
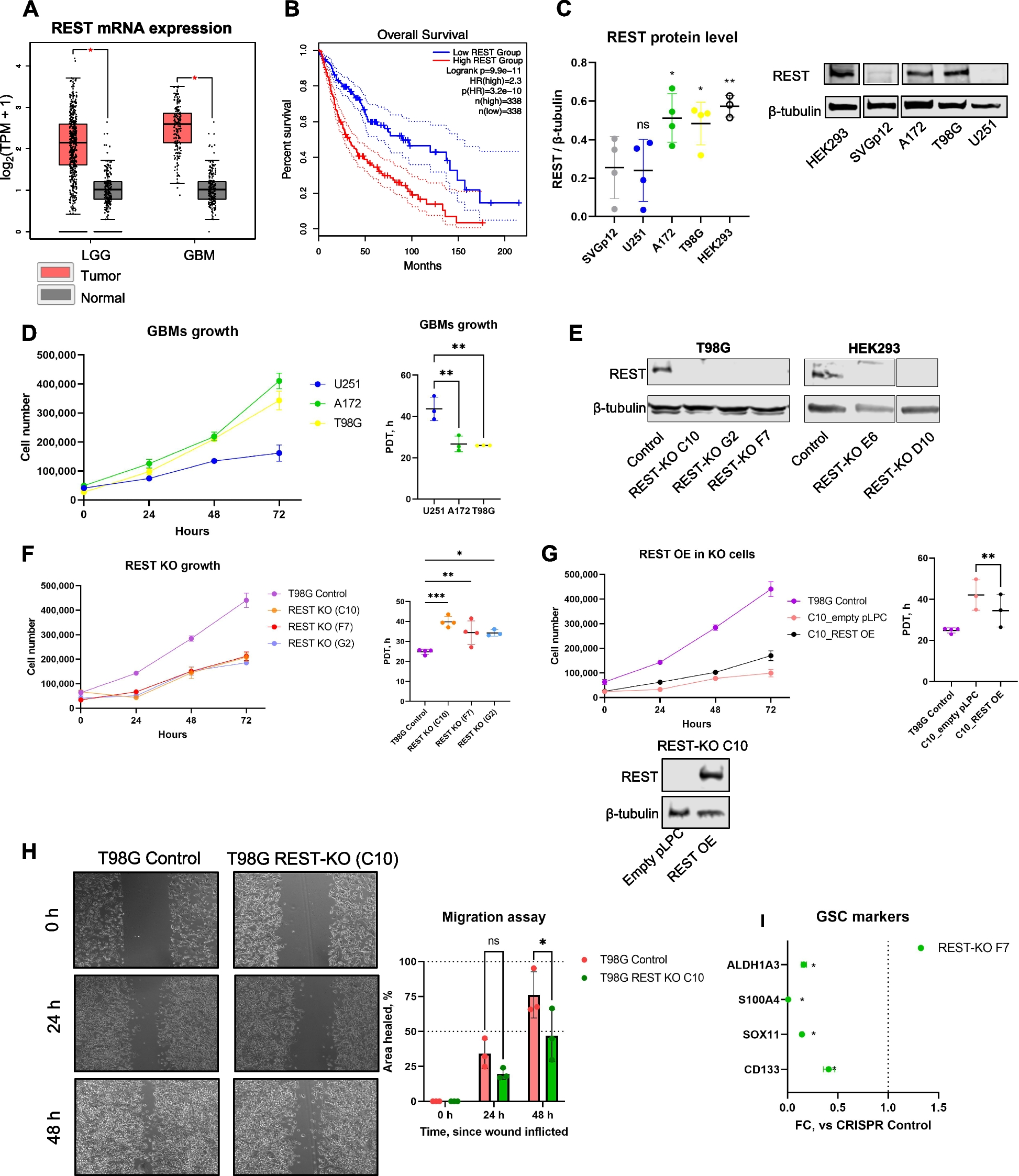
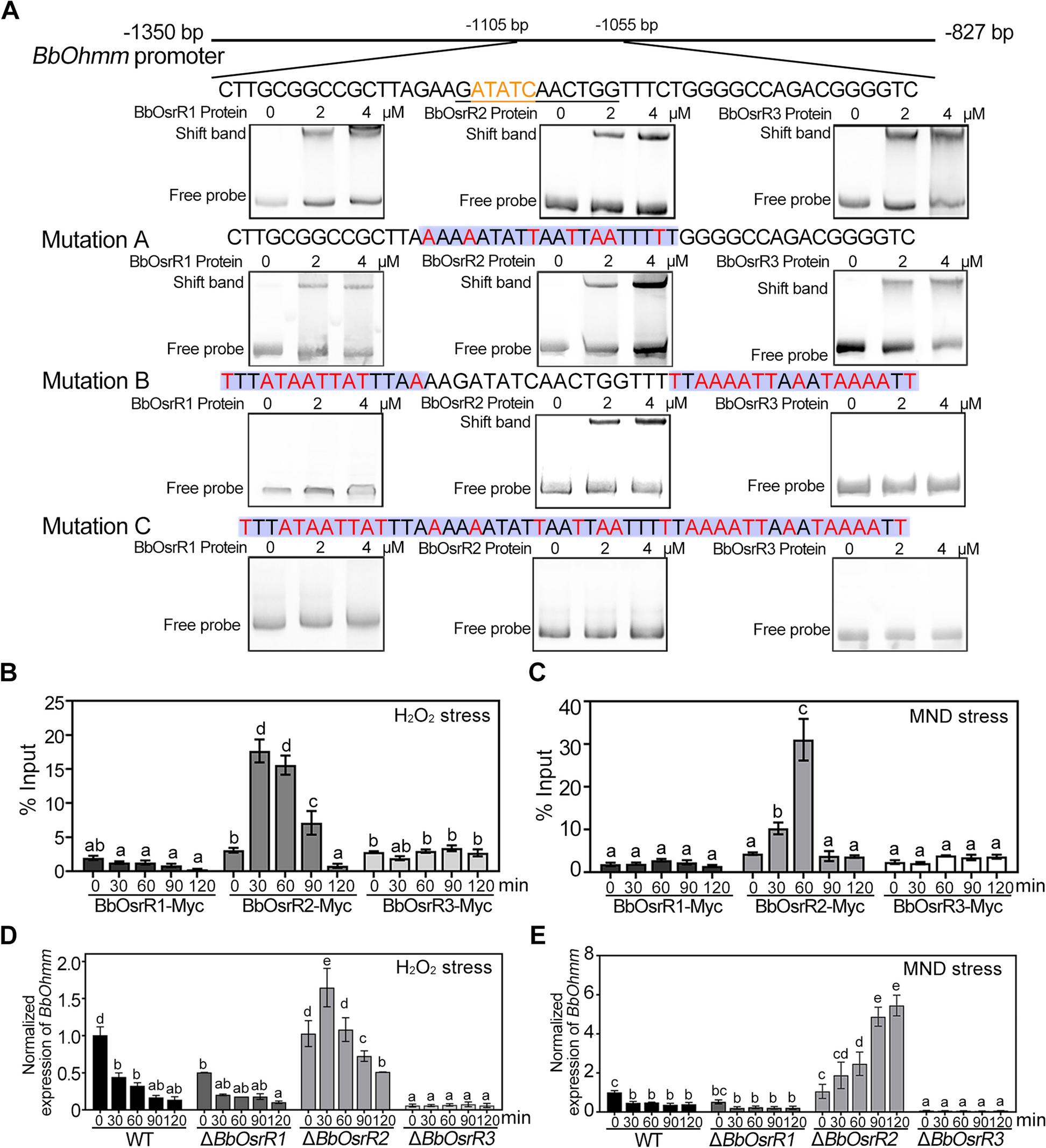
記住我








Using immunofluorescence microscopy (IFM), we investigated the subcellular localization of seven endogenous subunits of the SRCAP and p400/Tip60 chromatin remodeling complexes during cell division in fixed HeLa cells: CFDP1 (SRCAP complex), MRG15, p400 and Tip60 (p400/Tip60 complex), BAF53a, GAS41, and YL1 (both complexes).
As expected, for each chromatin remodeling subunit (CRS) tested, the antibody staining decorated the interphase nuclei (Additional file 1: Fig. S1). In addition, we observed a specific staining pattern at the mitotic apparatus (centrosomes, spindle, and midbody) during mitotic progression, with the exception of MRG15 that failed to show accumulation to the mitotic structures (Fig. 2). In particular, BAF53a, CFDP1, GAS41, and YL1 were found at the spindle in both metaphase and/or anaphase; Tip60 localized at the centrosomes and also showed a kinetochore accumulation, as previously reported [33]. Finally, all these subunits were also found to the midbody (Fig. 2 and Table 1).
