
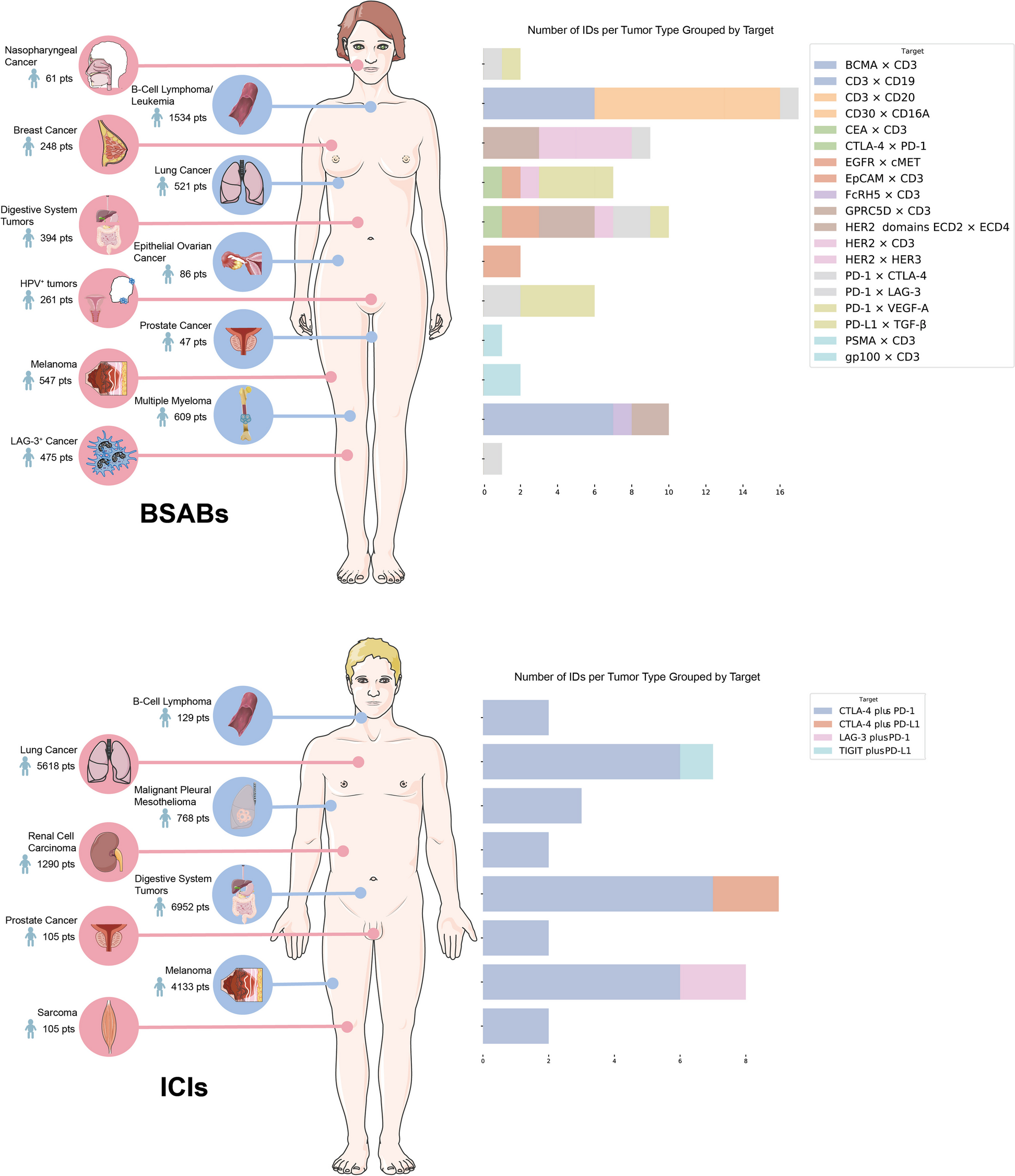
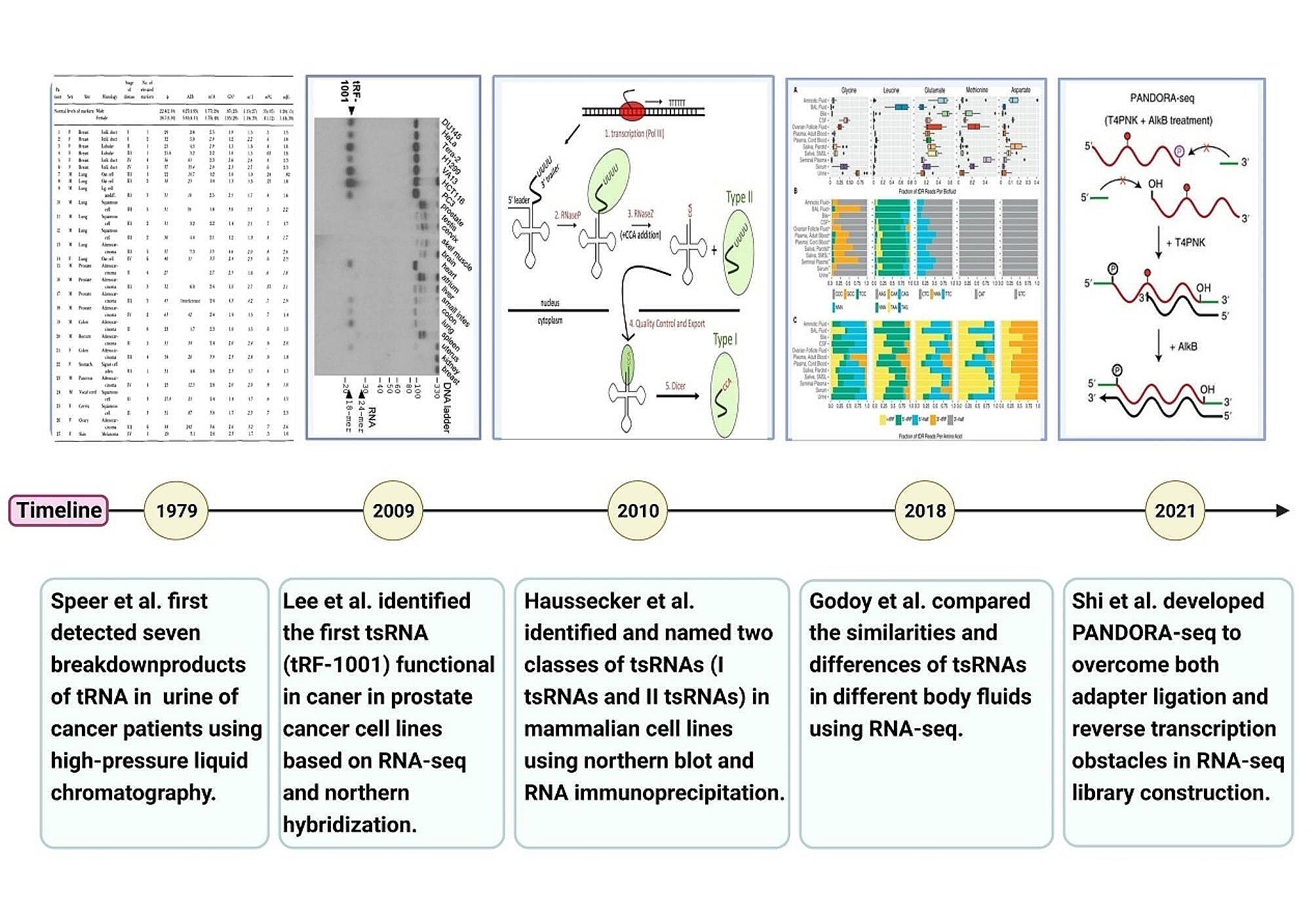
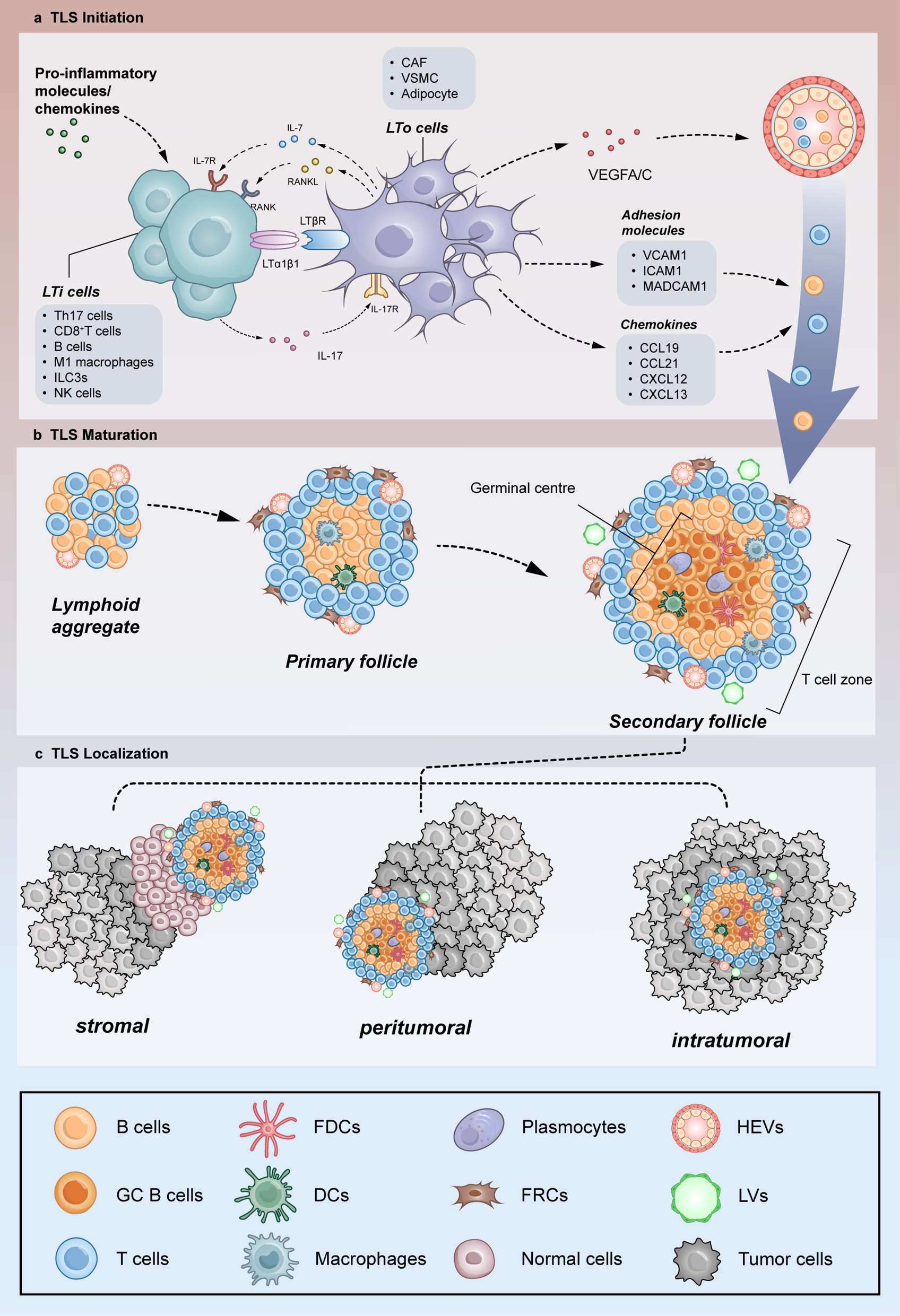
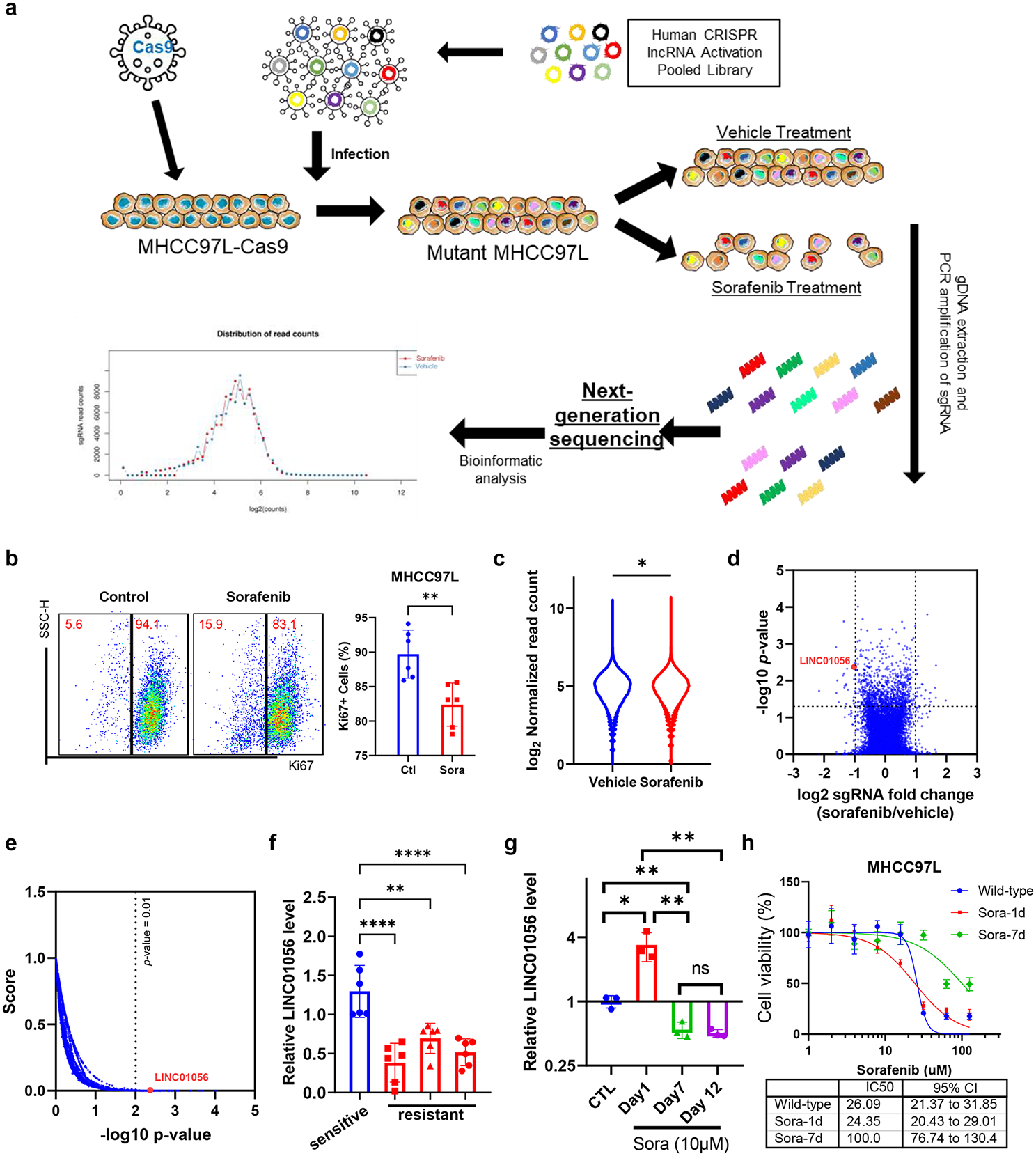
記住我
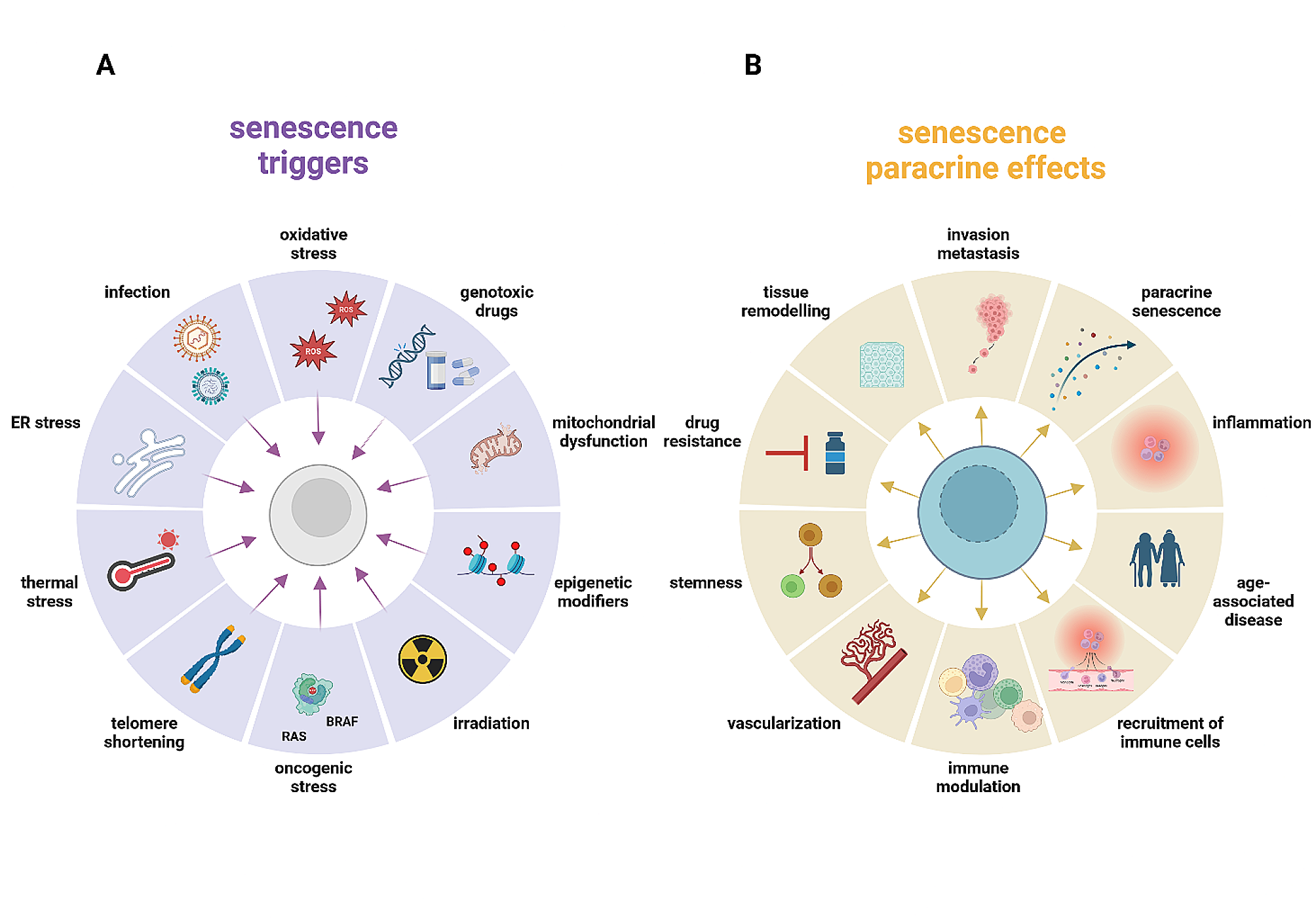
We designed and validated a multiplex allele-specific real-time polymerase chain reaction (qPCR) genotyping assay to detect three EBV BALF2 variants (V700L, I613V, V317M; Supplementary Methods, Fig. S1, Tables S1–4). The wild-type V700 allele was selected as an internal control for samples lacking these polymorphisms. The assay’s 95% lower limit of detection was 2.0 copies/reaction (95% CI 1.4–2.6) with < 20% coefficient of variation across six orders of magnitude (R2 ≥ 0.992, Fig. 1A-B, Tables S12–13). Non-specific amplification was not observed for off-target alleles, and replicates of the B95–8 wild-type whole-virus control also confirmed specificity. In mixing experiments ranging from 0 to 100% allele frequency, the assay detected allele frequencies as low as 10% for each of the four targets, below the host heterozygosity threshold (Fig. 1C, Table S14) [9].
