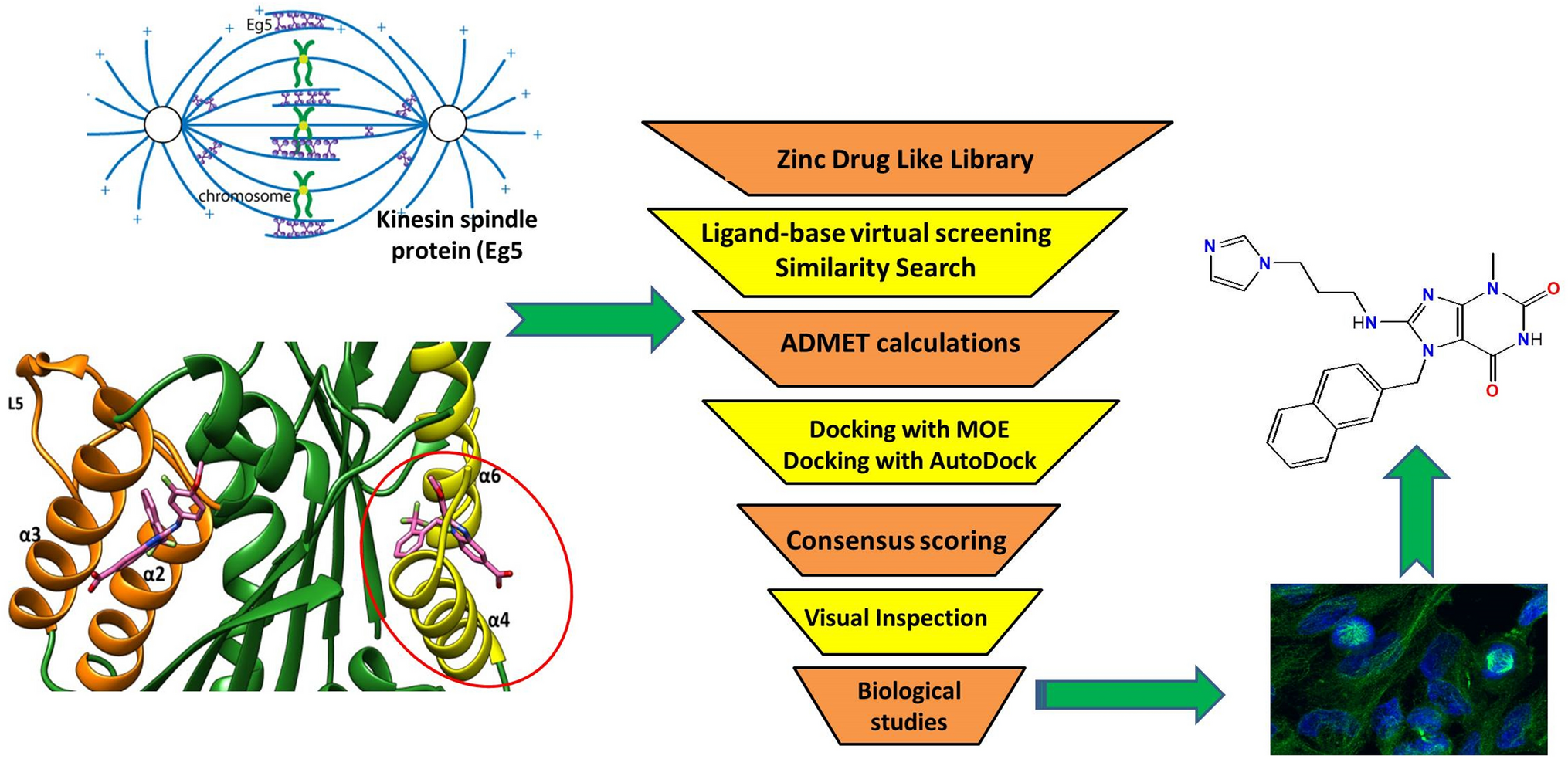
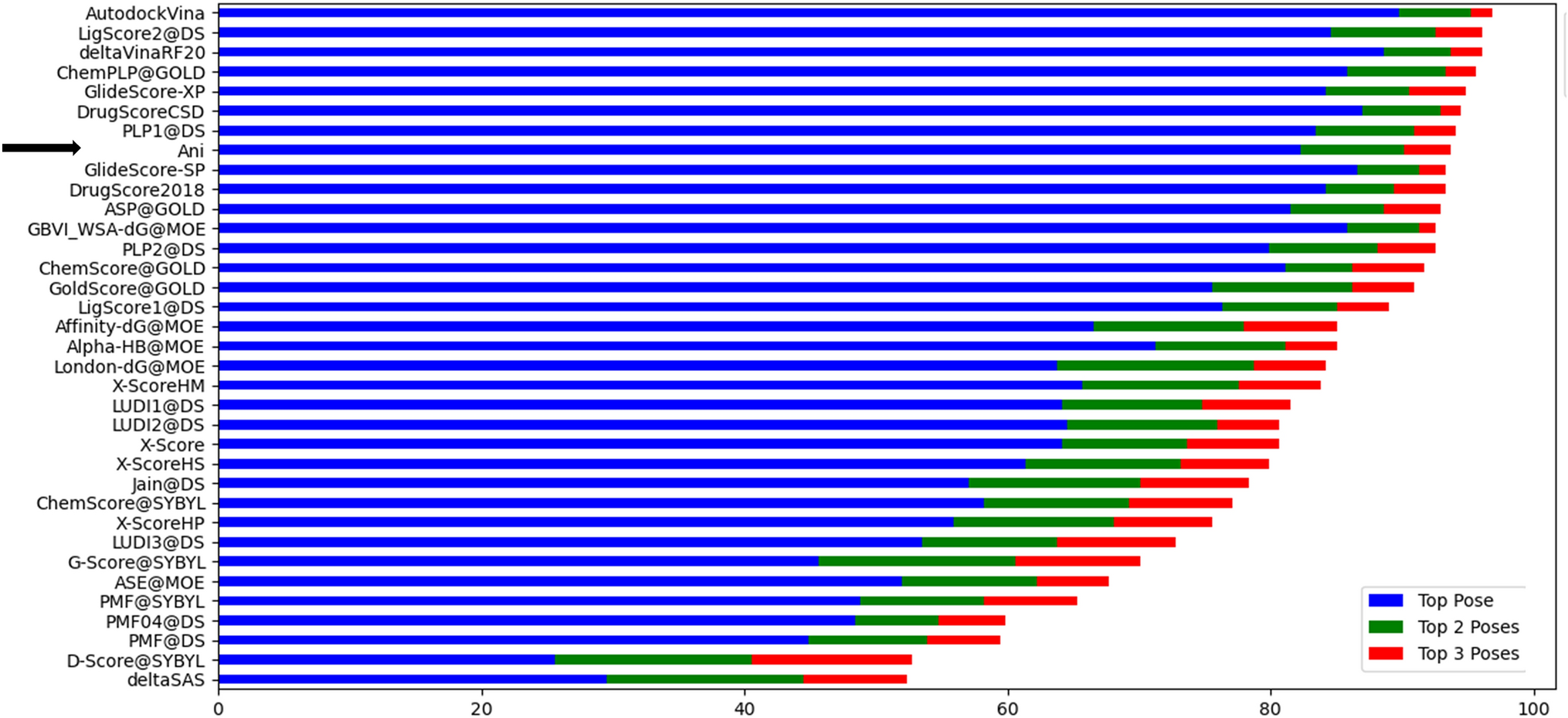
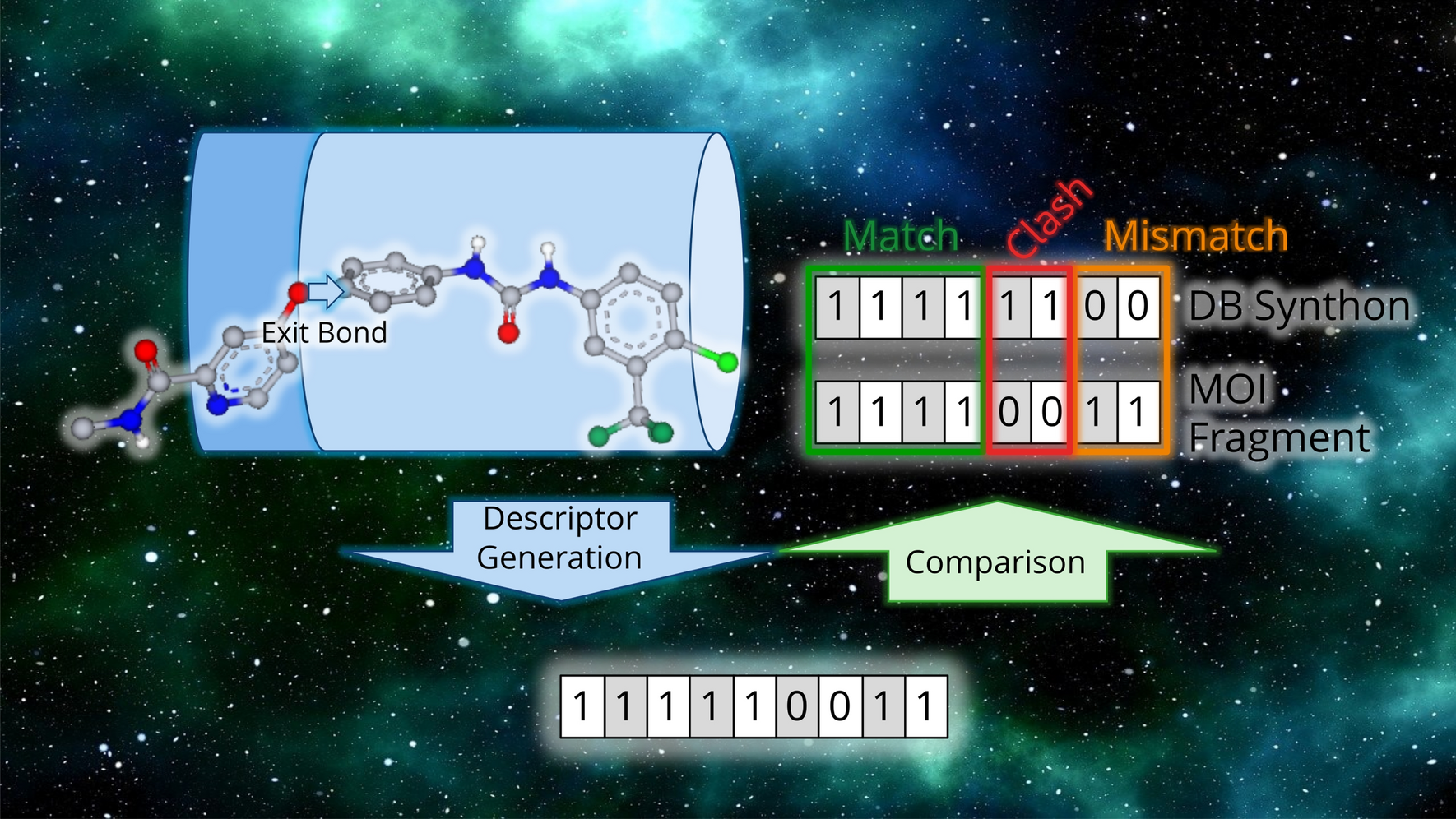
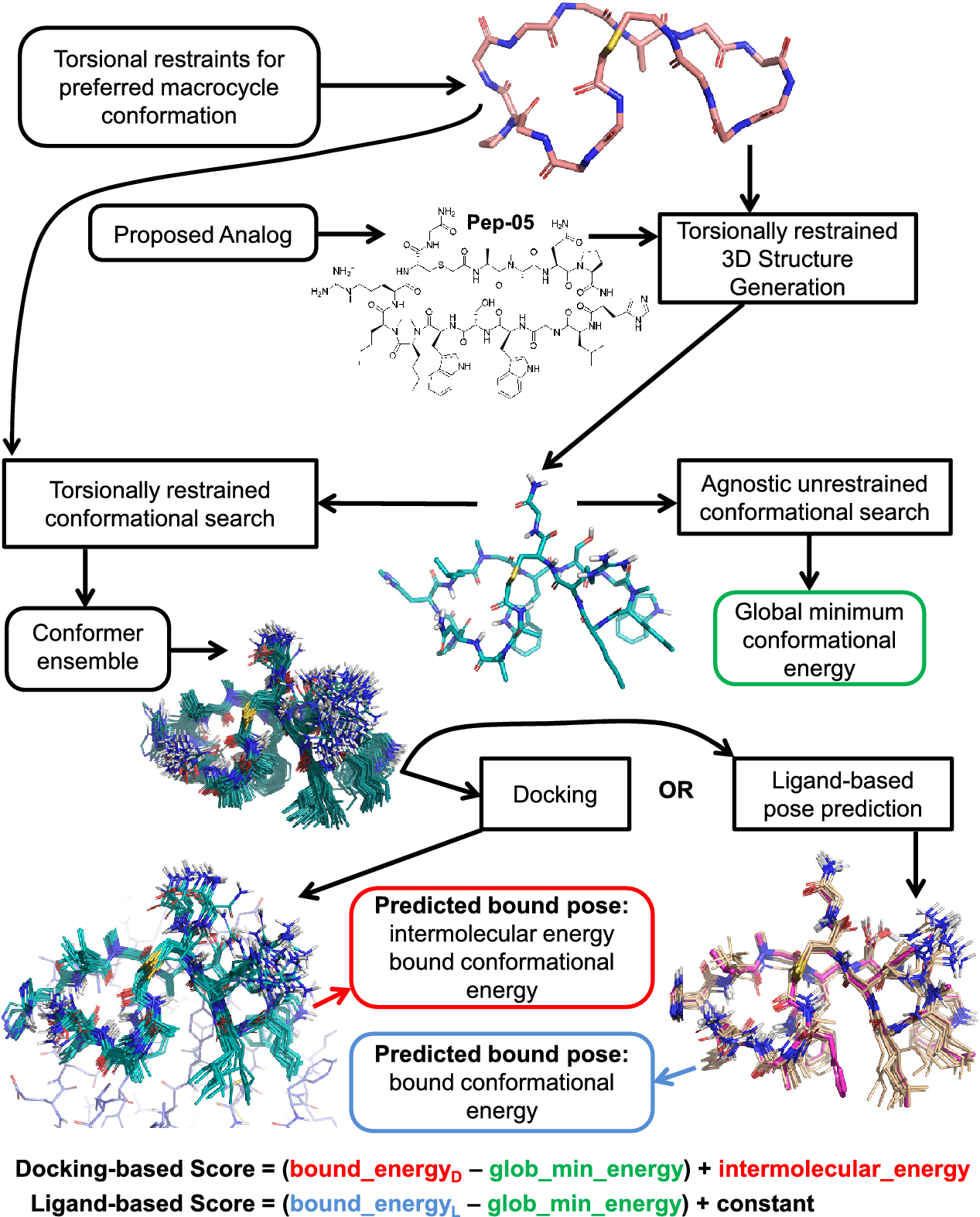
記住我

The Root-mean-square deviation (RMSD) of atomic positions is a standard measure of the structural changes mainly used for proteins or non-protein small organic/inorganic molecules. Nevertheless, due to the large conformational flexibility of dendrimers in aqueous solution, there is a debate as to whether it is an appropriate molecular descriptor to account for its structural stability. Here, we have employed RMSD analysis to find the most converged part of the simulations, in order to obtain reasonable initial structures for the following steps. As shown in Fig 2a, the PNH\(^n\) RMSD values undergo large changes from the starting structure in the first 50 ns, afterwards the system seems to reach a more stable behavior; meanwhile PNH\(^a\) RMSD values undergo big changes from the starting structure in the first 20 ns, afterwards the system seems to reach a more stable behavior fluctuating around a mean value. The POH\(^n\) RMSD values undergo a rapid change during the first ns, after that, the RMSD steadily deviates slowly from this new conformation, reaching