Fig. 2
Localization of SRCAP and p400/Tip60 chromatin remodeling subunits (CRS) to the centrosomes, spindle, and midbody in HeLa cells. Fixed HeLa cells stained with DAPI (blue), anti-CRS (red), and anti-α-tubulin (green). BAF53a, CFDP1, GAS41, and YL1 fluorescent signals were found at the spindle in both metaphase and/or anaphase; Tip60 localized at the centrosome and kinetochores (particularly evident at anaphase). In telophase, all the tested CRS were found at the midbody. Scale bar = 10 μm
Table 1 Localization of CRS to the mitotic apparatusThe antibodies against the CRS of interest were validated by both IFM and Western blotting (WB) on HeLa cells transfected with a specific siRNA mix targeting the transcripts of each gene encoding the CRS (see “Methods”). After RNAi knockdown, the total amount of each CRS decreased, as well as the fluorescence intensity of nuclei, spindles, and midbodies (Fig. 3, Additional file 2: Fig. S2).
Fig. 3
Validation of antibodies against the CRS. A Total CRS amount assessed by Western blotting in RNAi-treated and mock-treated cells. B Histograms showing the fluorescence intensity of immunostaining in RNAi-treated and mock-treated cells. The colors of bars are referred to Fig. 1B. Bars with different dashed lines indicate SRCAP complex-specific subunits (CFDP1) or p400/Tip60-specific subunits (p400 and Tip60). Plain colored bars indicate subunits common to both complexes (BAF53a, GAS41, and YL1). The fluorescence intensity showed a decrease ranging from about 60 up to 75% in RNAi-treated HeLa cells compared to the mock-treated cells. Fluorescence intensity was assessed using the ImageJ software and statistical significance was verified by T- test. C Examples of IFM staining. The fluorescence intensity of BAF53a and Tip60 clearly decreased in RNAi-treated compared to mock-treated cells. Scale bar = 10 μm
The subcellular localizations of the endogenous CRS found in HeLa cells, were also examined in human MRC5 fibroblast-derived cell line. Here again, in addition to chromatin, the CRS accumulated at different sites of the mitotic apparatus (Additional file 3: Fig. S3). These results suggested that the mitotic localizations of the CRS reflect intrinsic properties of the proteins, independently of the cell type.
The midbody associations of CRS were also evaluated on isolated midbodies from HeLa cells using IFM and WB. In addition to the CRS analysed in fixed HeLa cells (Fig. 2), we tested Pontin and Reptin, which were found to localize to the spindle and midbody [24,25,26, 29]. We also tested the histone variant H2A.Z, which replaces the canonical histone H2A, due to the action of SRCAP and p400/Tip60 complexes [23]. As shown in Fig. 4, all the proteins under investigation were found to the isolated midbodies by both IFM and WB, with the exception of MRG15 that was detected only by WB. By contrast, the ISWI remodeler [34] was not found. Interestingly, the localization of Tip60 at the isolated midbodies was widely distributed in early telophase and became gradually more restricted in late telophase, similarly to that of other known cytokinesis regulators [35].
Fig. 4
Localization of CRS on isolated midbodies. A Immunolocalization. Fixed preparations of isolated midbodies were stained with DAPI (blue), antibodies against the protein of interest (red), and anti-α-tubulin (green). No DAPI staining was detected. The antibody staining for each tested protein clearly decorated the isolated midbodies. Scale bar = 5 μm. B Detection of CRS by Western blotting on midbody extracts from HeLa cells. Aurora B and α-tubulin were used as positive controls. ISWI was not detected (negative control)
Taken together, these findings revealed that the subcellular localization of CRS tested is dynamic during cell division, in that they relocate to different structures of the mitotic apparatus. Remarkably, SRCAP and p400/Tip60 complexes belong to the INO80 family of ATP-dependent chromatin remodelers, whose main function is to govern the deposition of the variant histone H2A in evolutionary distant species [23], thus such a massive relocation to the mitotic apparatus was not obvious.