Fig. 1
Multiplex EBV BALF2 genotyping qPCR design, validation, and association studies with nasopharyngeal carcinoma in endemic and non-endemic populations. A Analytical sensitivity for each of the four BALF2 qPCR targets. The 95% lower limit of detection with 95% confidence interval is reported for each target in units of EBV copies/mL plasma. In conjunction with the LLODs, the corresponding plasma viral load for 34 screen-detected preclinical NPC cases is presented to indicate likelihood of genotyping success. B Analytical linearity for each of the four BALF2 qPCR targets, plotting cycle threshold (Ct) against nominal dsDNA control concentration in units of log10 copies/μL template. C Mixing studies at fixed total template concentration (100 copies/μL template) combining high-risk and low-risk dsDNA controls, demonstrating detection of minor allele fractions as low as 10% for each of the four targets. Measured concentration is plotted against nominal concentration. In the presence of mixed alleles, the assay is approximately linear as allele fraction decreases. D Study overview and experimental workflow. First, the multiplex BALF2 genotyping assay was analytically validated using synthetic dsDNA controls and wild-type B95–8 whole virus control. Next, our non-endemic cohort of 24 NPC cases and 155 non-NPC controls contributed to BALF2 qPCR/NGS validation, longitudinal BALF2 genotyping, and BALF2-NPC association. Finally, our non-endemic cohort and three predominantly endemic cohorts contributed to a meta-analysis of 755 EBV+ NPC cases and 981 non-NPC controls. This validated the association between BALF2 haplotypes and NPC in multiple cohorts, further defined regional EBV genomic diversity, and was used to develop a variant-informed screening model. E Prevalence of I613V and V317M between EBV+ NPC cases and non-NPC controls in the present study and in the three prior EBV GWAS cohorts. F Log-transformed odds ratios with 95% confidence intervals for association between BALF2 high-risk haplotypes (C-C-T, C-C-C, or both) and EBV+ NPC or