
記住我







Phosphate (Pi) homeostasis is precisely regulated. In adults, normal plasma Pi ranges from 0.80 to 1.45 mmol l−1[1], which is maintained by inter-organ interplay, including intestinal absorption, bone (de)mineralization, and renal excretion. The kidney is the key organ for fine-tuning Pi homeostasis; therefore, impaired renal function results in Pi imbalance, including hypophosphatemia (plasma Pi levels < 0.8 mmol l−1) and hyperphosphatemia (plasma Pi levels >1.4 mmol l−1). Hyperphosphatemia is closely associated with chronic kidney disease (CKD) in later stages of the disease and cardiovascular diseases [2].
Several Pi transporters have been identified in the renal proximal tubule, including sodium-phosphate cotransporters Npt2a (SLC34A1), Npt2c (SLC34A3), Pit1 (SLC20A1), and Pit2 (SLC20A2) (for a review, see [3,4▪,5]). The estimated relative contribution of Npt2a for renal Pi reabsorption is ∼70%, as evidenced by studies using brush border membrane vesicles isolated from Npt2a−/− mice [6]. Pathologically these mice are characterized by increased urinary Pi excretion and decreased plasma Pi concentration [7▪▪]. The contribution of Npt2c shows species differences and depends on the development stage, with more contribution to Pi reabsorption in juveniles compared to adults [8]. In humans, SLC34A3 mutations cause hereditary hypophosphatemic rickets with hypercalciuria (HHRH) [9,10]. In contrast, in mice, loss of renal Npt2c shows no prominent impact on Pi homeostasis [11▪], and Npt2a/c double knockout mice demonstrate a similar effect on urinary Pi excretion compared with Npt2a−/− mice [12], highlighting the small contribution of Npt2c for renal Pi reabsorption. Similarly, both Pit1 and Pit2 appear to have a small contribution to renal Pi transport [13,14]. Interestingly, a recent study localized Npt2b to the thick ascending limb of the kidney; however, its abundance was not regulated by dietary Pi[15] and further studies are needed to address the importance of this finding.
FGF23 and PTH are primary regulators for systemic Pi homeostasis, and both reduce renal Pi reabsorption via retrieval of Npt2a/c from the luminal membrane [16]. Additionally, FGF23 downregulates vitamin D, leading to reduced Pi absorption from the intestine, whereas PTH exerts the opposite effects: increasing vitamin D and intestinal Pi absorption [17]. On the other hand, plasma Ca2+ and Pi levels regulate PTH release from the parathyroid gland via the calcium-sensing receptor (CaSR) [18]. Although the exact mechanism for regulating FGF23 release is incompletely understood, it has been suggested that FGF23 production by bone is regulated in response to dietary Pi at the transcription, translation, and posttranslational levels [19]. Elevated plasma Pi level upregulate both, FGF23 and GALNT3 production, the latter an enzyme protecting FGF23 from proteolytic cleavage via posttranslational glycosylation [20]. The Chronic Renal Insufficiency Cohort study showed that FGF23 is an early biomarker during CKD progression, with elevated plasma FGF23 preceding the increase in PTH and Pi[21▪], implying that FGF23 is the principal Pi homeostasis regulator compared to PTH, at least in the early stages of CKD. The increased plasma FGF23 in the early stages of CKD, followed by an increase in PTH, is sufficient to prevent hyperphosphatemia until stage 4–5 CKD. This hypothesis is supported by animal studies showing that targeting FGF23 production impairs Pi homeostasis in CKD animal models [22,23]. High plasma FGF23 and Pi levels are independently associated with poor cardiovascular outcomes in patients with CKD, but the primary physiological function of FGF23 is to protect the body from hyperphosphatemia, which subsequently can cause detrimental cardio-renal consequences [24–26].
 Box 1:
Box 1: no caption available
ADAPTATION OF RENAL Pi TRANSPORTERS IN CHRONIC KIDNEY DISEASEIn CKD, renal function declines gradually with age, showing a reduction in nephron numbers and elevated FGF23 and PTH levels. These changes can potentially downregulate Npt2a/c expression. Supporting this theory, adenine-induced CKD animal models show markedly reduced Npt2a expression at the protein and mRNA levels [27–29]. Consistently, in rats and mice with reduced kidney function (5/6 Nx), a substantial reduction in Npt2a mRNA expression is also observed [30]. Additionally, Npt2a activity is affected by urine pH: more alkaline urine increases Npt2a activity. Notably, CKD is characterized by low urine pH, possibly associated with lower Npt2a activity. Therefore, these observations need to be considered when targeting renal Na+/Pi cotransporters as therapeutic strategies.