RNAi-mediated depletion of SRCAP and p400/Tip60 complex subunits affects mitosis and cytokinesis in HeLa cellsNext, we examined the functional relevance of the CRS relocation to the mitotic apparatus, by investigating the progression of cell division after their RNAi-mediated depletion. These experiments were performed by transfecting HeLa cells with specific siRNAs against the transcripts encoding the following CRS: BAF53a, CFDP1, GAS41, MRG15, p400, Tip60, and YL1. Two negative control samples were considered: scramble siRNAs transfected Hela cells and mocktreated HeLa cells (see “Methods”).
As shown in Fig. 5A–E and Table 2, depletion of each CRS tested gave rise to cells showing five major classes of cell division defects: multipolar spindles (MS), chromosome misalignments in metaphase (CM), chromatin bridges (CB), long intercellular bridges (LIB), and multinucleated cells (MC). The strength of the observed defects varies between CRS, but a clear increase of mitotic abnormalities was observed which in most cases is statistically significant compared to the control samples.
Fig. 5
RNAi mediated depletion of CRS affects cell division in HeLa cells. Cells were stained with DAPI (blue) and anti-α-tubulin (green). Scale bar = 10 μm. Five classes of defects were categorized. A Histograms showing the quantitative analysis of cell division defects. Colors are referred to Fig. 1B. Bars with two different dashed lines indicate SCRAP complex-specific subunits (CFDP1) or p400/Tip60-specific subunits (MRG15, p400, and Tip60). Plain colored bars indicate subunits common to both complexes (BAF53a, GAS41, and YL1). B Mock, left panel; multipolar spindles (MS), middle panel; chromosome misalignments (CM), right panel. C Mock, left panel; chromatin bridges (CB) right panel. D Mock, left panel; long intercellular bridges (LIB) right panel; no DAPI-stained trapped chromatin was observed. E Mock, left panel; multinucleated cells (MC), right panel. Three independent experiments were performed. The quantitative analysis of defects scored in RNAi-treated and control cells (Table 2) is based on the following numbers: at least 100 prometaphases and metaphases for MS, 70 metaphases for CM and ASM, 300 telophases for LIB and CB, and 5500 interphases for MC. Experimental groups were compared with control groups (mocks and scrambles) by Fisher’s exact test.*=P < 0.05; **=P < 0.005; and ***=P < 0.0005
Table 2 Cell division defects found in CRS-depleted HeLa cellsDepletion of GAS41, p400, Tip60, and YL1 resulted in a significant increase of metaphases showing multipolar spindles (Fig. 5A, B): 12.77, 11.95, 10.77, and 10.49%, respectively, compared to mock and scramble controls. CFDP1 depletion did not result in an increase of multipolar spindle, but led to 32% of metaphases with monopolar spindles (Additional file 4: Fig. S4).
A significant increase of metaphases showing CM (Fig. 5A, B) was detected in BAF53a, CFDP1, GAS41, MRG15, Tip60, and YL1 depleted cells: 29, 23.9, 31.6, 7.16, 29.8, and 35% respectively. A high percentage of telophases with CB (Fig. 5A, C) was found only in BAF53a and CFDP1 depleted cells, while milder effects were seen in MRG15 and Tip60 depleted cells, and no significant effects were observed for the other subunits tested. Notably, mitotic defects in terms of lagging chromosomes were seen after depletion of Pontin [29].
Cytokinesis was also perturbed by depletion of all CRS under investigation. Reduction of BAF53a, GAS41, MRG15, TIP60, and YL1 amounts clearly resulted in dysfunctional abscission, leading to a significant increase in the percentage of figures showing LIB: 30.27, 9.41, 3.19, 4.88, and 8.32% respectively (Fig. 5A, D). Consistently, after depletion of the aforementioned CRS, the intercellular distance during cytokinesis increased (Additional file 5: Fig. S5). Finally, the depletion of all the subunits tested, with the exception of GAS41, also produced a statistically significant increase of MC (Fig. 5A, E).
In summary, a significant increase of mitosis and cytokinesis defects was observed after depletion of SRCAP and p400/Tip60 complex subunits, suggesting that their relocation to mitotic apparatus reflects functional roles in cell division.
BAF53a andTip60 interact with each other and with cytokinesis regulators in telophase synchronized cellsIn a previous report, we have found that the ATPase SRCAP interacts with a plethora of cytokinesis regulators and positively controls their recruitment to the midbody [31].