other EBV-associated diseases in the current cohort and in the three prior EBV GWAS cohorts. G Individual patient characteristics from current study of 24 NPC cases and 155 non-NPC controls. BALF2 haplotypes are defined by presence or absence of V700L, I613V, and/or V317M, which are associated with clinical phenotype. H Plasma EBV viral load (log10 IU/mL plasma) across phenotypes of 155 patients included in current study, demonstrating no significant difference between plasma viral load and phenotype. I and J Association between NPC and other BALF2 single nucleotide variants identified by next-generation sequencing. Log-transformed P-value from association test and log-transformed odds ratios with 95% confidence intervals are presented for three variants of interest (V700L = 162215C > A, I613V = 162476C > T, V317M = 163364C > T) and 13 additional variants differentially associated with NPC. V700L is mutually exclusive with I613V and V317M, was rare in this population, and was not associated with NPC risk. Only one other variant (163287G > A, synonymous) exceeded the Bonferroni-corrected P-value threshold
Multiplex BALF2 genotyping qPCR has near-perfect concordance with next-generation sequencingWe sequenced the BALF2 region in 258 clinical plasma specimens genotyped by qPCR, and 152 had adequate sequencing depth and coverage (Supplementary Methods). Samples with adequate sequencing depth and coverage had higher viral load (median 1600 vs. 201 IU/mL, p < 0.01). There was a single discordant genotyping call between qPCR and NGS. In a 43-year-old immunosuppressed woman with heart/lung transplantation, the sixth of six plasma specimens collected over 4.9 months showed qPCR loss of I613V which was detected on all five prior specimens. The specimen was sequenced and revealed the I613V mutation in 35/36 (97.2%) reads, reflecting false negative qPCR, possibly due to low viral load (EBNA-1 < 100 IU/mL). Positive and negative percent agreements for V700L, I613V, and V317M were otherwise 100%, and overall haplotype concordance between qPCR and NGS was 99.3% (151/152, Table S15).