THERAPEUTIC STRATEGIES FOR LOWERING HYPERPHOSPHATEMIA IN CHRONIC KIDNEY DISEASETreating hyperphosphatemia in CKD is challenging due to sophisticated regulatory mechanisms for Pi homeostasis. Currently available treatment options include dietary Pi restriction, oral Pi binders, as well as niacin/nicotinamide [31–33]; however, all show severe limitations. Dietary Pi restriction and oral Pi binders are supposed to reduce Pi entry; however, the maladaptive upregulation of Pi uptake from the gastrointestinal tract limits their efficacy [34,35]. An alternative approach to lower intestinal Pi absorption is targeting Npt2b, responsible for >90% of active Pi uptake in the intestine [36]. Unfortunately, results from clinical trials indicate the limited efficacy of two newly developed Npt2b inhibitors (ASP3325; DS-2330b) for lowering plasma Pi in healthy volunteers and patients on hemodialysis [37▪,38▪]. Similarly, downregulation of Npt2b by niacin/nicotinamide also shows unsatisfactory results [39,40]. Compared to the Npt2b-selective inhibitor, a novel pan-phosphate transporter inhibitor (EOS789, targeting Npt2b, Pit1/2) shows a more potent serum Pi lowering effect with decreased FGF23 and PTH in rats with adenine-induced hyperphosphatemia [41▪▪]. Although a recent phase 1b clinical trial supports the safety of ESO789 in patients on hemodialysis [42], its efficacy needs to be confirmed by further studies. Inhibition of intestinal Na+/H+ exchanger isoform 3 (NHE3) by a nonabsorbable inhibitor, tenapanor, shows a serum Pi-lowering effect in patients on hemodialysis [43▪,44▪,45]. Mechanistically it was proposed that tenapanor inhibits paracellular rather than transcellular Pi absorption. In order to unravel such a mechanism we studied inducible intestinal epithelia cell-specific NHE3 knockout mice [46]. Of note, genetic deletion of intestinal NHE3 resulted in enhanced rather than reduced intestinal Pi uptake [47▪▪], implying that different mechanisms/conditions are causing the differences between pharmacological inhibition vs. genetic deletion. Last year the United States Food and Drug Administration denied the approval of tenapanor due to small effect of unclear clinical significance [48]; however, this decision has been appealed by Ardelyx, Inc.
PHOSPHATURIC EFFECT OF SODIUM PHOSPHATE COTRANSPORTER 2a INHIBITIONIn addition to reducing intestinal Pi absorption, promoting renal Pi excretion is another strategy for lowering plasma Pi. Until recently, two Npt2a-selective inhibitors (PF-06869206 from Pfizer; BAY-767 from Bayer) have been developed. The selectivity of these inhibitors for Npt2a has been confirmed in vitro[49▪,50▪▪] and in vivo[7▪▪,51▪▪]. Using opossum kidney (OK) cells, we showed a dose-dependent inhibition of Na+-dependent 32Pi uptake (half maximal inhibitory concentration, IC50 ∼ 1 mmol L−1) by PF-06869206, with a maximum inhibitory effect of ∼70% at 100 mmol L−1[7▪▪]. In wild type (WT) (C57Bl/6J) mice, the dose of 100 mg kg−1 PF-06869206 increases urinary Pi excretion by ∼6-fold (3 h period) compared to vehicle (median effective dose, ED50 ∼ 21 mg kg−1) [52▪▪]. Another study reported a similar phosphaturic effect of PF-06869206 where a dose of 500 mg kg−1 caused a ∼17-fold increase in the fractional excretion index (FEI) of Pi (4 h period) in WT mice [51▪▪]. Of note, this phosphaturic effect is consistently observed in different hyperphosphatemic animal models (FGF23−/− and GALNT3−/− mice), with a ∼9-fold and ∼2-fold increase in FEI of Pi (4 h time frame), respectively, in response to PF-06869206 at a dose of 300 mg kg−1 compared with vehicle [51▪▪]. Another Npt2a inhibitor, BAY-767, also showed a profound phosphaturic effect and a dose of 10 mg kg−1 resulted in a ∼1.7-fold increase in fractional urinary Pi excretion (16 h time frame) [50▪▪].
RESPONSES OF PLASMA Pi, PTH, AND FGF23 TO SODIUM PHOSPHATE COTRANSPORTER 2a INHIBITIONWill the phosphaturic effect of Npt2a inhibition reduce plasma Pi levels? Our studies employing PF-06869206 at a dose of 30 mg kg−1 showed a reduction in plasma Pi levels starting 30 min after oral administration in WT mice, with a maximum reduction at 2 h (−35%) and complete recovery after 24 h [7▪▪,52▪▪]. Another study in WT mice by Clerin et al.[51▪▪] showed that 300 mg kg−1 of PF-06869206 significantly reduced plasma Pi levels. Importantly, PF-06869206 showed similar plasma Pi-lowering effects in FGF23−/− (−20%), GALANT3−/− (−20%), and Npt2c−/− (−33%) mice 2–4 h after administration, the former two mouse models are hyperphosphatemic. In contrast, no effect was observed in Npt2a−/− mice, supporting the selectivity of PF-06869206 for Npt2a [7▪▪,51▪▪]. Another Npt2a inhibitor, BAY-767, at a dose of 10 mg kg−1, reduced plasma Pi levels (∼20%) in rats after 3 days of treatment [53].