a more steady-state after 100 ns; meanwhile, POH\(^a\) RMSD values undergo big changes from the starting structure in the first 20 ns, afterwards the system seems to reach a more stable behavior fluctuating around a mean value. Ang-(1-7)\(^n\) and Ang-(1-7)\(^a\) RMSD values show a more stable behavior from the first ns. According to these results, we agreed to use the last 100 ns of the simulations as the most equilibrated data for all the systems. Large changes observed on dendrimer structures are mainly due to the fact that the initial structures were obtained by PyPolyBuilder algorithm which only gives us an initial guess for the dendrimer geometry in vacuum.
Fig. 2
Structural features of PAMAM-NH\(_2\), PAMAM-OH and Ang-(1-7) systems. a RMSD and b R\(_\) values as function of time
Table 3 Radius of gyration R\(_\) of PAMAM dendrimers and comparison with available experimental and theoretical studies in the literatureThe radius of gyration (R\(_\)) might be a more appropriate molecular descriptor to account for the structural stability of dendrimers, it gives a measure for the compactness of a molecular structure. As shown in Fig. 2b, the PNH\(^n\) and PNH\(^a\) R\(_\) values show a more stable behavior from the first nanoseconds around a mean value. POH\(^n\) R\(_\) undergoes a rapid change during the first nanoseconds, afterwards it steadily deviates slowly from this new conformation, reaching a more steady-state after 100 ns. POH\(^a\) R\(_\) values show that the dendrimer undergoes a rapid change during the first nanoseconds and afterwards fluctuates around a mean value; finally, Ang-(1-7)\(^n\) and Ang-(1-7)\(^a\) R\(_\) values show a more stable behavior from the first nanoseconds around a mean value in both cases.
PNH\(^n\) and PNH\(^a\) R\(_\) mean values in the most equilibrated zone were 1.94 ± 0.05 nm and 2.3 ± 0.04 nm, respectively. On the other hand, POH\(^n\) and PNOH\(^a\) R\(_\) mean values were 1.46 ± 0.02 nm and 2.16 ± 0.06 nm, respectively. Results show that dendrimers at acidic pH assume a stretched, open configuration similar to the dense-shell model as a consequence of the strong Coulomb repulsion between the charged tertiary/primary amines present in the structure. R\(_\) values are in very well agreement with experimental and theoretical results available [31, 46,47,48,49,50,51], as also compared in Table 3, confirming that GROMOS-compatible 2016H66 force field, together with our chosen methodology were capable to represent the structural properties of the dendrimers used in this work. It can be observed that POH\(^n\) and POH\(^a\) has in general a more compact structure (lower R\(_\)) compared with PNH\(^n\) and PNH\(^a\), respectively (Fig. 2b). At neutral pH, this is partly due to the preference of the group terminals to be surrounded by waters (PNH\(^n\), Fig. 3a) or to interact with internal dendrimer groups (POH\(^n\), Fig. 3b), from the total possible interactions between group terminals and waters/internal dendrimer groups, we found that PNH\(^n\) forms 95 \(\%\) of the interactions with waters and 5 \(\%\) with internal groups, meanwhile, POH\(^n\) forms 85 \(\%\) of interactions with waters and 15 \(\%\) with the internal groups. The R\(_\) and RMSD fluctuations in the case of PNH\(^n\), PNH\(^a\) and POH\(^a\) throughout the simulation time, compared to that of POH\(^n\), suggests that the formers have a more flexible structure than the latter. This result might be attributed to the continuous movement of the protonated terminal amine groups in PNH\(^n\) and terminal/internal protonated groups in PNH\(^a\) and POH\(^a\) to minimize the unfavorable enthalpic penalty due to electrostatic repulsion on charges of the same sign.