To further investigate the contribution of SRCAP and p400/Tip60 complex subunits to cell division control, we carried out co-immunoprecipitation (co-IP) assays to test interactions at the midbody between two CRS, BAF53a and Tip60, and crucial regulators of cytokinesis. We focused on Alix, anillin, Aurora B, CEP55, citron kinase (CIT-K), MKLP2, and spastin, seven well-known midbody-associated proteins that ensure proper cytokinesis [7, 10,11,12,13,14,15, 20]. We also tested α-tubulin, a main structural component of spindle and midbody and MRG15.
We choose BAF53a and Tip60 for several reasons: (i) in HeLa and MRC5 cells they both colocalized at the midbody with a sharp fluorescence signal mapping to the central zone (Figs. 2, 3, and 4; Additional file 3: Fig. S3 and Table 1); (ii) their depletion affected cytokinesis, with BAF53a depletion producing a strong effect on the formation of binucleated cells and long intercellular bridges (Fig. 5); (iii) BAF53a has structural features common to actin [23], a contractile ring component required for cytokinesis [36]; (iv) Tip60 is an acetyl-transferase that interacts and acetylates Aurora B in metaphase [33]; (v) both BAF53a and Tip60 coding genes have been implicated in cancer [37, 38]; (vi) The BAF complex subunits are implicated in a group of genetic diseases called BAFopathies [39].
Co-IP assays were performed on chromatin-free protein extracts from the cytoplasmic fraction of HeLa cells synchronized in telophase (see “Methods”), using antibodies against BAF53a and Tip60, previously validated by IFM and WB (Fig. 3). Telophase synchronization was followed by subcellular fractionation assays to recover the cytoplasmic component (S2 fraction) and segregate away the chromatin-associated components (see “Methods”).
As shown in Fig. 6A, from the comparison between the negative control (−BAF53a and −Tip60) and the IP (+BAF53a and +Tip60), BAF53a and Tip60 interacted with each other, consistent with their co-localization to the midbody (Fig. 2), and with all cytokinesis regulators tested, but Alix. Moreover, both BAF53a and Tip60 interacted with MRG15 and α-tubulin.
Fig. 6
Interactions between CRS and cytokinesis regulators. A Immunoprecipitation of protein extracts from cytoplasmic fraction of telophase synchronized HeLa cells (S2 fraction). IP sample immunoprecipitated with BAF53a or Tip60 antibodies (+anti-BAF53a; +anti-Tip60) were compared to negative controls (−anti-BAF53a or −anti-Tip60). Anillin, Aurora B, CEP55, CIT-K, MKLP2, MRG15, spastin, and α-tubulin were found in the IP samples immunoprecipitated with BAF53a or Tip60 antibodies, but not in the negative controls. Three independent IP experiments were performed. IN = input, IP = immunoprecipitation. Synchronization of the HeLa cells was performed according to Messina et al. [31]. B Histograms showing the quantitative analysis of mislocalizations of cytokinesis regulators at the midbody in mocks and BAF53a or Tip60 depleted cells. Three independent experiments were performed and at least 300 telophases were scored in both RNAi-treated and control cells. *=P<0.05,**=P<0.005, ***=P<0.0005. C Examples of mislocalization of cytokinesis regulators in mock and BAF53a or Tip60 depleted cells. From left to the right: DAPI (blue), anti-α-tubulin (green), cytokinesis regulators (red), and merge. Scale bar = 10 μm
The proper localization of cytokinesis regulators to the midbody is affected by depletion of BAF53a or Tip60The aforementioned results suggested that BAF53a and Tip60 may contribute to the recruitment of cytokinesis regulators to the midbody, with a role in the regulatory pathway that controls the final step of cytokinesis. To test this hypothesis, we asked whether depletion of BAF53a and Tip60 affects the recruitment at the midbody of the same cytokinesis regulators tested in the co-IP experiments. As shown in Fig. 6B, C and Table 3, the results of three independent replicates showed that the midbody localization pattern of the cytokinesis regulators under investigation was impaired in BAF53a and Tip60 depleted HeLa cells. In particular, a strong alteration of the localization pattern was observed for anillin, Alix, CEP55, MKLP2, and spastin.