As two predominant regulators of Npt2a membrane abundance, PTH and FGF23 [54,55] demonstrated distinct responses to PF-06869206. We found that PF-06869206 at a dose of 30 mg kg−1 reduced plasma PTH levels by ∼50% in mice after 3 h of administration [52▪▪]. Clerin et al.[51▪▪] also observed a reduction of PTH levels in mice (∼65%) at a dose of 300 mg kg−1 2–4 h after administration. In both studies, the reduction of PTH levels was fully recovered after 24 h. Three days of treatment with BAY-767 (10 mg kg−1) in rats reduced the PTH levels by 50% [53]. How do PTH levels decline upon Npt2a inhibition? The CaSR in the parathyroid gland acts as a plasma Pi sensor and thus modulates PTH secretion [18]. In contrast, plasma FGF23 levels were not affected by PF-06869206 [51▪▪,52▪▪]. Of note, BAY-767 reduced plasma FGF23 levels (∼25%) compared to the vehicle [53]. The inconsistency between PF-06869206 and BAY-767 in FGF23 responses is unknown and needs further study.
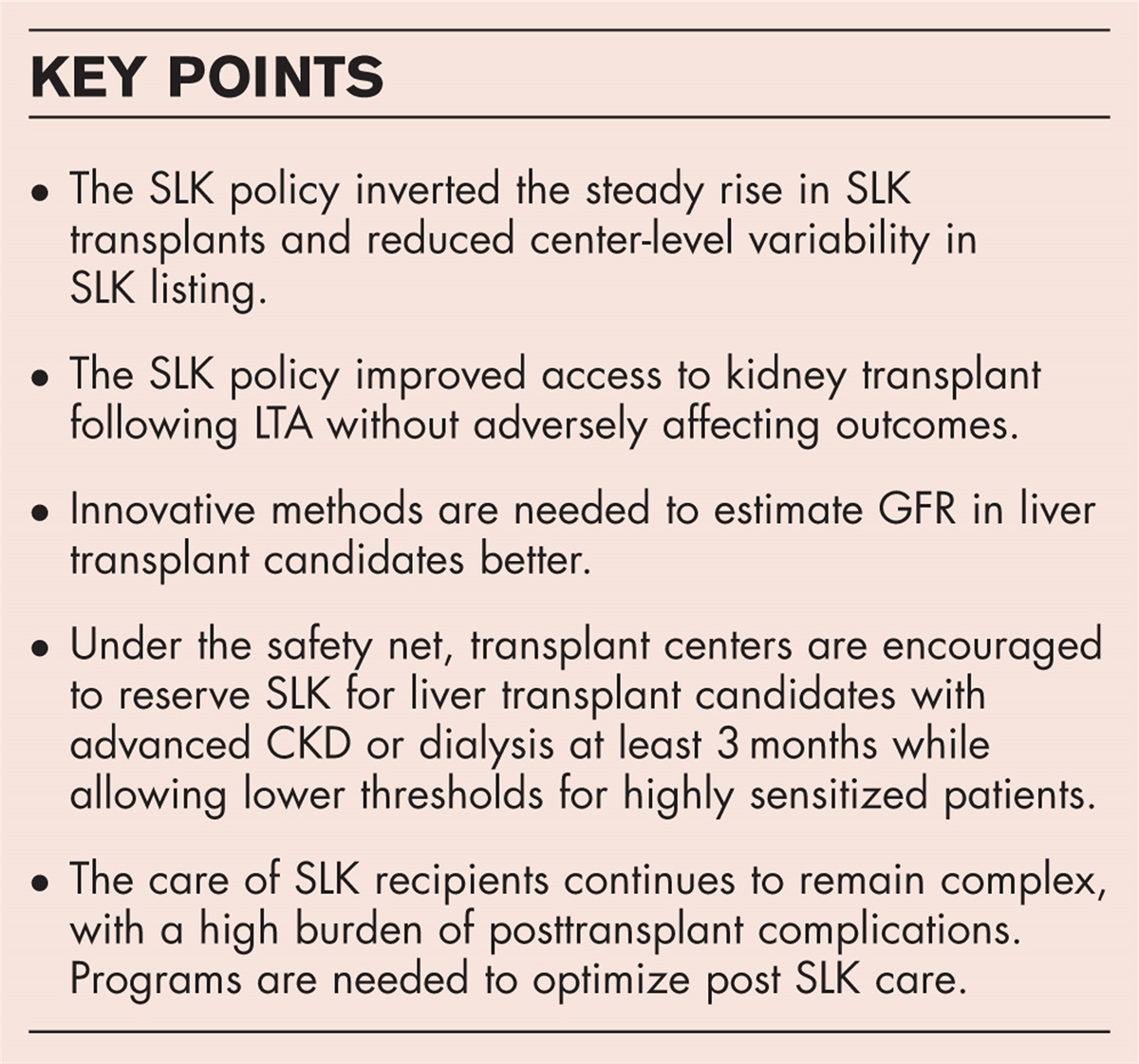
EFFICACY OF SODIUM PHOSPHATE COTRANSPORTER 2a INHIBITION IN CHRONIC KIDNEY DISEASE/REDUCED KIDNEY FUNCTIONIn 5/6 Nx mice, a model of reduced kidney function, acute administration of PF-06869206 increased urinary Pi excretion dose-dependently (Fig. 1a); however, the maximum phosphaturic effect at the dose of 100 mg kg−1 (compared to vehicle) was lower compared to sham mice (∼2-fold vs. ∼10-fold, respectively). After 3 h of administration of 100 mg kg−1, PF-06869206 significantly reduced plasma Pi (Fig. 1c) and PTH levels (Fig. 1d) in both 5/6 Nx and sham mice. Clerin et al.[51▪▪] treated 5/6 Nx rats with 300 mg kg−1 PF-06869206 for 8 weeks and observed higher FEI of Pi (∼2.5-fold) and lower plasma Pi (−15%) compared to vehicle-treated rats. However, 5/6 Nx rats lacked hyperphosphatemia, whereas elevated PTH and FGF23 levels were observed compared to sham rats. Surprisingly, long-term treatment with PF-06869206 did not affect plasma PTH or FGF23 level. So far, the efficacy of BAY-767 has not been investigated in CKD models.