Fig. 3
Waters sorrounding group terminals and hydrogen bonds formed between terminal groups and waters/dendrimer internal groups. a PAMAM-NH\(_2\) and b PAMAM-OH, at neutral pH
Fig. 4
Dendrimers shape comparision. a PAMAM-NH\(_2^n\), b PAMAM-OH\(^n\), c PAMAM-NH\(_2^a\) and d PAMAM-OH\(^a\) at 200 ns of MD simulation. Leters “a” and “n” states for neutral and acidic pH
Fig. 5
Dendrimers cavities comparision. a PAMAM-NH\(_2^n\), b PAMAM-OH\(^n\), c PAMAM-NH\(_2^a\) and d PAMAM-OH\(^a\) at 200 ns. Leters “a” and “n” states for neutral and acidic pH
To characterize the shape of the dendrimers, an asphericity parameter \(\delta\) was estimated. It is important to note that the closer this value is to zero, the more spherical the dendrimer becomes and its calculation is described in detail elsewhere [31]. As shown in Table 4 and Fig. 4, PNH\(^n\) assume a less spherical structure compared to POH\(^n\), meanwhile, PNH\(^a\) and POH\(^a\) show an almost spherical behavior. This high asphericity observed in PNH\(^n\), might be due to an asymmetry induced by a high degree of charged terminal groups back folding as also reported in the previous works [31], in contrast, the neutral terminal in POH\(^n\) seems to be able to accommodate in a more compact way due to its less charge-charge terminal repulsion and its preference of interactions with internal dendrimer groups. In the case of PNH\(^a\) and POH\(^a\), an almost spherical shape might be mainly due to the repulsion between charged tertiary amine groups in the interior of the dendrimers.
Table 4 Asphericity \(\delta\) of PAMAM dendrimer and comparison with theoretical studies available. Leters “a” and “n” states for neutral and acidic pHTo characterize how particle density (waters, terminal nitrogens, internal nitrogens and ions) varies as a function of the distance from the dendrimer center, radial distribution functions were measured. As shown in Fig. 6a, a higher level of structuration is evidenced by the pronounced peaks on the distributions on PNH\(^n\), PNH\(^a\) and POH\(^a\) internal nitrogens compared with those of POH\(^n\), this is mainly due to a more dense POH\(^n\) dendrimer compared with less dense, PNH\(^n\), being more evident on PNH\(^a\) and POH\(^a\), as evidenced by the height of the peaks. As it can be seen, the dense-core model [53] is confirmed at neutral pH, this model states that the monomer (internal Nitrogen atoms) density distribution is high close to their centers and decays as it approaches the periphery, as evidenced of the height of the peaks from the center to the periphery at neutral pH. However, at acidic pH, dendrimers assume a less dense inner cavity, stretched, open conformation like to the dense-shell model [53] as a consequence of the strong Coulomb repulsions between the charged units. This is in agreement with theoretical models that predicts that the PAMAM-NH\(_\) core becomes denser upon decreasing the acidity of the medium. Interestingly, this could led to a different drug interaction behaviour and load capacity, probably allowing better drug encapsulation in the big inner cavities at acidic pH (Fig. 5).
As shown in Fig 6b, PNH\(^n\) N-ter can be found in the intermediate zone (around 1 nm) as well as in the external surface (around 2 nm), compared with the case of PNH\(^a\) where distribution is near the external surface (around 2.3 nm), and POH\(^n\) and POH\(^a\) O-ter which are mainly found on the surface (around 1.5 nm and 2.16 nm). In PNH\(^n\), terminal groups exhibit a broader distribution that might indicate a certain degree of backfolding, being the backfolding increased from low to neutral pH, as shown by the intensity of the density peaks.