Table 3 Mislocalization of cytokinesis regulators in CRS-depleted HeLa cellsThe proper localization of BAF53a and Tip60 to the midbody is affected by the inhibition of Aurora B kinase activityAn important question is how SRCAP and p400/TIP60 chromatin remodelers are recruited to the midbody. The results of our IP assays indicated that BAF53a and Tip60 interact with Aurora B kinase, a key regulator of mitosis and cytokinesis which interacts with several microtubule-binding proteins [10, 14, 34].
To understand the relationships between Aurora B and the CRS under investigation, we asked whether the inhibition of its kinase activity affects the recruitment of both BAF53a and TIP60 to the midbody. We also tested the SRCAP ATPase, since it was found to control Aurora B localization to the midbody [31]. To inhibit of the Aurora B kinase activity, the ZM447439, a specific selective ATP-competitive inhibitor was used [40]. We found that the midbody localizations of BAF53a, Tip60, and MKLP1 are significantly affected in HeLa cells treated for 35 min with ZM447439, compared to that in DMSO-treated controls (Fig. 7A–C and Table 4). No effect was seen on Aurora B and on SRCAP localization. The use of Barasertib, another inhibitor of Aurora B kinase activity [41], produced similar results (Fig. 7D, E), the only difference being a slight effect on the localization of Aurora, with a p-value at the limit of significance. As control of inhibition efficiency, phosphorylation of the histone H3 during mitosis was assessed (Fig. 7C, E). Together, the results obtained using two different inhibitors strongly suggested that the Aurora B kinase activity is required for proper localization of BAF53a and Tip60 to the midbody and also support the hypothesis of a cross-regulation occurring during cytokinesis between chromatin remodelers and cytokinesis regulators.
Fig. 7
Inhibition of Aurora B kinase activity with ZM447439 and Barasertib affected the localization of BAF53a and Tip60 to the midbody. A Treatment with ZM447439: from left to right: DAPI (blue), anti- α-tubulin (green), tested protein (red), and merge. Anti-MKLP1 and anti-Aurora B immunostaining were used as positive and negative controls, respectively. Scale bar = 10 μm. The localization pattern of BAF53a and Tip60, as well as that of MKLP1 were affected, while no effect was seen for SRCAP and Aurora B. B The effects found with ZM447439 treatment are summarized in the graph. At least 300 telophases were scored in three independent experiments for both treated cells and control HeLa cells. C To test the effectiveness of ZM447439 inhibition, phosphorylation of the histone H3 (target of Aurora B kinase) during mitosis was evaluated; α-tubulin was used as loading control. D The effects found with Barasertib treatment are summarized in the graph. At least 200 telophases were scored in two independent experiments for both treated cells and control HeLa cells. A slight effect on Aurora localization was seen at the limit of significance values. E To test the effectiveness of Barasertib inhibition, phosphorylation of the histone H3 (target of Aurora B kinase) during mitosis was evaluated; α-tubulin was used as loading control. *=P<0.05, **<=P<0.005, ***=P<0.0005
Table 4 Treatment with ZM447439 (ZM) and Barasertib (BS) inhibitorsThe proper localization of BAF53a and CFDP1 to the midbody is affected by SRCAP depletionA further question regarding the mechanisms underpinning the recruitment of CRS to the mitotic apparatus is whether they maintain their interactions during the relocation or, alternatively, move independently. We have shown that BAF53a and Tip60 colocalize and interact with each other and with MRG15 at the midbody (Figs. 2, 4 and 6), suggesting that the three subunits are still interacting during their “trip” from interphase chromatin to the mitotic apparatus. To further investigate this aspect, we studied the localization to the midbody of CFDP1 (SRCAP complex specific subunit), BAF53a, GAS41, YL1 (subunits shared by both the SRCAP and p400/Tip60 complexes), p400, and Tip60 (p400/Tip60 complex specific subunits) in HeLa cells depleted for SRCAP, the main platform of the homonymous complex. As shown in Fig. 8, SRCAP depletion clearly affects the localization of both CFDP1 and BAF53a, but not that of p400, Tip60, GAS41, and YL1.