 FIGURE 1: Inhibition of Npt2a with PF-06869206 affects urine and plasma parameters in mice with normal and reduced kidney function (5/6 Nx). An acute inhibition (3 h) of Npt2a with PF-06869206 resulted in a dose-dependent increase in urinary Pi (a) and Ca2+ (b) excretion in both sham and 5/6 Nx mice. The enhanced phosphaturia was associated with the acute reductions in plasma Pi (c) and PTH (d). PF-06869206 also induced a dose-dependent natriuresis (e) in both groups. Because of the absence of kaliuresis and persistence of dose-dependent natriuresis in Npt2a–/– mice (both data not shown), we assumed that the cause of natriuresis could be from the aldosterone sensitive segment of the distal nephron, where the epithelial sodium channel (ENaC) is located. In the further electrophysiological studies in the acutely split-open cortical collecting ducts of C57BL/6 mice, ENaC open probability was acutely inhibited (∼85%) by PF-06869206, possibly explaining the cause of natriuresis. A trace of continuous current is shown in (f), where dashed lines are the respective levels of current, with “o” denoting the open state and “c” denoting the closed state. Areas 1 and 2 under the bars over the continuous traces are shown below at expanded timescales. Figure 1a–e reused with permission from [52▪▪]. Figure 1f reused with permission from [7▪▪]. ∗P§P#PFigure 1 reprinted with permission from Biochem Soc Trans. 2022;50(1):439–446. CKD, chronic kidney disease; Npt2a, sodium phosphate cotransporter 2a.“OFF-TARGET” EFFECTS OF SODIUM PHOSPHATE COTRANSPORTER 2a INHIBITION
FIGURE 1: Inhibition of Npt2a with PF-06869206 affects urine and plasma parameters in mice with normal and reduced kidney function (5/6 Nx). An acute inhibition (3 h) of Npt2a with PF-06869206 resulted in a dose-dependent increase in urinary Pi (a) and Ca2+ (b) excretion in both sham and 5/6 Nx mice. The enhanced phosphaturia was associated with the acute reductions in plasma Pi (c) and PTH (d). PF-06869206 also induced a dose-dependent natriuresis (e) in both groups. Because of the absence of kaliuresis and persistence of dose-dependent natriuresis in Npt2a–/– mice (both data not shown), we assumed that the cause of natriuresis could be from the aldosterone sensitive segment of the distal nephron, where the epithelial sodium channel (ENaC) is located. In the further electrophysiological studies in the acutely split-open cortical collecting ducts of C57BL/6 mice, ENaC open probability was acutely inhibited (∼85%) by PF-06869206, possibly explaining the cause of natriuresis. A trace of continuous current is shown in (f), where dashed lines are the respective levels of current, with “o” denoting the open state and “c” denoting the closed state. Areas 1 and 2 under the bars over the continuous traces are shown below at expanded timescales. Figure 1a–e reused with permission from [52▪▪]. Figure 1f reused with permission from [7▪▪]. ∗P§P#PFigure 1 reprinted with permission from Biochem Soc Trans. 2022;50(1):439–446. CKD, chronic kidney disease; Npt2a, sodium phosphate cotransporter 2a.“OFF-TARGET” EFFECTS OF SODIUM PHOSPHATE COTRANSPORTER 2a INHIBITION
In addition to the dose-dependent phosphaturic effect, PF-06869206 also increased urinary Na+, Cl-, and Ca2+ excretion dose-dependently without affecting their plasma levels [52▪▪]. At a dose of 300 mg kg−1, PF-06869206 increased urinary Ca2+ excretion three-fold compared to vehicle in our studies (Fig. 1b), and Clerin et al.[51▪▪] observed a five-fold increase with the same dose. The possible mechanism for increased calciuria by PF-06869206 is the inhibition of Ca2+ reabsorption either in the proximal tubule (via the paracellular pathway) or the distal convoluted tubule (via TRPV5, transcellular). The latter may be affected by the decreased PTH levels observed after PF-06869206 treatment. On the other hand, PF-06869206 did not alter urinary excretion of K+, glucose, amino acids, or pH [52▪▪]. This implies that PF-06869206 does not lead to a generalized proximal tubular dysfunction, as seen with Fanconi syndrome. Like C57BL/6 mice, the urinary excretion of Na+ (Fig. 1e), Cl− and Ca2+ (Fig. 1b) was dose-dependently increased in response to PF-06869206 in mice with 5/6 Nx, whereas urinary excretion of K+, glucose, and pH were unaffected [52▪▪].