Fig. 6
Radial distribution function of molecular groups with respect of the dendrimer COM. a Internal nitrogen atoms and b terminal nitrogen/oxygen atoms. Schematic diagram at the right of the panel showing the internal and terminal nitrogens. GX states for the dendrimer generation. *Thinner grey line states for comparison from previous work in the literature [31]. Grey vertical dashed line represents the Rg of the dendrimers
As shown in Fig 7a, RDF profiles reveal a major presence of buried water molecules within the structure of PNH\(^n\), PNH\(^a\) and POH\(^a\) compared with POH\(^n\), at distances larger than 5 Å from the center of mass of the dendrimer. Hydroxyl terminals when internal tertiary amines are not protonated reduce the penetration of water into the dendrimer structure, which can be interpreted as an enhancement of the hydrophobic character of the dendrimer inner small cavities, ideal for encapsulation of small hydrophobic drugs. In general, dendrimers with both kind of terminal groups at acidic pH show a major presence of buried waters whithin its structure in order to solvate the charged groups. As shown in Fig 7b-c, RDF profiles reveal that chloride ions penetrate the innermost dendrimer cavities in PNH\(^n\), PNH\(^a\) and POH\(^a\), contrary to the case of POH\(^n\). Interestingly, a broad peak around 1 nm and a shoulder around 2 nm, where PNH\(^n\) positive terminals are localized, was found. Sodium counterions are found as far as posible of charged terminals in PNH\(^n\), PNH\(^a\) and POH\(^a\) and more disperse in POH\(^n\) around its surface.
Fig. 7
Radial distribution function of molecules with respect of the dendrimer COM. a Water molecules and b Cl\(^-\) ions and c Na\(^+\) ions. *Thinner grey line states for comparison from previous work in the literature [31]. Grey vertical dashed line represents the Rg of the dendrimers
Ang-(1-7) bioactive peptideAng-(1-7)\(^n\) and Ang-(1-7)\(^a\), R\(_\) mean values in the equilibrated zone were 0.65 ± 0.04 nm and 0.57 ± 0.05 nm, respectively (Fig. 2b). From the structural point of view , the solution structure of Ang-(1-7) was previously determined [54] by means of NMR and circular dichroism experiments performed at acidic pH. There, the authors claimed that Ang-(1-7) structure showed a conformational equilibrium between the random coil and \(\beta\)-sheet (or a mixture of bend structures), with a bend stabilized by interactions between residues VAL3 and TYR4, this is in very well agreement with our results in Fig. 8, where we can see that the most frequent secondary motif is bent between VAL3 and TYR4. In our classical MD simulations at acidic pH, we found that 60 \(\%\) of the structures were similar to the NMR solution structure (RMSD < 2 Å when superimposed), meanwhile, at neutral pH, this happened only in 40 \(\%\) of the frames, thus, this conformation was less frequent at neutral pH (Fig. 9).
Fig. 8
Ang-(1-7) secondary structure as a function of time. a At neutral pH and b at acidic pH
Fig. 9
Superimposition of: experimental NMR structure obtained at acidic pH (grey), structure repeated 40 \(\%\) of the frames in classical MD simulation performed under neutral pH (blue), structure repeated 35 \(\%\) of the frames in metadynamics simulation performed under neutral pH (cyan), structure repeated 60 \(\%\) of the frames in classical MD simulation performed under acidic pH (yellow)
The conformational sampling of peptides in solution that are fully or partially disordered and flexible is not always guaranteed with conventional MD due to the limiltations in the sampling time scale [55]. To get more insights about Ang-(1-7) heptapetide structure and extend this insight at neutral pH conditions, we performed a MTD simulation at neutral pH employing the R\(_\) and the C\(_\) as CVs. The final free energy landscape is shown in Fig. 10a, where it can be shown that the landscape has a broad and unique minimum or basin A, indicating that this peptide exist in a variety of conformations in solution. The global minima corresponds to a R\(_\) of 0.65 nm and a C\(_\) of 30, in very well agreement with R\(_g\) values from classical MD; however, the basin A samples a wide range of R\(_\) and C\(_\) values, this is a R\(_\) in the region spanning from 0.6 to 0.7 nm and contacts ranging between 30 and 35. Interestingly, the structures in the basin showed a persistent HB between ASP1 and TYR4 or between ASP1 and VAL3 (Fig. 10b). This HB was also frequently observed on the bigger cluster of the MD of Ang-(1-7)\(^n\) in solution, this conformation is probably frequent because it allows positive charged ARG2 to be far from positive charged N-terminal, both of them becoming able to interact with polar waters.
Fig. 10
Ang(1-7) 2D a Free Energy Surface (FES) as a function of the total number of donor-acceptor contacts and the R\(_\). Isoenergy lines are drawn every 5 kcal/mol. T = 300 K, pH 7. b Structures representative of the main free-energy basin A are superimposed in the inset Fig.. c Estimate of the free energy as a function of the R\(_\) from a well-tempered metadynamics simulation
The one dimensional FES as a funcion of R\(_\) is shown in Fig. 10c, as can be seen, there are not large free energy barriers (around 1 kT) dividing the metastable states in the main basin A. It is important to note that only 35 \(\%\) of the structures in basin A were similar to the NMR structure and in agreement with classical MD. This result is coherent with the conformational equilibrium between the random coil and a mixture of bend structures reported experimentally for Ang-(1-7). However, basin A is characterized by a significant degree of flexibility of the N-terminal and C-terminal tails which samples multiple conformations that deviate from the ensemble of the NMR structure as shown in Fig. 11.