Fig. 8
SRCAP-dependent localization of BAF53a and CFDP1 to the midbody. A Examples of mislocalization. DAPI (blue), anti-α-tubulin (green), tested CRS (red), and merge. Scale bar = 10 μm. B The graph summarized the results. At least of 300 telophases were scored in two independent experiments for both treated cells and control HeLa cells. ***=P<0.0005
Subcellular localization and RNAi-mediated depletion of MRG15, Tip60, and YETI, three subunits of the D. melanogaster Tip60 complexIn past studies, we found that the lack of D. melanogaster YETI protein affects chromosome organization and impairs cytokinesis [42,43,44,45,46]. YETI is the Drosophila ortholog of CFDP1 [47,48,49] and belongs to the dTip60 chromatin remodeling complex [50] which includes subunits sharing high sequence and function conservation with those of SRCAP and p400/Tip60 complexes. Recently, we found that DOM-A, the functional SRCAP orthologue in D. melanogaster, is recruited to centrosomes and midbody in Drosophila S2 cells, and its depletion affects both mitosis and cytokinesis [31]. Here, we asked whether this behavior is shared by other dTip60 complex subunits. To answer this question, we used IFM to study the distribution of MRG15, Tip60, and YETI in S2 cells. These subunits are structurally and functionally orthologous to human MRG15, Tip60, and CFDP1, respectively. As shown in Fig. 9, high levels of identity and similarity are found at specific functional domains of these proteins (BCNT, TUDOR, MRG, CHROMO; MOZ_SAS).
Fig. 9
Sequence conservation of Drosophila MRG15, dTip60, and YETI proteins with their human orthologs. A) BLAST alignments; B) High identity levels found in specific functional domains of CFDP1, MRG15 and Tip60
The results of this experiment showed that MRG15, Tip60, and YETI, apart from to their canonical location in chromatin, are found at the midbody (Fig. 10), similarly to their human orthologs. In addition, MRG15 was localized to the centrosomes. Thus, the recruitment of CRS to the mitotic apparatus appears to be an evolutionary conserved phenomenon.
Fig. 10
Immunostaining showing the localization of MRG15, Tip60, and YETI to the mitotic apparatus in D. melanogaster S2 cells. Cells were stained with DAPI (blue), anti-α-tubulin (green), and anti-MRG15, anti-Tip60, and anti-YETI (red). Scale bar = 5 μm. In addition to interphase nuclei, the anti-MRG15 decorated the centrosomes (metaphase) and midbody (telophase), while anti-Tip60 and anti-YETI staining was detected on the midbody
Next, we examined the phenotypes of S2 cells after RNAi against MRG15, Tip60, and YETI. The depletion of these subunits indeed resulted in five categories of defects (Fig. 11A–F and Table 5) comparable to those observed in HeLa cells depleted for their corresponding orthologs: multipolar spindles (MS), chromosome misalignments in metaphase (CM), chromatin bridges (CB), long intercellular bridges (LIB), and multinucleated cells (MC). The actual effectiveness of RNAi treatments was tested by IF and WB assays (Fig. 11G, H). The depletion of MRG15, Tip60, and YETI affected mitosis in terms of cells with MS and CM. Mild and yet statistically significant effects were seen on CB. MRG15 and YETI depletion also affected abscission with 9% and 11.54% of LIB, respectively. Finally, YETI depletion resulted in 4% of MC.
Fig. 11
RNAi-mediated depletion of MRG15, Tip60, and YETI in S2 cells affects mitosis and cytokinesis. Examples of mitotic defects found in MRG15, Tip60, and YETI depleted S2 cells and their quantification. DAPI staining is shown in blue, α-tubulin in green. RNAi knockdown experiments were performed by transfecting S2 cells with specific siRNAs against each protein (see “Methods”). Scale bar = 5 μm. Five classes of defects were considered: A multipolar spindle (MS); B chromosome misalignments in metaphase (CM); C chromatin bridges (CB); D long intercellular bridges (LIB); E multinucleated cells (MC); F Quantitative analysis of defects scored in RNAi-treated and control cells (Table 5) is based on the following numbers: at least 100 prometaphases and metaphases for MS, 70 metaphases for CM, 300 telophases for LIB and CB, 5500 interphases for MC. Three independent experiments were performed. G Percent of cells showing a decrease of fluorescence intensity at the midbody in MRG15, Tip60, or YETI depleted cells (black histograms) compared to the mocks (white histograms). The results are based on three independent experiments. H WB showing a decrease of total amount of MRG15, Tip60, and YETI proteins in RNAi-treated S2 cells compared to the mocks; a-tubulin is used as loading control
Table 5 Quantitative analysis of defects found in CRS-depleted S2 cells
留言 (0)