As a selective Npt2a inhibitor, we thought PF-06869206-induced natriuresis would be absent in Npt2a−/− mice; however, natriuresis was unaffected in Npt2a−/− mice in response to PF-06869206 [7▪▪]. Due to the lack of kaliuresis, we hypothesized that the observed natriuresis could be an off-target effect of PF-06869206 via inhibiting ENaC in the aldosterone-sensitive distal nephron. In the subsequent electrophysiological studies in acutely isolated and split-open cortical collecting ducts, the open probability of ENaC was ∼85% inhibited (Fig. 1f) in the presence of PF-06869206, giving a possible explanation for a natriuresis observed in Npt2a−/− mice. Blood pressure and total body Na+ levels are closely interconnected. However, Clerin et al.[51▪▪] observed no change in systolic blood pressure upon long-term treatment with PF-06869206 in 5/6 Nx rats, despite the presence of acute natriuresis and diuresis in 5/6 Nx mice [52▪▪]. Further studies are needed to determine the reason(s) for these differences.
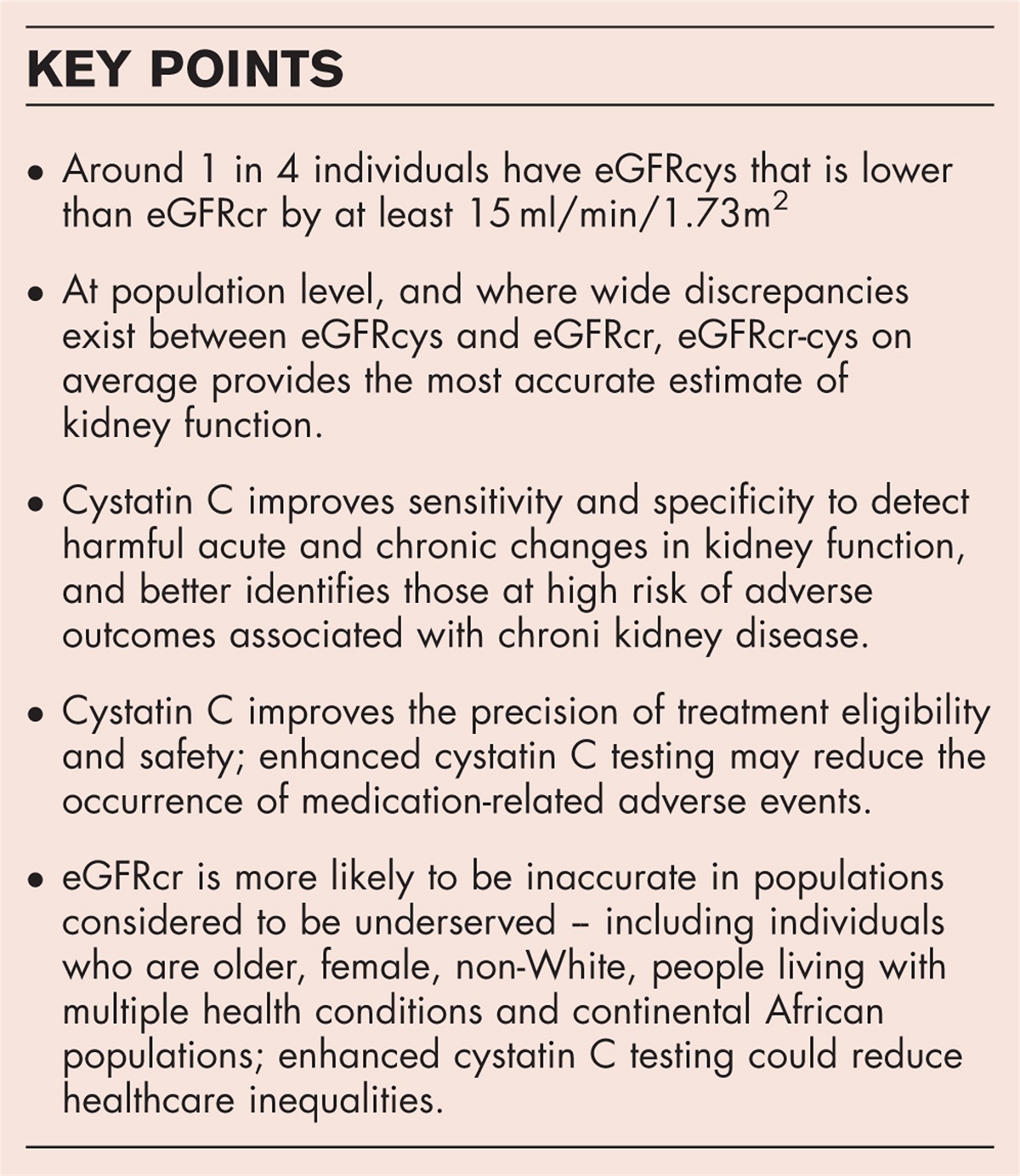
POTENTIAL CARDIOVASCULAR BENEFITS BY SODIUM PHOSPHATE COTRANSPORTER 2a INHIBITIONCalciuria is one of the critical pleiotropic effects caused by PF-06869206 and BAY-767. The imbalance of hormones and minerals (Pi and Ca2+) commonly seen in CKD, provide the perfect environment for the acceleration of vascular calcification. Vascular calcification causes reduced arterial elasticity and increased blood pressure and pulse wave velocity (Fig. 2). In conjunction with increased FGF23 levels, all of these pathological phenotypes lead to the development of left ventricular hypertrophy and consequently heart dysfunction (Fig. 2). In hemodialysis patients, heart failure with left ventricular hypertrophy is commonly observed and is associated with cardiovascular mortality [56]. The effect of BAY-767 on vascular calcification was studied in rats with vascular calcification induced by a pan-FGF receptor inhibitor [53]. In this study, oral treatment with BAY-767 at the dose of 10 mg kg−1 reduced plasma Pi levels (∼1.4-fold) and aortic calcium content (∼75%). In contrast, the oral Pi binder lanthanum carbonate (2.2%, administered via diet) did not reduce aortic Ca2+ content [50▪▪].