BALF2 haplotypes are associated with NPC in a non-endemic cohortWe genotyped plasma specimens from 179 unique patients in a non-endemic population, including 155 non-NPC controls and 24 EBV-positive NPC cases (Table S5, Fig. 1G). Among controls, the most common indication for plasma EBV PCR was monitoring after solid organ transplant (44%) or bone marrow transplant (33%). Seventy-six control patients (49%) had hematologic neoplasms with (66%) or without (33%) prior bone marrow transplant, including EBV-positive lymphomas/leukemias. Nineteen patients (12%) had no history of transplant or neoplasm, including ten patients with primary EBV infection. There was no significant association between plasma EBV EBNA-1 viral load and disease phenotype (Fig. 1H).
High-risk BALF2 haplotypes, defined by the presence of I613V with or without V317M, were rare among non-NPC controls (Fig. 1D-E, Table S16). The C-C-C and C-C-T high-risk haplotypes were present in 5.8 and 1.3% of controls, compared with 12.5 and 62.5% of NPC cases. Using the low-risk A-T-C and C-T-C haplotypes as reference, both the C-C-C (odds ratio [OR] 7.9 95% confidence interval [CI] 1.7–37.1) and C-C-T (OR 178.8, 95% CI 33.1–965.3) haplotypes were highly associated with NPC in this non-endemic population (Fig. 1F, Table S16). We observed no association between these haplotypes and other diseases, including hematologic neoplasms.
BALF2 haplotypes are associated with NPC in a meta-analysis of endemic and non-endemic cohortsIn a meta-analysis of 755 NPC cases and 981 non-NPC controls from this study and three previously-published EBV GWAS, the NPC odds ratios for the C-C-C and C-C-T haplotypes were 4.0 (95% CI 2.6–6.0) and 15.4 (95% CI 11.2–21.0), respectively (Fig. 1D-F, Table S16). While I613V and V317M were common (> 75%) in NPC cases across cohorts, they were uncommon in non-endemic controls (7.1%) relative to endemic controls (60.5%), suggesting that variable NPC incidence could be explained by underlying BALF2 haplotype prevalence.
We also evaluated the association between clinical phenotypes and other BALF2 SNVs. For example, the previously-described 162507C > T and 162852G > T synonymous polymorphisms have been rarely observed (3%) in NPC cases but are common in endemic controls (41–43%). Among 108 unique patients with sequenced specimens, neither mutation was significantly associated with NPC. Beyond I613V and V317M, only the synonymous 163287G > A SNV reached statistical significance (Fig. 1I-J, Table S17). We observed no BALF2 SNVs which were significantly associated with EBV-positive leukemias/lymphomas or post-transplant lymphoproliferative disorders. We also assessed whether other SNVs were associated with high-risk BALF2 haplotypes, and identified multiple variants which were significantly correlated with I613V and V317M. For example, seven BALF2 SNVs occur with 100% frequency in the I613V/V317M haplotype and with 0–2% frequency in low-risk haplotypes (p ≤ 1.31 × 10− 7). This supports the hypothesis that high-risk EBV variants are transmitted locally rather than developing de novo after primary infection (Table S18).