Fig. 11
RMSF of Basin A main clusters as a function of aminoacid backbone atoms. Structures representative of the basin A main clusters are superimposed in the right Fig.. Grey represent the NMR structure. Threshold of 0.15 nm is marked to visualize the most flexible atoms. Cluster 1 is marked also with impulses to guide visualization
Together, the R\(_g\), \(\delta\) and RDFs results validates our chosen methodology and confirms that GROMOS-compatible 2016H66 force field is capable to model the theoretical and experimental structural features of G4-PAMAM dendrimers with amino or hydroxyl group terminals. In case of Ang-(1-7), structural super impositions of MD and metadynamics structures with NMR structure confirms the experimental results previously reported.
Clusters and dockingThe middle structure of the most populated clusters for dendrimers and the peptide were used for the molecular docking of each dendrimer conformer with each of the peptide conformers. Our methodology allowed us to find a suitable binding site for the peptide by using a representative peptide conformation taken from the MD, once the peptide finds a reasonable site, the algorithm allows it to better accommodate into this local site, as can be schematized in Fig. 12.
Fig. 12
Double-docking schematic representation. Each of the peptides conformer is allowed to find a site by using a blind rigid docking, afterwards, once the peptide found a reasonable site, a flexible docking allows it to better accommodate into the site
Final complexes resulted from the docking are shown in Fig. 13. As can be seen, PNH\(^n\) allows the interaction of the peptides with its inner shells as well as with its surface. POH\(^n\), only allow the peptide interactions with its surface due to its more compact structure. We can observe that at acidic pH, dendrimers allow a better encapsulation of 3 peptides. Details on the HBs formed between peptides and dendrimers are presented in Tables 7 and 5.
Fig. 13
MD complexes initial structures obtained from double-docking approach: a PNH\(^n\) in complex with 3 Ang-(1-7)\(^n\) peptides, b POH\(^n\) complex with 3 Ang-(1-7)\(^n\) peptides, c PNH\(^a\) in complex with 3 Ang-(1-7)\(^n\) peptides and b) POH\(^a\) at in complex with 3 Ang-(1-7)\(^n\) peptides
Complexes stabilityPAMAM-NH/Ang-(1-7) complexTo identify global structural changes due to complex formation, the RMSD and the R\(_\) at bound state were measured and compared with that of the free structures. It can be seen in Fig. 14ac that the binding of peptides stabilizes the dendrimer structure according to RMSD values. Meanwhile, as can be seen in Fig. 14bd, the radius of gyration of the dendrimer and the peptides are not importantly affected by the binding.
Fig. 14
Structural stability changes of peptide or PAMAM-NH upon binding. RMSD of structures in complex compared with same structures free in solvent for a neutral pH and c acidic pH. R\(_\) of structures in complex compared with same structures free in solvent at b neutral pH and d acidic pH
To test complex stability, the distance between the center of mass (COM) of the dendrimer and each peptide was measured. As showed in Figs. 15a, 16a, and 18a, under neutral pH conditions, 2 peptides remained stable bonded during the simulation in the dendrimer internal domain around a distance of 1 nm and a third peptide remained stable bonded at the dendrimer surface around a distance of 3 nm, peptides remain mainly in the same initial sites. The dendrimer kept an open hourglass geometrical shape during the whole simulation time, allowing at least two peptides to be near the core. As shown in Figs. 15d and 22b, under acidic pH conditions, from the 3 peptides initially at around 1 nm in the dendrimer internal domain, one remained very close to the core during most of the simulation time, two went out very quickly and stayed near the superficial zone, even leaving the dendrimer at certain points. From the two peptides in the dendrimer surface, one went to the back of the core at the last 50 ns of the simulation, interacting with the other peptide in the core. It appears that for two peptides, transitions from in to out the dendrimer had low energetic barriers or was an unstable state, in such a way that it is easy to get out, meanwhile, the third peptide appears to be in a stable deep minimum that forces it to stay close to the dendrimer core, but further research is required to confirm this hypothesis, due to the system flexibility, a MTD simulation could give us more insight about this process. Whith these results we can conclude that only one peptide remains stably bonded at the core of the dendrimer at acidic pH during the whole MD simulation.