 FIGURE 2:
FIGURE 2: Overall effects of Npt2a inhibition on renal excretion of minerals/electrolytes and the proposed cardioprotection. Npt2a inhibition by either PF-06869206 or BAY-767 enhances the urinary Pi excretion, resulting in reduced plasma Pi and PTH levels. The PTH secretion is regulated by calcium-sensing receptor (CaSR) expressed in the parathyroid gland. The reduction in PTH may have cardioprotective effects because of the PTH-induced hypertrophy of cardiomyocytes, Ca2+ overload in heart tissues, and oxidative stress. Inhibition of Npt2a with BAY-767 resulted in decreased FGF-23 levels (a potent stimulator for developing left ventricular hypertrophy (LV) in CKD). Therefore, reducing the FGF-23 level might benefit LV and possibly other heart diseases. The natriuretic and diuretic effects of Npt2a inhibition might be beneficial for lowering blood pressure and effective circulating volume (ECV). The increase in urinary Ca2+ excretion upon Npt2a inhibition is either by the direct inhibition of Ca2+ reabsorption in the proximal tubule or the indirect inhibition of PTH mediated Ca2+ transport (via transient receptor potential cation channel 5, TRPV5). Together, phosphaturic and calciuric effects of Npt2a inhibition might decrease the vascular calcification, arterial stiffness, and pulse wave velocity (PWV). A new study observed the increased expression of renal Npt2b in CKD; however, further studies are needed to confirm and determine its (patho)physiological importance. Reprinted with permission from Biochem Soc Trans. 2022;50(1):439–446. CKD, chronic kidney disease; Npt2a, sodium phosphate cotransporter 2a.
CONCLUSIONNpt2a inhibitors reduce renal Pi reabsorption in the proximal tubule. We are beginning to recognize that in addition to phosphaturia there are several accessory effects that might be indirectly related to inhibition of Npt2a. Notably, the renal handling of Na+, Cl−, and Ca2+ are affected. It remains to be seen if Npt2a inhibition is still efficacious in conditions where hyperphosphatemia is present, for example, in severe CKD, hemolysis, acute tumor lysis syndrome, rhabdomyolysis, etc. Clearly, further studies are needed to determine if long-term treatment with Npt2a inhibitors will, via a feedback mechanism, increase intestinal Pi uptake and/or result in changes in bone mineralization.
AcknowledgementsNone.
Financial support and sponsorshipThis work was supported by the National Institute of Diabetes and Digestive and Kidney Diseases grant 1R01DK110621 (to Dr Rieg), VA Merit Review Award IBX004968A (to Dr Rieg) and an American Heart Association Transformational Research Award 19TPA34850116 (to Dr Rieg). Financial support for this work was also provided by the NIDDK Diabetic Complications Consortium (RRID:SCR_001415,www.diacomp.org), grants DK076169 and DK115255 (to Dr Rieg). Dr Thomas was supported by an American Heart Association postdoctoral fellowship (828731).
Conflicts of interestDr Rieg had consultancy agreements with Ardelyx and Akros Pharmaceuticals.