Longitudinal genotyping within hosts confirms temporal stability of BALF2 haplotypesBecause EBV establishes lifelong latent infection, BALF2 genotyping could facilitate once-lifetime screening triage. To assess whether EBV BALF2 haplotypes persisted over time, we genotyped 90 EBV-positive plasma specimens collected from a subset of 16 patients. These patients had a median of 5 (range, 2–7) specimens genotyped over a median period of 8.6 months (range, 2.8–13.9). Among the 90 genotyped specimens, 88 (97.8%) haplotype calls were concordant within a given individual over time (Fig. 2A). The two discordant specimens both occurred in individuals with solid organ transplantation (Fig. 2A, Patients #2 and #10) and may represent mutagenesis under immunosuppression or reactivation of distinct latent infections from the host and donor tissue.
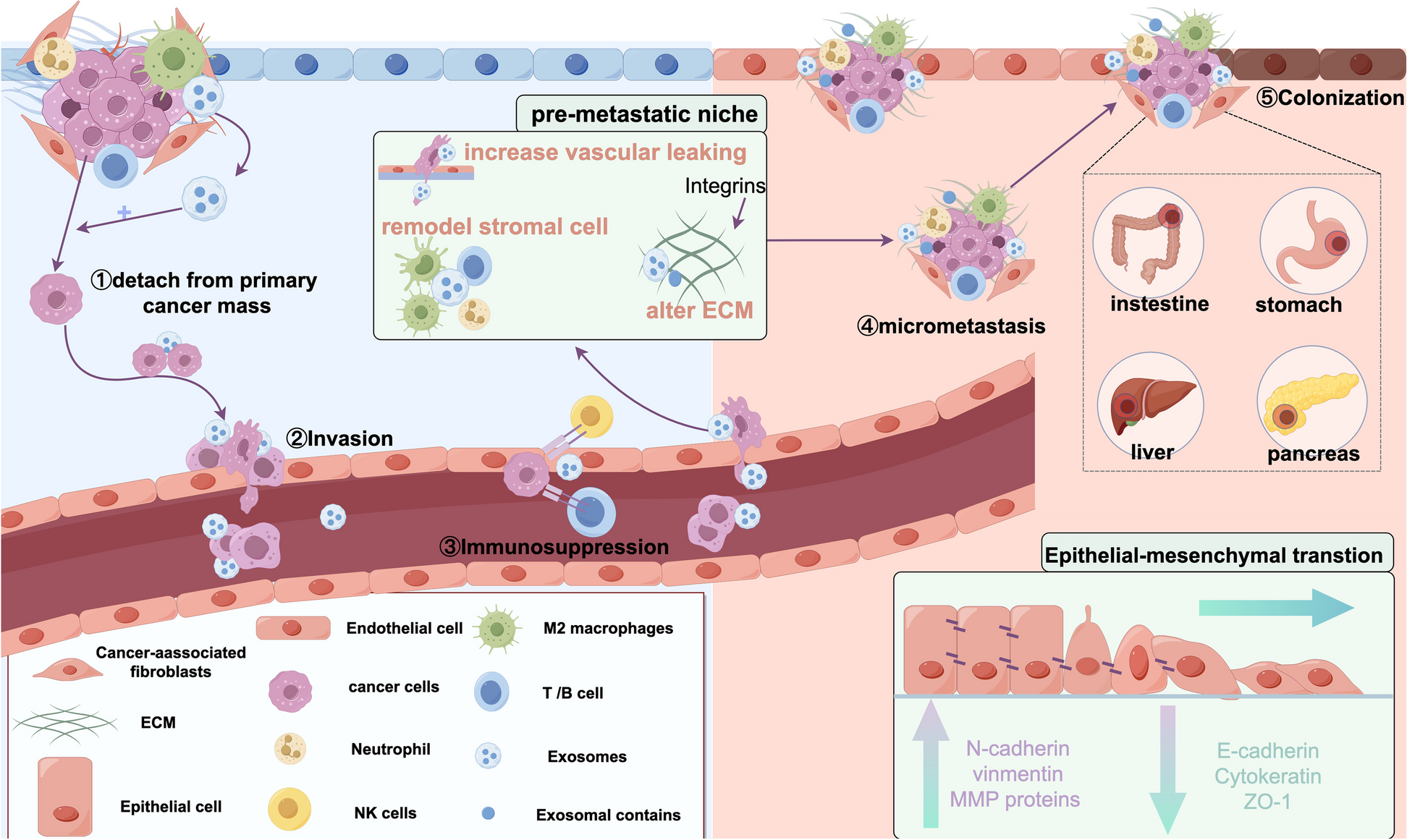
Fig. 2
Longitudinal EBV BALF2 genotyping and modeled variant-informed NPC screening strategies in 12 high-risk endemic populations. A A subset of 16 patients with serial EBV-positive plasma specimens were subject to BALF2 genotyping by qPCR and NGS to assess temporal haplotype stability. Variant allele fraction (VAF) is plotted against time from first specimen collection for the three qPCR targets (V700L, I613V, V317M). The sample’s viral load in log10 EBNA-1 IU/mL is plotted below the allele frequencies. Two patients had one specimen each with temporally-discordant haplotypes. Patient #2 was a lung/liver transplant recipient with I613V detected in only the third of seven plasma specimens collected over 8.7 months. Patient #10 was a kidney transplant recipient with large-cell lymphoma who had V700 detected only in the first of five specimens collected over 7.8 months, whereas the subsequent four specimens harbored V700L, possibly indicating mutagenesis. B Map of east/southeast Asia with 12 included high-risk populations. Shading represents the national NPC incidence rate. Each bubble indicates a single population with size proportional to incidence rate. Bubble color indicates the cost-effectiveness of variant-informed screening at variable willingness-to-pay thresholds. C Modeled survival in a hypothetical cohort of 50-year-old patients in southern China. Survival differs with no screening (black line), seven variant-agnostic screening strategies (red solid lines), and seven variant-informed screening strategies (blue dashed lines) due to weighted stage distributions dictated by effective screening sensitivity. D Cost-effectiveness of variant-agnostic and variant-informed screening strategies across variable screening frequencies. Box plots indicate median with interquartile range. E Resource utilization after initial biomarker screening for variant-agnostic (A0-G0) and variant-informed screening strategies (ABALF2-GBALF2). Bar charts indicate absolute number of screening endoscopies and MRIs per 100,000 screened subjects. Referrals for endoscopy/MRI decrease after triage with BALF2 qPCR. F NPC deaths per 100,000 screened individuals with variable screening frequencies and initial screening ages