Fig. 15
[PNH-A]\(^n\)) stability. a Distance from dendrimer COM to Ang-(1-7) peptides, b peptide coverage according to SASA values and c number of hydrogen bonds between dendrimer and peptides. # B-D refers to the different Ang-(1-7) peptides bonded to dendrimers. Grey dashed line represents the R\(_g\) of the dendrimer as a reference for dendrimer periphery
Fig. 16
PAMAM-NH/Ang-(1-7) coordination number along simulation time at a neutral pH and b acidic pH. Coordination mean the number of atoms from the peptides that are within 0.3 nm of the dendrimer. # B-D refers to the different Ang-(1-7) peptides bonded to dendrimers. Grey dashed line represents the threshold agreed that divides coupled from uncoupled states. A Bezier based smooth curve was applied for the sake of visibility
To test the protection capabilities of each dendrimer, the percent coverage of peptides due to binding, was measured. The free peptide in solution was taken as a reference, representing a molecule that is 100 % accessible to the solvent. As presented in Fig. 15b, in the case of the [PNH-A]\(^n\), an average of 54% of coverage was found, confirming PNH\(^n\) drug protection capabilities. As presented in Fig. 15e, in the case of the [PNH-A]\(^a\), an average of 20% of coverage was found, a much lower percentage than in the case of neutral pH. This is reasonable considering that 2 peptides are in the superficial zone during most of the simulation time and even leave the dendrimer at certain points. If we consider only the peptide that remains at the core, it still has less coverage percentage compared with the ones at the core at neutral pH, probably because due to the open structure at acidic pH, cavities are also deferentially hydrated. It could mean that the peptide might be a little more accessible to water but due to its localization inside the dendrimer, it is not necessarily accessible to peptides.
The peptide protection is especially relevant for bonds susceptible to hydrolytic attack by endopeptidasas, as schematized in Fig. 17. Once formed, Ang-(1-7) is rapidly hydrolyzed, especially by ACE [56] and dipeptidyl peptidase 3 (DPP3) [11]. Here, we found a 46 \(\%\) of [PNH-A]\(^n\) coverage on the region attacked by DPP3 and a 73 \(\%\) of [PNH-A]\(^n\) coverage on the region attacked by ACE. This result is important because the very short half-life of Ang-(1-7) in the circulation is primarily accounted for peptide metabolism by ACE [57].
Fig. 17
Proteolytic cleavage of Ang-(1-7) by ACE and DPP3
The main type of HBs formed between Ang-(1-7) and hydroxyl-terminated PAMAM dendrimer are presented in Fig.s S11 -S12, the number of NHBs through simulation time is presented in Fig. 15cf and detailed occupancy in Tables 5 and 6. At neutral pH, a number of 5.4 HBs in average are formed between PNH\(^n\) and Ang-(1-7) peptides, during the whole simulation time. The most populated HBs interactions are between core atoms, internal branches and terminal groups of PNH\(^n\) with PRO7, ASP1 and TYR4 aminoacids in the peptide. Side chain of TYR4 in one of the peptides behaves as a donnor interacting with the amide Oxigen atom from the internal dendrimer as an acceptor during half of the simulation time; all the peptide keeps its interaction between its negative charged C-terminal and the main Nitrogen of the positively charged primary amine in the dendrimer terminal groups during almost half of the simulation time; ASP1 negative side chain interacts with the protonated primary amine of the dendrimer terminal groups during at least 40% of the time and also side chain of TYR4 interacts with the amide NH group from the internal dendrimer. The characteristic ASP1-TYR4/ASP1-VAL3 HB found in Ang-(1-7) metadynamics is still frequently formed (50 \(\%\) of the simulation time) compared with its formation free in solvent (40 \(\%\) of the simulation time), this implies that these intra peptide HBs are at least equally broken in favor of a peptide/dendrimer interaction (Table 5) compared with peptide/waters interaction. At acidic pH, a number of 2.5 HBs in average are formed between PNH\(^a\) and Ang-(1-7) peptides, during the whole simulation time. The most populated HBs interactions are between internal branches of PNH\(^n\) with ASP1 side chain in the peptide, but with much low occupations than in the case of neutral pH.
Table 5 Main interactions (HBs) formed between Ang-(1-7) peptide and PNH\(^n\). Only occupancies above 35% are shown. SegA correspond to dendrimer and SegB-SegD to peptidesTable 6 Main interactions (HBs) formed between Ang-(1-7) peptide and PNH\(^a\). Only occupancies above 10% are shown. SegA correspond to dendrimer and SegB-SegD to peptidesAccording to Fig. 19a, at neutral pH, TYR4 and ILE5 residues, interacted preferably with internal dendrimer groups rather than with the terminal groups, which was expected due to its hydrophobic/partially hydrophobic nature. On the other hand, ASP1 and PRO7 residues, interacted preferably with terminal dendrimer groups rather than with the internal groups, which was expected due to its negative charged side-chain/C-terminal groups. ARG2, VAL3 and HIS6 interacts similarly with terminal or internal dendrimer groups but in a less frequent way. As expected, arginine residue was more exposed to solvent due to its charged aminoacid side chain. According to Fig. 19b, at acidic pH, ASP1, ARG2, TYR4 and HIS6 keep in contact preferably with internal dendrimer groups. Due to a more open dendrimer cavities, ASP1 and ARG2 are able to enter and interact with internal polar groups compared with the neutral case. Due to its hydrophobic/partially hydrophobic nature TYR4 keeps in contact preferably with internal dendrimer
留言 (0)