REFERENCES AND RECOMMENDED READINGPapers of particular interest, published within the annual period of review, have been highlighted as:
▪ of special interest
▪▪ of outstanding interest
REFERENCES 1. Amanzadeh J, Reilly RF Jr. Hypophosphatemia: an evidence-based approach to its clinical consequences and management. Nat Clin Pract Nephrol 2006; 2:136–148. 2. Hruska KA, Mathew S, Lund R, et al. Hyperphosphatemia of chronic kidney disease. Kidney Int 2008; 74:148–157. 3. Biber J, Hernando N, Forster I. Phosphate transporters and their function. Annu Rev Physiol 2013; 75:535–550. 4▪. Levi M, Gratton E, Forster IC, et al. Mechanisms of phosphate transport. Nat Rev Nephrol 2019; 15:482–500. 5. Biber J, Hernando N, Forster I, Murer H. Regulation of phosphate transport in proximal tubules. Pflugers Arch 2009; 458:39–52. 6. Beck L, Karaplis AC, Amizuka N, et al. Targeted inactivation of Npt2 in mice leads to severe renal phosphate wasting, hypercalciuria, and skeletal abnormalities. Proc Natl Acad Sci USA 1998; 95:5372–5377. 7▪▪. Thomas L, Xue J, Tomilin VN, et al. PF-06869206 is a selective inhibitor of renal Pi transport: evidence from in vitro and in vivo studies. Am J Physiol Renal Physiol 2020; 319:F541–F551. 8. Segawa H, Kaneko I, Takahashi A, et al. Growth-related renal type II Na/Pi cotransporter. J Biol Chem 2002; 277:19665–19672. 9. Bergwitz C, Miyamoto KI. Hereditary hypophosphatemic rickets with hypercalciuria: pathophysiology, clinical presentation, diagnosis and therapy. Pflugers Arch 2019; 471:149–163. 10. Yamamoto T, Michigami T, Aranami F, et al. Hereditary hypophosphatemic rickets with hypercalciuria: a study for the phosphate transporter gene type IIc and osteoblastic function. J Bone Miner Metab 2007; 25:407–413. 11▪. Myakala K, Motta S, Murer H, et al. Renal-specific and inducible depletion of NaPi-IIc/Slc34a3, the cotransporter mutated in HHRH, does not affect phosphate or calcium homeostasis in mice. Am J Physiol Renal Physiol 2014; 306:F833–F843. 12. Segawa H, Onitsuka A, Furutani J, et al. Npt2a and Npt2c in mice play distinct and synergistic roles in inorganic phosphate metabolism and skeletal development. Am J Physiol Renal Physiol 2009; 297:F671–F678. 13. Liu L, Sanchez-Bonilla M, Crouthamel M, et al. Mice lacking the sodium-dependent phosphate import protein, PiT1 (SLC20A1), have a severe defect in terminal erythroid differentiation and early B cell development. Exp Hematol 2013; 41:432.e7–443.e7. 14. Bon N, Frangi G, Sourice S, et al. Phosphate-dependent FGF23 secretion is modulated by PiT2/Slc20a2. Mol Metab 2018; 11:197–204. 15. Motta SE, Imenez Silva PH, Daryadel A, et al. Expression of NaPi-IIb in rodent and human kidney and upregulation in a model of chronic kidney disease. Pflugers Arch 2020; 472:449–460. 16. Hernando N, Gagnon K, Lederer E. Phosphate transport in epithelial and nonepithelial tissue. Physiol Rev 2021; 101:1–35. 17. Blau JE, Collins MT. The PTH–Vitamin D–FGF23 axis. Rev Endocr Metab Disord 2015; 16:165–174. 18. Centeno PP, Herberger A, Mun HC, et al. Phosphate acts directly on the calcium-sensing receptor to stimulate parathyroid hormone secretion. Nat Commun 2019; 10:4693. 19. Takashi Y, Fukumoto S. Phosphate-sensing. Adv Exp Med Biol 2022; 1362:27–35. 20. Takashi Y, Kosako H, Sawatsubashi S, et al. Activation of unliganded FGF receptor by extracellular phosphate potentiates proteolytic protection of FGF23 by its O-glycosylation. Proc Natl Acad Sci USA 2019; 116:11418–11427. 21▪. Isakova T, Wahl P, Vargas GS, et al. Fibroblast growth factor 23 is elevated before parathyroid hormone and phosphate in chronic kidney disease. Kidney Int 2011; 79:1370–1378. 22. Shalhoub V, Shatzen EM, Ward SC, et al. FGF23 neutralization improves chronic kidney disease-associated hyperparathyroidism yet increases mortality. J Clin Invest 2012; 122:2543–2553. 23. Clinkenbeard EL, Noonan ML, Thomas JC, et al. Increased FGF23 protects against detrimental cardio-renal consequences during elevated blood phosphate in CKD. JCI Insight 2019; 4:e123817. 24. Vogt I, Haffner D, Leifheit-Nestler M. FGF23 and phosphate-cardiovascular toxins in CKD. Toxins (Basel) 2019; 11:647. 25. Faul C, Amaral AP, Oskouei B, et al. FGF23 induces left ventricular hypertrophy. J Clin Invest 2011; 121:4393–4408. 26. Chen J, Budoff MJ, Reilly MP, et al. Coronary artery calcification and risk of cardiovascular disease and death among patients with chronic kidney disease. JAMA Cardiol 2017; 2:635–643. 27. Eto N, Miyata Y, Ohno H, Yamashita T. Nicotinamide prevents the development of hyperphosphataemia by suppressing intestinal sodium-dependent phosphate transporter in rats with adenine-induced renal failure. Nephrol Dial Transplant 2005; 20:1378–1384. 28. Pulskens WP, Verkaik M, Sheedfar F, et al. Deregulated renal calcium and phosphate transport during experimental kidney failure. PLoS One 2015; 10:e0142510. 