Variant-informed NPC screening strategies reduce false positives and unnecessary proceduresWe estimated population-level NPC mortality reduction, resource utilization, and cost-effectiveness of BALF2 variant-informed screening strategies using a previously-validated time-inhomogeneous decision-analytic cohort model (Fig. S2) [14]. Full details regarding the model framework, population selection, screening strategies, and sensitivity analyses are provided in the Supplementary Methods and Tables S6–9.
First, we conducted a meta-analysis of three prior EBV GWAS to model endemic BALF2 haplotype prevalence among NPC cases and non-NPC controls [9,10,11]. Thereafter, we compared seven variant-agnostic screening strategies from prospective studies to seven variant-informed strategies wherein positive plasma/nasopharyngeal EBV PCR are triaged using the BALF2 genotyping qPCR (Table S10). Twelve high-risk populations in southern China, Hong Kong SAR, Macao SAR, Republic of China, and Singapore met inclusion criteria (Fig. 2B, Table S11).
Variant-informed screening increased PPV by a median of 46% (range, 26–51%) with an absolute decrease in screening sensitivity of 7%. Variant-informed screening reduced referrals for endoscopy and/or MRI by approximately 40% relative to the corresponding variant-agnostic strategy (Table S10). This reduction in referrals for further screening steps averted a median of 2969 screening visits per 100,000 subjects (Table S19).
For a hypothetical cohort of 50-year-old men and women who develop NPC in southern China, 10-year survival improved from 70.4% (95% CI 68.1–72.5%) in an unscreened cohort to a median of 85.7% (range, 85.4–87.0%) with variant-agnostic screening and 85.2% (range, 84.3–85.9%) with variant-informed screening (Fig. 2C, Table S19). In the highest incidence region, the small reduction in screening sensitivity after BALF2 triage resulted in approximately 3.4 excess NPC deaths and 600 fewer false-positives requiring endoscopy/MRI per 100,000 subjects screened.
Variant-informed NPC screening is cost-effective and facilitates once-lifetime testingThe base case screened adult men and women once at age 50 years (Fig. 2D, Table S19–20). Variant-informed screening was cost-effective in all populations except Hengdong, China (due to lower NPC incidence). Across the 12 populations and 14 screening strategies, an initial screening age of 40–45 tended to be most cost-effective irrespective of screening interval (Table S21). Screening intervals as short as every two years could be cost-effective. Variant-informed screening became more cost-effective as the number of lifetime screens increased due to the increasing proportion of subjects known to have low-risk BALF2 haplotypes that were never subsequently screened (Fig. S3). Sensitivity analysis identified parameters that most impacted cost-effectiveness (Fig. S3, Tables S22–23).
留言 (0)