29. Kaludjerovic J, Komaba H, Sato T, et al. Klotho expression in long bones regulates FGF23 production during renal failure. FASEB J 2017; 31:2050–2064. 30. Pawlak D, Znorko B, Kalaska B, et al. LP533401 restores bone health in 5/6 nephrectomized rats by a decrease of gut-derived serotonin and regulation of serum phosphate through the inhibition of phosphate co-transporters expression in the kidneys. Bone 2018; 113:124–136. 31. Drueke TB, Massy ZA. Lowering expectations with niacin treatment for CKD-MBD. Clin J Am Soc Nephrol 2018; 13:6–8. 32. Block GA, Wheeler DC, Persky MS, et al. Effects of phosphate binders in moderate CKD. J Am Soc Nephrol 2012; 23:1407–1415. 33. Covic A, Rastogi A. Hyperphosphatemia in patients with ESRD: assessing the current evidence linking outcomes with treatment adherence. BMC Nephrol 2013; 14:153. 34. Schiavi SC, Tang W, Bracken C, et al. Npt2b deletion attenuates hyperphosphatemia associated with CKD. J Am Soc Nephrol 2012; 23:1691–1700. 35. Marks J, Churchill LJ, Srai SK, et al. Intestinal phosphate absorption in a model of chronic renal failure. Kidney Int 2007; 72:166–173. 36. Sabbagh Y, O’Brien SP, Song W, et al. Intestinal npt2b plays a major role in phosphate absorption and homeostasis. J Am Soc Nephrol 2009; 20:2348–2358. 37▪. Larsson TE, Kameoka C, Nakajo I, et al. NPT-IIb inhibition does not improve hyperphosphatemia in CKD. Kidney Int Rep 2018; 3:73–80. 38▪. Maruyama S, Marbury TC, Connaire J, et al. NaPi-IIb inhibition for hyperphosphatemia in CKD hemodialysis patients. Kidney Int Rep 2021; 6:675–684. 39. Malhotra R, Katz R, Hoofnagle A, et al. The effect of extended release niacin on markers of mineral metabolism in CKD. Clin J Am Soc Nephrol 2018; 13:36–44. 40. Lenglet A, Liabeuf S, El Esper N, et al. Efficacy and safety of nicotinamide in haemodialysis patients: the NICOREN study. Nephrol Dial Transplant 2017; 32:870–879. 41▪▪. Tsuboi Y, Ohtomo S, Ichida Y, et al. EOS789, a novel pan-phosphate transporter inhibitor, is effective for the treatment of chronic kidney disease-mineral bone disorder. Kidney Int 2020; 98:343–354. 42. Hill Gallant KM, Stremke ER, Trevino LL, et al. EOS789, a broad-spectrum inhibitor of phosphate transport, is safe with an indication of efficacy in a phase 1b randomized crossover trial in hemodialysis patients. Kidney Int 2021; 99:1225–1233. 43▪. King AJ, Siegel M, He Y, et al. Inhibition of sodium/hydrogen exchanger 3 in the gastrointestinal tract by tenapanor reduces paracellular phosphate permeability. Sci Transl Med 2018; 10:eaam6474. 44▪. Block GA, Rosenbaum DP, Yan A, Chertow GM. Efficacy and safety of tenapanor in patients with hyperphosphatemia receiving maintenance hemodialysis: a randomized phase 3 trial. J Am Soc Nephrol 2019; 30:641–652. 45. Pergola PE, Rosenbaum DP, Yang Y, Chertow GM. A randomized trial of tenapanor and phosphate binders as a dual-mechanism treatment for hyperphosphatemia in patients on maintenance dialysis (AMPLIFY). J Am Soc Nephrol 2021; 32:1465–1473. 46. Xue J, Thomas L, Tahmasbi M, et al. An inducible intestinal epithelial cell-specific NHE3 knockout mouse model mimicking congenital sodium diarrhea. Clin Sci (Lond) 2020; 134:941–953. 47▪▪. Xue J, Thomas L, Murali SK, et al. Enhanced phosphate absorption in intestinal epithelial cell-specific NHE3 knockout mice. Acta Physiol (Oxf) 2022; 234:e13756. 49▪. Filipski KJ, Sammons MF, Bhattacharya SK, et al. Discovery of orally bioavailable selective inhibitors of the sodium-phosphate cotransporter NaPi2a (SLC34A1). ACS Med Chem Lett 2018; 9:440–445. 50▪▪. Klar J, Anja G, Ehrmann A, et al. Inhibition of sodium phosphate transporter Npt2a improves experimental vascular calcification and phosphate homeostasis in rats (ndt abstract MO053). Nephrol Dial Transplant 2020; 35:35doi: 10.1093/ndt/gfaa140.MO053. 51▪▪. Clerin V, Saito H, Filipski KJ, et al. Selective pharmacological inhibition of the sodium-dependent phosphate cotransporter NPT2a promotes phosphate excretion. J Clin Invest 2020; 130:6510–6522. 52▪▪. Thomas L, Xue J, Murali SK, et al. Pharmacological Npt2a inhibition causes phosphaturia and reduces plasma phosphate in mice with normal and reduced kidney function. J Am Soc Nephrol 2019; 30:2128–2139. 53. Klar J, Anja G, Ehrmann A, et al. Inhibition of sodium phosphate transporter npt2a increases urinary phosphate excretion and improves experimental vascular calcification in rats (kidney week abstract TH-PO542). J Am Soc Nephrol 2019; 30: 258. 54. Thomas L, Bettoni C, Knopfel T, et al. Acute adaption to oral or intravenous phosphate requires parathyroid hormone. J Am Soc Nephrol 2017; 28:903–914. 55. Tomoe Y, Segawa H, Shiozawa K, et al. Phosphaturic action of fibroblast growth factor 23 in Npt2 null mice. Am J Physiol Renal Physiol 2010; 298:F1341–F1350. 56. House AA, Wanner C, Sarnak MJ, et al. Heart failure in chronic kidney disease: conclusions from a Kidney Disease: Improving Global Outcomes (KDIGO) controversies conference. Kidney Int 2019; 95:1304–1317.
留言 (0)